Роланд Глазер
Биология в новом свете
Перевод с немецкого канд. биол. наук К. М. Близник и С. В. Беневоленского
под редакцией д-ра биол. наук Ю. Г. Капульцевича
ИЗДАТЕЛЬСТВО "МИР" МОСКВА 1978
ИБ № 1010
Редактор А. Кондрашова
Художник Ф. Инфанте
Художественный редактор Л. Безрученков
Технический редактор Н. Манохина
Корректор Н. Матюхина
Предисловие редактора перевода
Последние два-три десятилетия биология переживает период бурного развития. Замечательные успехи, достигнутые во многих областях этой науки, привели к "информационному взрыву". Количество биологической литературы лавинообразно растет с каждым годом, так что даже специалисты, непосредственно занимающиеся биологией, не в состоянии уследить за развитием всех многочисленных биологических дисциплин. Представители других наук чувствуют себя еще более беспомощными перед этим морем информации. В особенно трудном положении оказывается молодежь, желающая посвятить себя науке. Современная наука, и биология в частности, многогранна, а в учебниках находят отражение лишь твердо установленные факты и теории, выдержавшие проверку временем. Как же выбрать свой путь? Здесь на помощь должна прийти научно-популярная литература.
За последние годы наши издательства выпустили немало научно-популярных книг по биологии, но все они были посвящены каким-то отдельным ее разделам, в основном вопросам генетики, молекулярной биологии и микробиологии.
Предлагаемая вниманию читателя книга Роланда Глазера, профессора Берлинского университета им. Гумбольдта, выгодно отличается от других научно-популярных изданий на эту тему по крайней мере в двух отношениях.
Во-первых, автор рассматривает биологию как науку в целом, а не какие-то отдельные ее области. Читатель знакомится с различными процессами, протекающими на разных уровнях биологической организации, — от молекул до животного и даже целых сообществ живых организмов, так называемых биоценозов.
Во-вторых, основное внимание в книге уделено вопросам, которые в научно-популярной биологической литературе, как правило, вообще не обсуждаются, а в обширных курсах по общей биологии упоминаются лишь мимоходом. Это вопросы, связанные с использованием в биологии физики и математики. Книгу Р. Глазера можно было бы назвать "Жизнь с точки зрения физика", если бы сходное название не было уже использовано выдающимся австрийским физиком Э. Шредингером для своей известной книги, носящей более специальный характер
[1].
Глазер предлагает взглянуть на биологию по-новому. Он обращает внимание читателя на целый ряд биологических явлений и процессов, механизмы которых можно понять и объяснить, привлекая на помощь математический аппарат, физические законы и теории. Проблемы эти довольно сложны, многое в их решении остается неясным и по сей день, но автор ведет свой рассказ увлекательно, в доступной для непосвященного читателя форме. И это, несомненно, большое достоинство книги.
Первые же ее страницы буквально интригуют читателя: автор приводит удивительные результаты сопоставления биологических и технических объектов. Телевизионная башня кажется нам очень высокой и стройной. Как, наверное, трудно спроектировать и построить такое сооружение! Отношение высоты башни к ее диаметру составляет примерно 20. Много это или мало? Оказывается, не так уж много, ибо у обыкновенного тростника это отношение равно 200! Муха-журчалка за одну секунду пролетает расстояние, в 1000 раз превышающее длину ее тела. Если бы скорость самолета находилась в таком же соотношении с его длиной (равной, скажем, 30 м), то она должна была бы достигать 30 км/с — в 100 раз больше скорости звука! Значит ли это, что технические объекты безнадежно отстают от живых организмов? Прочитав книгу, читатель получит ответ и на этот, и на многие другие вопросы, подчас совершенно неожиданные.
Наивные детские "почему?", от которых столь часто с досадой отмахиваются взрослые! Почему из мухи нельзя сделать слона? Почему слон не больше, а мышь не меньше, чем они есть на самом деле, и т. д. Автор призывает не бояться таких вопросов. Часто они помогают нам взглянуть на привычный мир иначе и увидеть в нем что-то новое...
Математика поистине всесильна. Оказывается, она помогает нам описать форму живого организма и проследить ее изменение в процессе эволюции. В настоящее время много говорится о теории оптимальных процессов, которая позволяет решать сложные вопросы экономики, управления народным хозяйством. Но эта же теория находит широкое применение и в биологии.
Современная биология использует и термодинамику. Законы термодинамики позволяют объяснить, откуда живые организмы берут необходимую энергию, как эта энергия преобразуется в процессе жизнедеятельности организмов, в каких формах она накапливается в биологических системах и как расходуется ими.
Читатель вместе с автором совершает увлекательное путешествие в царство молекул, где перед ним открывается совершенно иной, не знакомый ему мир — мир, в котором многие привычные представления теряют смысл. Читатель узнает, как происходит синтез белковых молекул в клетке, какое важное значение имеет хаотическое тепловое движение молекул для протекания биохимических реакций, для самой жизни.
В книге Р. Глазера затронут чрезвычайно широкий круг вопросов, и вполне естественно, что отнюдь не на все из них автор мог дать исчерпывающие ответы. Подробное изложение всех этих вопросов потребовало бы чрезмерного увеличения объема книги. Кроме того, многие из них еще ждут своего решения. Однако главная задача автора — пробудить у читателя интерес к тем проблемам биологии, для решения которых необходимо знание физики и математики.
И нам кажется, что с этой задачей он удачно справился.
Книга "Биология в новом свете" адресована в первую очередь школьникам старших классов, перед которыми стоит большая и трудная проблема — найти свой путь в жизни" И мы надеемся, что, ознакомившись с этой книгой, многие школьники с математическим складом ума поймут, что и в биологии перед ними открываются широкие возможности для применения своих способностей, Эту книгу с интересом прочтут и специалисты — как биологи, так и представители других наук, а преподавателям биологии она несомненно принесет особую пользу.
Ю. Капульцевич
Введение. Тем, кто хочет изучать биологию, хотя и не собирается стать директором зоопарка
Биология вчера, сегодня, завтра. Кто такой биолог? Нужны ли ему математика и физика? Является ли биология точной наукой? Должен ли биолог только собирать и сравнивать или можно подойти к изучению биологии как-то иначе?
Итак, кто же такой биолог? Разные люди наверняка ответили бы на этот вопрос по-разному. "Биолог — человек с ботанизиркой и сачком для ловли бабочек, чудак, работа которого больше похожа на хобби", — сказал бы один. "Биолог — это исследователь, который выращивает в реторте людей, людей с выдающимися способностями, сверхлюдей будущего", — заявил бы другой. Оба этих крайних суждения одинаково абсурдны; между ними, вероятно простирается широкий спектр мнений. И это не удивительно, поскольку ни в одной науке за последние десятилетия не произошли такие революционные изменения, как в биологии. Сейчас биология находится где-то в середине этого этапа развития, и вряд ли кто-нибудь возьмется дать долгосрочный прогноз на будущее. Часто даже самим биологам нелегко бывает уследить за новыми направлениями, возникающими в той огромной области науки, которая связана с познанием жизни — высшей формы движения. Насколько же трудно должно быть юноше, школьнику, когда ему приходится решать, в какой области работать, какой профессии больше всего соответствуют его способности и интересы! Для чего изучают биологию? Чтобы стать директором зоопарка? Не зря ли стараются заработать пятерки по математике и физике те, кто собирается заниматься "всего лишь" биологией? Можно ли отнести биологию к "точным" наукам?
Издавна человек, видя перед собой изобилие биологических форм — растений и животных, — испытывал потребность разобраться во всем этом многообразии, систематизировать его. С этого и началась биология. В наше время кое-кто склонен недооценивать биологов, занимающихся сбором растений и наблюдением за животными: "Гербариями и коллекциями бабочек пристойно заниматься юным натуралистам или взрослым в свободное время!" Но не следует забывать, что любые успехи современной биологии — будь то новые нейрофармакологические препараты и антибиотики, знания о наследственных болезнях или новые сорта и породы полезных растений и животных и многое другое — были бы невозможны без работ наших неустанно коллекционировавших и классифицировавших предшественников. Что делали бы мы сейчас без детальнейших сведений о многообразии биологических форм и их систематизации! И хотя центр тяжести биологических исследований находится теперь не в области систематики, многие вопросы в ней остаются по-прежнему открытыми и продолжают служить предметом научных дискуссий.
Наряду с исследованием видов, их морфологического и анатомического строения ученые изучают механизмы жизнедеятельности. Эту область биологии называют физиологией. Почему бьется сердце? У всех ли животных оно есть? Как оно работает у тех или иных животных? Как связан образ жизни живого существа с функциями его органов? В физиологии аспект сравнения, систематизации выражен еще достаточно сильно и играет заметную роль, хотя главным здесь, конечно, является вопрос о принципах строения организма. Элементы сравнения можно обнаружить даже в одном из самых современных направлений биологии — биохимии. Из каких химических кирпичиков сложен организм? Какие в нем протекают химические реакции и зачем они нужны? Что общего или различного между реакциями у разных организмов?
"В настоящее время все более возрастает стремление выявить функциональные связи между отдельными элементами живых организмов, разобраться в механизмах биологических процессов; неуклонно расширяется использование в биологии химических методов исследования. Но вместе с тем все чаще напрашивается мысль: а нельзя ли применить в биологии физические законы, справедливые для неживой природы? Не пригодится ли физическая наука с ее столь солидной аналитической базой для познания функциональных схем животных и растений?
В сущности, эта мысль такая же древняя, как сами биология и физика. На каждой ступени развития науки люди снова и снова пытались приложить физические знания к исследованию биологических систем. Но каждая такая попытка проникнуть в тайны живого кончалась неудачей. Попробуйте, пренебрегая этим опытом, бездумно применить недопустимо упрощенные физические постулаты для объяснения биологических явлений, и вы обязательно натолкнетесь на противоречия. А отсюда неизбежно следовал вывод: законы живого мира, — очевидно, законы особого рода. Живые организмы, вероятно, не подчиняются законам, действующим в неживых системах. И вот вследствие незрелости естественной науки на этой почве расцвели философские учения, утверждавшие некую жизненную силу —
vis vitalis[2].
Повсюду вода течет сверху вниз. Но в деревьях она движется снизу вверх, от корней к листьям. Значит ли это, что здесь нарушен закон тяготения? К такому заключению неизбежно пришел бы естествоиспытатель, если бы он применил к биологической системе исключительно законы гидростатики. И только обратившись к термодинамике, мы можем понять, что никакая таинственная
vis vitalis не противостоит здесь физическим законам. Просто в данном случае мы имеем дело с обычным осмотическим давлением, наблюдаемым, кстати, и в неживой природе; это давление противодействует силе тяготения, конечно, действующей и в растениях, и вызывает подъем воды в стволе.
Если при изучении биологических процессов не ограничиваться простым описанием их, а стремиться выяснить управляющие ими механизмы (в физическом смысле этого слова), то, учитывая сложность биологических систем, следует обратиться к физике. Но, поскольку физика также находится в процессе развития, она к сожалению, не в состоянии объяснить многие явления жизни. Действительно, для объяснения некоторых даже простейших биологических явлений приходится прибегать к очень сложным физическим понятиям. И не только физика, но и связанная с ней математика подчас не способны нам помочь. Даже большие успехи в области электронно-вычислительной техники, позволившие произвести кое-какие расчеты биологических систем, еще недостаточны для решения сложных систем уравнений, описывающих поведение живых систем.
Но не будем унывать, а попытаемся прикинуть, что же все-таки могут дать нам здесь современные физика и математика. "Выжимая" из этих небиологических дисциплин все возможное, мы тем самым подтолкнем их развитие. Во многих случаях уже сейчас использование в биологии физических и математических методов приносит успех. Наши знания, наши научно-технические достижения, по-видимому, находятся на том уровне, который позволяет осмысленно использовать физику и математику для изучения жизни. К этим достижениям относятся прежде всего такие крупные теоретические дисциплины, как волновая механика, статистическая физика, термодинамика; высокочувствительные электронные устройства, позволяющие с очень высокой точностью определять чрезвычайно малые электрические и магнитные потенциалы биологических макромолекул, клеток и организмов, и, наконец, электронные вычислительные машины, производящие с фантастической скоростью счетные операции, для выполнения которых человеку с карандашом и бумагой в руках понадобились бы годы и даже десятки лет.
Таким образом, не удивительно, что в последние десятилетия бурно развиваются биофизика и биоматематика. Все шире становится круг исследователей, работающих в этих смежных областях, все больше появляется научных статей. Но что знает об этом развитии небиолог?
Так попытаемся задуматься над некоторыми из их проблем и теорий. Попробуем взглянуть на биологию не в традиционном плане, а с иной точки рения. Для этого, несомненно, необходим весь комплекс знаний физики и математики, которым читатель, может быть, и не обладает. Поэтому в нашей книге мы рассмотрим такие примеры, которые позволят даже несведущему в естественных науках человеку понять основные принципы. Мы начнем с повседневного, с того, что нас окружает, познакомимся с удивительными свойствами и особенностями растений и животных. Скоро мы убедимся, что физическое объяснение форм и функций возможно только в результате очень серьезных размышлений. Так, переходя со ступени на ступень, от живых организмов вплоть до молекулярного уровня, мы узнаем, на чем основана в биологии причинная связь между функцией и формой, между временем и пространством.
Большое и малое в сравнении
Былинка и телевизионная башня! Муха-журчалка и реактивный самолет! Жужелица и гоночный автомобиль! Есть ли смысл в этих сравнениях? Чему может поучиться инженер у живых организмов? Почему деревья не растут до неба и почему из мухи нельзя сделать слона? Почему слон не больше, а землеройка не меньше, чем они есть? Длина прыжка блохи во много раз превышает длину ее тела. Почему же так не может прыгнуть кенгуру? Теория подобия в фармакологии, медицине и спорте.
С удивлением смотрят посетители на телевизионную башню. Высокая и тонкая, тянется она к небу. Внутри бетонной башни проложены различные коммуникации, а скоростной лифт перевозит людей и материалы. Вверху на головокружительной высоте башня расширяется. Здесь инженеры разместили аппаратуру; отсюда же, со смотровых площадок и из окон ресторана, перед посетителями открывается красивая панорама. Не правда ли, чудо техники?!
Однако кто же в наш век, когда мы буквально осыпаны всевозможными техническими достижениями, еще способен удивляться? И все-таки она удивительна, эта новая башня, и поражает нас своей высотой и стройностью. Но действительно ли башня так тонка, как кажется?
Посмотрим в рекламных проспектах, каковы высота и диаметр башни, и вычислим их отношение. Оно равно примерно 18, т. е. высота башни в 18 раз больше ее ширины. Много ли это?
Для сравнения вспомним тростник, который достигает в высоту 2 м, имея в диаметре лишь 1 см.
 Отношение высоты h к среднему диаметру d у телевизионной башни значительно меньше, чем у стебля. Свидетельствует ли это о превосходстве биологической организации?
Отношение высоты h к среднему диаметру d у телевизионной башни значительно меньше, чем у стебля. Свидетельствует ли это о превосходстве биологической организации?
Следовательно, у тростника отношение длины стебля к его диаметру равно 200. А ведь стебель травы может быть еще тоньше! Выходит, телевизионная башня не "чудо техники", а всего лишь несовершенная копия того, что уже давно создано природой? Но не будем спешить с заключениями.
Живые организмы за миллионы лет эволюции действительно нашли такие решения различных технических задач, что у них могли бы поучиться и уже учатся инженеры. Внедрением подобных решений в технику занимается теперь специальная наука. Ее назвали бионикой, соединив таким образом воедино два слова: "биология" и "техника". Использовать в технике то, что уже создала природа, — такая задача стоит сейчас перед многочисленными группами исследователей во всем мире. На счету специалистов по бионике уже есть некоторые успехи. К их числу, несомненно, относится описанная нами телевизионная башня. И все же не будем торопиться с выводами, поскольку здесь сразу возникает множество разнообразных вопросов, которые ждут ответа. Прежде чем пытаться ответить на них, вновь обратимся к примерам.
 Мухи-журчалки могут неподвижно висеть в воздухе и мгновенно исчезать, с большой скоростью срываясь с места
Мухи-журчалки могут неподвижно висеть в воздухе и мгновенно исчезать, с большой скоростью срываясь с места
Наверное, каждый из нас, гуляя в спокойный и теплый летний день за городом, замечал насекомых, которые, подобно вертолету, неподвижно висят в воздухе. Но когда мы, желая их получше рассмотреть, приближаемся к ним, они мгновенно исчезают. Мы удивленно озираемся и обнаруживаем, что они мерцают уже где-то в метре от нас. Это мухи-журчалки, названные зоологами сирфидами (
Syrphidae). Журчалки могут недвижно висеть на одном месте, а при малейшей опасности тотчас его менять. Они летят по прямой да так быстро, что мы даже не в состоянии проследить за ними глазами: мы лишь замечаем, что насекомое, только что бывшее здесь, уже находится в Другом месте и снова, точно вертолет, висит в воздухе.
Предположим, муха меняет свое местоположение всего за какую-нибудь десятую долю секунды, и за это время она пролетает около метра. Это значит, что ее скорость равна 10 м/с. (Вероятно, эта цифра несколько приуменьшена, но сейчас для нас это не существенно.) Следовательно, муха пролетает за секунду расстояние, в 1000 раз большее длины ее тела, которая составляет примерно 1 см.
А теперь представим себе самолет, длина корпуса которого, скажем, около 30 м. Если бы он мог покрыть за секунду расстояние, в 1000 раз превышающее эту длину, то его скорость должна была бы равняться 30 км/с, что в 100 раз больше скорости звука. В таком случае наш воображаемый самолет обогнал бы любую ракету. Но чтобы выдержать сравнение с мухой-журчалкой, он одновременно должен был бы обладать способностью неподвижно висеть в воздухе, подобно вертолету.
Этот пример еще поразительнее, чем сравнение былинки с телевизионной башней. Неужели действительно биологические системы настолько совершеннее технических? Уж не правы ли виталисты, говорящие о существовании некой "чудодейственной силы" (
vis vitalis), которая присуща биологическим системам и непостижима для естественных наук?
Таких примеров сколько угодно. Читатель мог бы понаблюдать за проворной жужелицей и сравнить ее с автомобилем, подобно тому как мы сравнивали муху и самолет, или вспомнить водомерок, стремительно бегущих по поверхности пруда. Все эти сравнения удивительны. Как их понять?
Однако продолжим наши расчеты. Итак, муха пролетает за секунду расстояние, в 1000 раз большее длины ее тела. Поистине природа творит чудеса! А как быстро летают птицы? Способны ли они достичь такой же скорости?
 Жужелица — один из самых проворных 'бегунов'. Сравнится ли с ней гоночный автомобиль, если сопоставить для них отношения скорости движения к длине (v/l)? Есть ли смысл в таком сравнении?
Жужелица — один из самых проворных 'бегунов'. Сравнится ли с ней гоночный автомобиль, если сопоставить для них отношения скорости движения к длине (v/l)? Есть ли смысл в таком сравнении?
Один из самых быстрых наших "летунов" — ласточка. Длина ее тела около 10 см, следовательно, если использовать прежние соотношения, ее скорость должна быть 100 м/с, или 360 км/ч. Ласточка действительно летает очень быстро, но все же ее скорость по крайней мере в 4 раза меньше рассчитанного нами значения.
Что же тогда можно сказать о Скорости полета крупных птиц: лебедя, орла или аиста?
Если бы мы продолжили подобные сопоставления, то пришли бы к ошеломляющим результатам: слоны с огромной скоростью мчались бы по саваннам; кенгуру, соревнуясь с блохами, совершали бы километровые прыжки. А могли ли бы мы, подобно водомеркам, бежать по поверхности воды, если бы у нас было много ног? Конечно, нет! Мы должны признать, что простое сравнение, учитывающее только пропорции, себя не оправдывает. Почему? Ведь правило углов справедливо для треугольников любых размеров, и законы геометрии применимы как для расчета модели атома, так и для определения расстояния между Землей и Луной. Все это действительно так, но в науке следует остерегаться скороспелых обобщений.
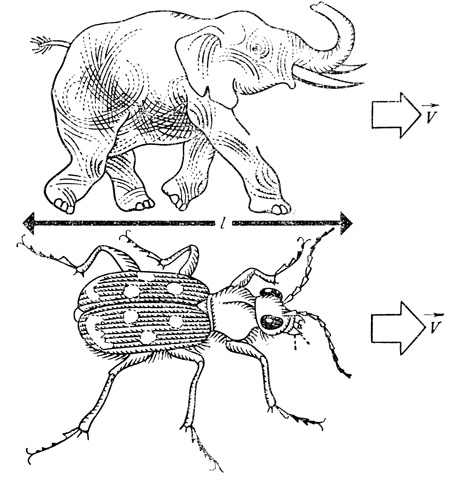 Если природа творит чудеса, почему же слон не бежит быстрее? С какой скоростью он должен мчаться, чтобы отношение v/l было у него таким же, как у жужелицы. По-видимому, мы что-то не то сравниваем
Если природа творит чудеса, почему же слон не бежит быстрее? С какой скоростью он должен мчаться, чтобы отношение v/l было у него таким же, как у жужелицы. По-видимому, мы что-то не то сравниваем
Любая домашняя хозяйка по собственному опыту знает, что килограмм крупной картошки можно очистить быстрее, чем килограмм мелкой. Как известно из математики, поверхность шара увеличивается пропорционально квадрату его диаметра, а объем шара связан с диаметром кубической зависимостью, и потому в килограмме мелкой картошки кожуры больше, чем в килограмме крупной. Даже такой несложный геометрический пример показывает, что в расчетах не всегда можно исходить из простой пропорциональности. Инженерам это давно известно, и какой-нибудь сведущий в технике читатель уже на первом примере сморщил бы нос: "Телевизионную башню, тонкую как стебелек, я бы мог построить, но пусть она будет не выше травинки". Или: "Почему же в природе трава не вырастает до 200 м?" Последний вопрос заставляет о многом задуматься, и мы еще не раз к нему вернемся.
Каждый мальчуган, который когда-либо строил модель самолета, знает, что ее можно смастерить двумя способами. Можно построить уменьшенную копию настоящего большого самолета — серебристую птицу с двигателями, окошечками кабины и другими деталями. Однако подобная модель годится только для того, чтобы повесить ее над письменным столом, и, конечно, не следует ожидать, что она сможет летать. Если же мы хотим иметь летающую модель такого же размера, ее надо делать иначе, и в первую очередь следует изменить размеры и профиль крыла. В результате модель будет мало похожа на настоящий самолет.
За этим примером стоят серьезные проблемы техники и биологии.
Начнем с техники. Здесь на основе анализа сравнительно простых систем удалось выявить важные теоретические закономерности, которые использует и развивает сейчас биофизика.
Остановимся на авиации. Чтобы проверить расчеты конструкций и при необходимости исправить их, инженеры испытывают модели новых самолетов в аэродинамической трубе. При этом в большинстве случаев поневоле приходится обращаться к уменьшенным копиям, а для того чтобы результаты модельных испытаний можно было использовать на практике, ученые разработали теорию подобия.
Очень скоро выяснилось, что некоторые величины характеризуют различные движущиеся тела и позволяют сравнивать их между собой гораздо лучше, чем использованные нами ранее коэффициенты пропорциональности. Примером такой величины может служить так называемое число Рейнольдса (Re), которое играет огромную роль в авиации и судостроении; его рассчитывают по следующей формуле:

 Форма потока, обтекающего шар, при различных числах Рейнольдса (Re). Поведение потока определяется не размерами тела, а только числом Рейнольдса, которое, правда, зависит от размеров
Форма потока, обтекающего шар, при различных числах Рейнольдса (Re). Поведение потока определяется не размерами тела, а только числом Рейнольдса, которое, правда, зависит от размеров
Кинематический коэффициент вязкости — это параметр, характеризующий "густоту" среды. Мы не будем подробно на нем останавливаться, а лишь отметим, что, если выразить скорость и длину в метрах и секундах, то кинематический коэффициент вязкости равен для воды 1,06⋅10
-6, а для воздуха — 14,9⋅10
-6.
Практический смысл числа Рейнольдса заключается в следующем: поведение потока жидкости или газа, обтекающего тело определенной формы при постоянном значении числа Рейнольдса, не зависит от размеров тела.
В качестве примера рассмотрим движущийся шар. Независимо от того, большой он или маленький, при числе Рейнольдса меньше 1000 воздух, вода или любая другая среда обтекают шар плавно, или, как говорят в гидродинамике, ламинарно. Как только число Рейнольдса превысит критическое значение (вследствие увеличения диаметра шара или скорости потока), сразу же появятся завихрения. Таким образом, если мы хотим определить аэродинамические свойства крыла самолета по поведению в аэродинамической трубе его уменьшенной модели, нам надо сначала определить число Рейнольдса для крыла самолета, исходя из реальных размеров и скорости последнего. Затем, зная размеры модели, следует установить такую скорость воздуха в трубе, при которой числа Рейнольдса для модели и настоящего самолета одинаковы.
 Биологический объект в аэродинамической трубе. Такие устройства позволяют изучать поведение воздушного потока при обтекании летающих объектов
Биологический объект в аэродинамической трубе. Такие устройства позволяют изучать поведение воздушного потока при обтекании летающих объектов
Специалисты по бионике рассчитали значения числа Рейнольдса для многих животных. Так, для ласточки — ее скорость полета 10 м/с и длина тела 0,01 м — мы получим
Re = (10 ⋅ 0,01) / (14,9 ⋅ 10
-6) = 6700
Подобное значение числа Рейнольдса столь мало, что оно вряд ли может заинтересовать авиаконструктора. Если мы подставим в приведенную выше формулу значения скорости и размеров современного самолета, то сразу поймем, почему интерес авиаконструктора вызывают лишь шести- или восьмизначные числа. Как видно из рисунка, такие значения числа Рейнольдса (1 000 000 и выше) характерны лишь для дельфинов — наиболее крупных и быстрых пловцов.
Итак, даже технические системы, гораздо менее сложные, чем системы в живой природе, бессмысленно сравнивать только на основании пропорций. Сравнение систем одинаковой формы, но отличающихся друг от друга размерами можно проводить, опираясь лишь на безразмерные величины, определяемые на основе различных параметров систем. На сегодняшний день известны и применяются около ста таких безразмерных величин.
Но довольно техники. Мы познакомились в общих чертах с теорией подобия в технике, и, может быть, этим ограничиться? Действует ли теория подобия в биологии или это всего лишь интересная игра?
Конечно, теория подобия не есть основное направление исследований в биофизике, но с ней связаны многие проблемы, представляющие общебиологический интерес. Многое нам кажется совершенно очевидным и не вызывает никаких вопросов. Мы часто говорим: это так, потому что иначе и быть не может! Однако детей такой ответ обычно не удовлетворяет. Они терзают нас своими "почему?". Эта детская черта отличает и многих исследователей. Не одно крупное открытие было бы еще сделано, если бы человек не боялся спрашивать о тривиальном и удивляться вещам, ставшим для других повседневными.
 Значения числа Рейнольдса для разных животных различаются на много порядков
Значения числа Рейнольдса для разных животных различаются на много порядков
Так не будем бояться спрашивать об обычном; почему мышь не может быть меньше, а слон — больше, чем они есть, почему кенгуру не может прыгать еще выше, почему деревья не растут до неба?
Мы узнали, что бессмысленно связывать "максимальные возможности" объектов с их размерами. Но почему? Лучше всего ответить на это на примере вопроса, уже поставленного выше: почему мы не можем бегать по воде, как водомерки? За ответом далеко ходить не надо: мы слишком тяжелы, и поверхностное натяжение воды нас не удержит. Можно возразить, что площадь наших ступней намного больше поверхности, которую занимают шесть лапок насекомого. При необходимости мы можем даже стать на водные лыжи. Но известно, что "водный лыжник" способен скользить по воде, лишь прицепившись к быстродвижущемуся катеру. И в этом случае его держит на воде не поверхностное натяжение, т. е. гидростатическая сила, а сила гидродинамическая.
Но одно, во всяком случае, несомненно: свойства материала не зависят от величины объекта, поэтому поверхностное натяжение воды совершенно одинаково как для водомерки, так и для человека. Однако человек создает слишком большую нагрузку на поверхность и "проваливается".
Чтобы рассмотреть этот вопрос подробнее, обратимся к математике. Здесь нам понадобятся два символа: знак l — характеристический размер тела, например его длина или диаметр, и знак ∼, обозначающий пропорциональность. Две величины являются пропорциональными, если, например, удвоение одной из них влечет за собой удвоение другой. Очевидно, совсем не обязательно знать, сколько стоит килограмм картошки, чтобы утверждать, что два килограмма ее стоят вдвое дороже, чем один. Цена картошки пропорциональна ее весу. Выше, когда мы сравнивали чистку больших и маленьких клубней картошки, мы упомянули об отношении поверхности клубней и их объема к диаметру. Утверждение: поверхность возрастает пропорционально квадрату диаметра — можно записать следующим образом:
поверхность ∼ l
2.
 Площадь поверхности куба с длиной ребра l равна 6⋅l2, а его объем составляет l3. Таким образом, площадь поверхности пропорциональна l2, а объем пропорционален l3. Эти соотношения справедливы для тел разной формы
Площадь поверхности куба с длиной ребра l равна 6⋅l2, а его объем составляет l3. Таким образом, площадь поверхности пропорциональна l2, а объем пропорционален l3. Эти соотношения справедливы для тел разной формы
Теперь мы хотим узнать, какую картошку можно очистить быстрее. Предположим, что время чистки прямо пропорционально поверхности, т. е. для получения одного квадратного метра кожуры нужно в обоих случаях затратить одно и то же время. Тогда мы можем написать

Таким образом, производительность работы пропорциональна величине 1/l. Отберите картошку вдвое крупнее, и на ее очистку потребуется вдвое меньше времени!
Однако наша книга — не пособие для домашних хозяек, поэтому обратимся снова к биологии и вспомним водомерок. И в этом случае метод расчета остается тем же. Тело лежит на воде, но касается воды только некоторая "определенная" часть его поверхности, а точнее поверхность лапок насекомого. Слово "определенная" можно выразить посредством коэффициента пропорциональности, а так как величина этого коэффициента нас не интересует, мы можем спокойно написать:
поверхность опоры ~ l
2.
Нагрузка на поверхность определяется как вес, деленный на площадь опоры. Вес тела пропорционален объему тела, т. е. l
3, а площадь опоры пропорциональна поверхности тела, или l
2. Следовательно, можно написать

т. е. нагрузка на поверхность растет пропорционально длине тела. А теперь не спеша подсчитаем: человек в 200 раз длиннее водомерки, и, чтобы мы могли бегать по воде, нам необходима вода с 200-кратным поверхностным натяжением или водные лыжи с поверхностью порядка 10 квадратных метров, к тому же не обладающие собственным весом. Таким образом, даже водомерка, пропорционально увеличенная, не смогла бы бегать по воде.
 Давление, оказываемое водомеркой на поверхность воды, равно отношению ее веса к площади занимаемой ею поверхности и пропорционально ее длине l
Давление, оказываемое водомеркой на поверхность воды, равно отношению ее веса к площади занимаемой ею поверхности и пропорционально ее длине l
Можно провести множество подобных расчетов и показать, почему биологические объекты не могут иметь безгранично большие размеры. По всей вероятности, максимальная нагрузка, которую способны выдержать наши кости и мышцы, должна определяться свойствами образующих их тканей. Это относится и к стволам деревьев. Нагрузка на поверхность здесь также играет решающую роль, и, как мы видели, она увеличивается пропорционально длине ствола. Мышь, будучи пропорционально увеличенной до размеров слона, просто переломилась бы. Только мощно укрепленный скелет исполинов придает им неуклюжую устойчивость. Доисторические ящеры достигли, по-видимому, максимальных размеров — собственный вес погубил их.
 Так как давление прямо пропорционально величине тела, кости ног у бегемота (а) нагружены гораздо больше, чем у мелкого млекопитающего — лемминга (б). У бегемота такая нагрузка компенсируется малым отношением высоты ноги к ее диаметру
Так как давление прямо пропорционально величине тела, кости ног у бегемота (а) нагружены гораздо больше, чем у мелкого млекопитающего — лемминга (б). У бегемота такая нагрузка компенсируется малым отношением высоты ноги к ее диаметру
А почему мышь не может быть меньше, чем она есть в действительности? На этот вопрос мы ответим позже, потому что он не связан с проблемами механики, которые интересуют нас в первую очередь.
 Внешним скелетом обладают только мелкие животные. В то время как насекомому (А — долгоносик) хитиновый панцирь обеспечивает защиту и опору, черепахе панцирь служит лишь для защиты от врагов; опорную оке функцию выполняет внутренний скелет. К аналогичным выводам мы приходим на основании теории подобия
Внешним скелетом обладают только мелкие животные. В то время как насекомому (А — долгоносик) хитиновый панцирь обеспечивает защиту и опору, черепахе панцирь служит лишь для защиты от врагов; опорную оке функцию выполняет внутренний скелет. К аналогичным выводам мы приходим на основании теории подобия
Однако с помощью расчетов, аналогичных проделанным, можно показать, почему из мухи нельзя сделать слона. Если бы млекопитающие, подобно насекомым, имели внешний скелет, то их панцирь был бы невыносимо тяжелым. Таким образом, и в этом случае принцип строения определяется свойствами материала. Твердый панцирь крупных животных, таких, как черепахи и броненосцы, уже не является элементом скелета, поддерживающим тело, а служит лишь дополнительной защитой.
Теперь попытаемся с помощью теории подобия решить задачу о прыжках кенгуру. Для этого нам необходимо знать кое-что из физики. Желая прыгнуть, мы сгибаем ноги в коленях, затем быстро их выпрямляем (при этом центр тяжести тела перемещается на высоту f ), сообщая телу кинетическую энергию, которая позволяет нам на короткое время преодолеть силу земного тяготения и взлететь вверх (центр тяжести перемещается на высоту h). Создаваемая силой отталкивания кинетическая энергия переводит тело в состояние с более высокой потенциальной энергией. Это можно записать в виде следующего уравнения:
вес ⋅ h — сила отталкивания ⋅ f.
Отсюда вычислим высоту прыжка h:

Сила отталкивания пропорциональна размеру поперечного сечения мышцы, который пропорционален квадрату длины мышцы, а последняя в свою очередь пропорциональна длине тела l. Таким образом, мы можем написать: сила отталкивания ~ l
2. Величина f пропорциональна длине тела l, а вес ~ l
3. Подставив эти значения в выражение для h, получим нечто удивительное:

Итак, высота прыжка h вообще не зависит от длины тела, или, как сказал бы математик, она не является функцией длины тела! Это означает, что животные, имеющие сходное строение, должны прыгать на одинаковую высоту независимо от размеров тела! Слон прыгает на такую же высоту, как и мышь, причем речь идет не о пропорциональной зависимости высоты от размеров тела, а об абсолютном ее значении!
Конечно, за физикой не следует забывать биологию. Природа предоставляет живым организмам широкие возможности для адаптации. У прыгающих животных особенно сильно развиты задние конечности, и поэтому эти животные прыгают выше других. Не следует забывать, что наши расчеты дают лишь грубую оценку. Но сделанный на их основании вывод имеет большое значение. Теперь мы знаем, почему высота прыжка не должна быть пропорциональна величине тела. Не будем забывать: одни лишь математические соотношения без учета физических свойств материала не дают реальной картины.
 Биомеханика прыжка. Перемещая центр тяжести тела на отрезок f, прыгун сообщает телу ускорение, и тогда его центр тяжести поднимается на высоту h
Биомеханика прыжка. Перемещая центр тяжести тела на отрезок f, прыгун сообщает телу ускорение, и тогда его центр тяжести поднимается на высоту h
Проведенные нами расчеты и оценки основаны на положениях механики, а именно статики и кинематики. Подобные расчеты часто используют, для анализа различных форм движения и условий стабильности в мире животных. Аналогичным образом решают и некоторые задачи аэро — и гидродинамики. С помощью таких расчетов можно получить параметры подобия, например число Рейнольдса, о котором мы уже говорили. Мы коснулись здесь раздела науки, называемого биомеханикой.
Наши рассуждения показали, что простое сравнение былинки и телевизионной башни, жужелицы и гоночного автомобиля недопустимо. Но даже если привлечь теорию подобия и проводить сравнения с учетом физически обоснованных параметров, "способности" живой природы не перестают поражать воображение. Природа дает богатую пищу для развития бионики. Конструкции многих изящных архитектурных сооружений заимствованы у растений, а конструкции самолетов — у птиц, рыб или водных млекопитающих. Как видим, в бионике кроется богатый источник технических знаний и для биологов.
Однако вопросы, касающиеся размеров биологических объектов, далеко не всегда можно решить с помощью механики. От чего зависит ограничение размеров снизу? Почему самое мелкое млекопитающее не меньше землеройки? Почему нет мышки величиной с жука? Эти вопросы поначалу кажутся тривиальными и бессмысленными.
Механика здесь нам помочь не в силах, поэтому мы попытаемся глубже проникнуть в сущность биологических процессов. Это еще один шаг на пути, который постепенно приведет нас от внешнего вида организма и его строения к физиологии организма и клетки и, наконец, в последней главе книги — к молекулам.
Что собственно отличает млекопитающих, а также птиц от других животных? Они достигли высшей ступени эволюции благодаря приобретению, которое обеспечило им преимущества во многих жизненно важных процессах, и таким приобретением явилась постоянная температура тела. Их называют поэтому "теплокровными" (гомойотермными) животными в отличие от "холоднокровных" (пойкилотермных), температура тела которых обычно лишь немного выше температуры окружающей среды и понижается вместе с ней.
Для обеспечения постоянной температуры необходима достаточно сложная система терморегуляции. Какое же преимущество дает она животному?.
Мы знаем, что температура тела теплокровных животных значительно выше температуры окружающей среды. Температура человека (37 °С) лежит примерно в середине интервала температур, присущих теплокровным. При высокой температуре химические процессы ускоряются. Например, сахар растворяется в чае быстрее, если чай горячий. И конечно же, при 37 ° С биологическая система гораздо работоспособнее, чем при 10 или 20 °С. Оправдывает ли само по себе повышение температуры затраты на термостатирование у теплокровных животных? Ответить на этот вопрос достаточно сложно. Возможно, что температура тела выше температуры окружающей среды просто потому, что в этом случае легче осуществить термостатирование тела, т. е. поддержание в нем постоянной температуры. Нагрев всегда осуществить проще, чем охлаждение, а термостат с более высокой по сравнению с окружением температурой охлаждается сам.
Главное заключается даже не в том, что температура организма выше, чем температура окружающей среды, а в том, что она постоянна. Мы знаем, как чувствительны к колебаниям температуры регулирующие устройства и счетные приборы. Электрическое сопротивление изменяется в зависимости от температуры, и некоторые электронные схемы при колебаниях температуры становятся нестабильными и более чувствительными к помехам. Процессы нервной деятельности также подвержены сильному влиянию температуры. Известно, как неприятно действуют колебания температуры на работу мозга. Постоянство температуры нашего тела, имеющего много сложных систем регуляции, и в особенности постоянство температуры человеческого мозга, следует считать важнейшим условием существования высокоорганизованной жизни. Учет этого физиологического параметра вызывает новые вопросы. Как действует тепло? Как оно возникает, преобразуется, передается? Эти вопросы относятся к компетенции огромной и в высшей степени важной области биофизики, на которой в дальнейшем мы остановимся подробнее. Здесь же мы изложим лишь некоторые соображения, которые помогут нам решить вопрос о нижней границе размеров тела млекопитающих или вообще теплокровных животных.
Каждый, кого хоть раз бросало в пот, испытал на себе процесс образования тепла в теле. Любое превращение энергии в теле, будь то мышечная деятельность или пищеварение, дает тепло. Если такого тепла неоткуда "взять", мы производим его при необходимости собственными силами, например начиная дрожать от холода. При этом, как принято говорить, мы "сжигаем" калории.
Так наш "термостат" нагревается. Он должен работать постоянно, потому что тепло непрерывно уходит в более холодную окружающую среду. Если же в теле вдруг возникает излишек тепла, мы потеем, то есть включаем механизм охлаждения.
Однако какое отношение имеет тепловой баланс организма к размерам мыши"? Самое прямое! Здесь мы также можем вывести уравнение баланса подобно тому, как получали различные соотношения, рассматривая примеры из биомеханики. Предположим, что количество тепла, образующегося в единице массы тела, одинаково для всех животных. Один грамм тела мыши производит в единицу времени столько же тепла, сколько производит один грамм тела человека или слона. Вес животного пропорционален его объему, а последний, как мы видели, пропорционален линейному размеру тела в третьей степени, т. е. величине l
3. Теперь посмотрим, каким образом тело отдает тепло окружающей среде. У всех теплокровных животных температура тела приблизительно одинакова. И мы можем смело постулировать, что теплоотдача увеличивается пропорционально поверхности тела, т. е. квадрату его размера, или величине l
2. Тогда соотношение между теплоотдачей и образованием тепла в теле выглядит так:
(теплоотдача ∼ l
2) / (образование тепла ∼ l
3) = 1/l
О чем говорит полученное отношение? Чем от больше, тем больше теплоотдача, и чем оно меньше, тем больше тепла образуется в теле животного.
В
первом случае животное с трудом обогревает себя, во втором — легко потеет. В действительности это соотношение ограничено и сверху и снизу, т. е. оно может изменяться лишь в границах, которые определяются возможностью существования теплокровных животных. Верхняя граница размеров не представляет интереса, потому что с точки зрения теплоотдачи величина l может быть сколь угодно большой и размеры слона определяются не температурой, а только механическими свойствами костей и мышц. Другое дело мышь. В этом случае значение l так мало, что отношение .1/l почти достигает максимума. Соотношение между массой и поверхностью тела у мыши настолько неблагоприятно, что необходимая температура тела поддерживается лишь благодаря интенсивному обмену веществ.
Землеройка, питающаяся насекомыми, — самое маленькое из теплокровных животных. У теплокровных еще меньшего размера соотношение между, теплоотдачей и образованием тепла было бы таково, что они не могли бы существовать.
У водных млекопитающих критическое значение этого соотношения еще выше: даже при наличии изолирующего жирового слоя теплоотдача тела в воде больше, чем в воздухе. Поэтому самые маленькие теплокровные, обитающие в воде, намного крупнее своих сородичей, живущих на суше.
В своих расчетах мы исходили из предположения, что образование тепла в теле пропорционально его массе. Но это лишь очень грубое приближение. Флегматичность слона не сравнима с подвижностью мыши. Природа при помощи различных приспособлений "добилась" понижения критического значения отношения 1/l. Так как образование тепла у млекопитающих в общем связано с энзиматически управляемым процессом сгорания, то его мерой вполне может служить потребление кислорода. Это наглядно представлено на рисунке: чем меньше животное, тем интенсивнее у него протекает обмен веществ. Аналогичным образом регулируется частота дыхания и сердцебиения.
Итак, мы установили, что размеры, внешний облик и поведение животных в значительной мере определяются не только механическими закономерностями, но и такими процессами, как передача тепла.
 Количество тепла, вырабатываемое землеройкой, пропорционально ее объему и, следовательно, величине l3. Теплоотдача пропорциональна поверхности тела землеройки, т. е. l2. Отношение обеих величин l2/l3 = 1/l определяет наименьшую длину тела теплокровного животного
Количество тепла, вырабатываемое землеройкой, пропорционально ее объему и, следовательно, величине l3. Теплоотдача пропорциональна поверхности тела землеройки, т. е. l2. Отношение обеих величин l2/l3 = 1/l определяет наименьшую длину тела теплокровного животного
Мы коснулись области энергетики, или, как говорят физики, термодинамики. Позже мы увидим, насколько важна эта область для понимания различных биологических процессов. С приведенным примером тесно связан вопрос: как образуется тепло? Действуют ли мышцы подобно тепловой машине, которая превращает тепло в механическую работу? Этот вопрос не связан прямо с темой данной главы, и мы ответим на него позднее. Сейчас же нас интересуют сравнение размеров различных живых организмов и их ограничения как сверху, так и снизу.
Постановку вопроса об обмене энергией следует рассматривать как еще один шаг, приближающий нас к реальному биологическому объекту. Конечно, мы не можем продвинуться в этом направлении далеко: для этого понадобилось бы множество чисто математических расчетов. Мы остановимся лишь на некоторых аспектах проблемы.
Для осуществления обмена веществ в клетках и тканях необходимо, чтобы к ним поступал кислород, а из них удалялась выделяющаяся там углекислота.
 Тепловой баланс обеспечивается с помощью сложной системы регуляции. Температура тела как функция его теплоты определяется разницей между производимым теплом и теплоотдачей
Тепловой баланс обеспечивается с помощью сложной системы регуляции. Температура тела как функция его теплоты определяется разницей между производимым теплом и теплоотдачей
У насекомых воздушные трубки, так называемые трахеи, подходят непосредственно к органам, где они разветвляются; таким образом обеспечивается газообмен. Этот принцип оставляет желать лучшего. В таких трубках газообмен протекает вяло и воздух проходит только на малые расстояния — это одна из причин ограничения размеров насекомых сверху. Позвоночные животные обладают жидкостной системой газообмена, а именно системой кровообращения между легкими и тканями. Такая сложная система обеспечивает снабжение кислородом более крупных органов. Частоту сердцебиения у животных различной величины можно объяснить на основе анализа системы кровообращения.
Подобным же образом можно проанализировать форму и размеры растений. Только по их "артериям" транспортируется не растворенный кислород, а вода, которую корни растений забирают из почвы.
Приведем пример также из области нейрофизиологии. Нервный импульс распространяется значительно медленнее, чем передаются знаки азбуки Морзе по телеграфному проводу. Вовремя ли заметит удав, что ему наступили на хвост? Это конечно, шутка, но у крупных животных проблема связи действительно существует.
Совершенно новая глава биофизики открывается вопросом о предельных минимальных размерах биологических объектов. Почему бактерии не стали еще мельче? Почему клетки тканей именно такой величины, а не меньше? И здесь нам на помощь придут законы физики, но уже не "макрофизики", которая имеет дело с видимыми, весомыми, непосредственно измеряемыми телами, а "микрофизики", или статистической физики малых частиц. В дальнейшем мы коснемся этого раздела науки, а пока снова обратимся к макрообъектам.
 Мышь вдыхает за минуту воздух, объем которого больше объема ее тела. У кошки объем воздуха, вдыхаемого за то же время, составляет 1/10, у козы — 2/100 и у лошади — 6/1000 объема тела. Так энергетические соотношения, рассчитанные с помощью теории подобия, находят отражение в физиологии
Мышь вдыхает за минуту воздух, объем которого больше объема ее тела. У кошки объем воздуха, вдыхаемого за то же время, составляет 1/10, у козы — 2/100 и у лошади — 6/1000 объема тела. Так энергетические соотношения, рассчитанные с помощью теории подобия, находят отражение в физиологии
Итак, последний вопрос: насколько интересны и важны обсуждавшиеся здесь проблемы для самой биологии, для объяснения различных принципов организации в природе и ограничивается ли этим их значение? Даже если бы это было действительно так и наши попытки разобраться в подобных проблемах служили бы "только" расширению нашего кругозора, утоляли нашу неистощимую жажду познания, даже тогда они были бы оправданны!
В действительности же за каждой такой проблемой стоят чисто практические интересы. О бионике мы уже говорили. Можно привести и другие примеры. Наша химическая промышленность ежедневно выпускает новые соединения, фармакологическое действие которых нужно тщательно изучить. Эти соединения испытывают на лабораторных животных и на основании результатов опытов делают заключение о действии препаратов на человека. Дозы обычно определяют, исходя из веса тела. Верно ли это? Нет ли более подходящего параметра сравнения? Вспомним также об использовании экспериментальных животных в космических исследованиях. Как, исходя из действия изменившейся гравитации на мышь, собаку или обезьяну, судить о влиянии этого фактора на людей? Не следует забывать и о том, что люди тоже отличаются друг от друга размерами и пропорциями. Как, например, сравнивать физиологические показатели или спортивные достижения детей и взрослых? Как действует ожирение на работу сердца? Перечень подобных вопросов можно продолжить, и это говорит о практической важности поставленных выше проблем, которые для наглядности мы рассмотрели на очень простых примерах.
А теперь подведем итоги. В начале этой главы мы сравнивали технические и биологические объекты. Это вызвало у нас, казалось бы, довольно будничные вопросы. В поисках ответа на них мы привлекали на помощь физику, а также отчасти механику и математику. По мере углубления в эти проблемы мы узнали, что физические законы действуют в живой природе независимо от уровня биологической организации. И даже если нам не всегда удавалось свести биологические закономерности непосредственно к основным физическим законам, противоречия между ними не обнаруживалось. Основная задача этой главы — поставить как можно больше вопросов. Дальше мы будем постепенно переходить от видимого к невидимому, от большого к малому, обращая особое внимание на связь между формой объекта и его назначением, структурой и функцией.
Форма живых организмов в системе координат
Можно ли с помощью чисел описать форму живых организмов? Маргаритка — облачко точек в n-мерном фазовом пространстве. ЭВМ хранит информацию о многообразии форм и рассчитывает процессы эволюции. Изменчивость и отбор — процесс оптимизации жизни; как это можно использовать? Можно ли подсчитать целесообразность? Анализ оптимальности — что, как и сообразно с чем оптимизируется? Оптимальность и изменчивость форм — основа биологического многообразия.
В предыдущей главе мы говорили о величинах и измерениях. Теперь мы обратим свое внимание на форму и внешний вид растений и животных. Пригодятся ли нам и здесь математика и физика?
Анализ проблемы формы в биологии приводит нас к вопросам эстетики. Достаточно вспомнить, например, книгу Эрнста Геккеля "Красота форм в природе". Однако мы не будем заниматься эстетикой, а останемся на твердой почве естествознания. Тем не менее факт остается фактом — эстетика и проблемы формы в биологии тесно связаны. Эстетика и целесообразность в живой природе, очевидно, взаимозависимы. В повседневной жизни целесообразное отнюдь не всегда можно назвать "прекрасным". В технике целесообразность и красоту часто даже рассматривают как две противоположности. Биологическим же объектам свойственна совершенно особая целесообразность, и она представляется нам прекрасной.
Форма и целесообразность в биологии связаны с третьим понятием — приспособленностью. Более столетия назад мир познакомился с работой Чарлза Дарвина "Происхождение видов путем естественного отбора, или сохранение — благоприятствуемых пород в борьбе за жизнь". Эволюционная теория Дарвина является одной из составляющих частей современного научного мировоззрения, и каждому из нас знакомо выражение "борьба за существование". Эволюционное развитие определяется совокупностью двух процессов — вариабельности и селекции, или, проще говоря, случайной изменчивости и отбора.
 Целесообразность той или иной формы в биологии неразрывно связана с красотой. Красивы ли симметричные организмы? Сейчас нам кажется забавным, что Эрнст Геккель в своей книге 'Красота форм в природе' относит к миру прекрасного даже ленточного червя
Целесообразность той или иной формы в биологии неразрывно связана с красотой. Красивы ли симметричные организмы? Сейчас нам кажется забавным, что Эрнст Геккель в своей книге 'Красота форм в природе' относит к миру прекрасного даже ленточного червя
Изменчивость и отбор обусловливают процесс приспособления, а приспособление есть не что иное, как повышение целесообразности в соответствии с определенным образом жизни. Ласточка целесообразна в качестве изящного летуна, приспособленного к поиску пищи в воздухе; лебедь и нырок приспособлены для жизни в тихих водах. Таким образом, целесообразность не есть нечто абсолютное: она проявляется лишь во взаимоотношениях между организмом и окружающей его средой.
Можно ли "подсчитать" целесообразность? Вероятно, привлечение математики к такому сугубо биологическому вопросу вызовет недоумение. Но так ли это бессмысленно? Разве человек не старается всегда принимать в расчет вопросы целесообразности? Во многом, что касается нашей повседневной жизни, целесообразность представляется нам само собой разумеющейся; мы с недоумением качаем головой, когда некоторые хитрецы пытаются доказывать то, что кажется нам совершенно очевидным. Еще Евклид говорил в своей геометрии: самый короткий путь между двумя точками — прямая.
 Рыбы (плавающие организмы) имеют различные формы. Какая из них наиболее целесообразна? Ответить на этот вопрос нельзя без учета образа жизни данного вида. Можно ли рассчитать целесообразность? А — линь; Б — красная крылатка; В — сом; Г — камбала
Рыбы (плавающие организмы) имеют различные формы. Какая из них наиболее целесообразна? Ответить на этот вопрос нельзя без учета образа жизни данного вида. Можно ли рассчитать целесообразность? А — линь; Б — красная крылатка; В — сом; Г — камбала
Эмпирический факт, тривиальность! Но доказать его математически довольно сложно. И вероятно, вначале попытка доказать это давно известное положение казалась не чем иным, как проявлением излишней педантичности. Но очень скоро выяснилось, что найденный метод позволяет доказать и нечто другое, ранее не известное. В результате в математике возникло новое направление: теория оптимальных процессов. Это довольно сложная область математики, первоначально разработанная лишь для простейших процессов. В наш век, когда в экономике и технике расчеты осуществляются с помощью электронных вычислительных устройств, эта теория приобрела большое значение. Посмотрим, в состоянии ли теория оптимальных процессов объяснять и обосновывать целесообразность в мире организмов.
 Прямая — самая 'целесообразная' линия, если мы хотим найти кратчайший путь между А и Б. Эта древнейшая проблема оптимизации занимала еще Евклида
Прямая — самая 'целесообразная' линия, если мы хотим найти кратчайший путь между А и Б. Эта древнейшая проблема оптимизации занимала еще Евклида
Нельзя забывать, что процессы оптимизации в технике и биологии совершенно различны. Сначала обратимся к технике. Предположим, инженер проектирует мост, который должен быть надежным, легким, дешевым и пропускать в единицу времени определенное количество машин и пешеходов. Эти требования противоречат друг другу. Самый надежный мост всегда тяжелый и дорогой, а самый дешевый — легкий, но ненадежный. Оптимизация заключается в том, чтобы найти такую конструкцию моста, которая, обеспечивая достаточно большую надежность, требовала бы минимум затрат.
У природы возможности несравненно богаче. Она находит оптимальные решения не путем предварительного расчета, а через изменение и селекцию, т. е. отбор. Сначала появляется большое количество различных вариантов, но из них остаются только те, которые выдержали борьбу за существование. Неподходящие образцы "отбрасываются". В действительности это как бы движение на ощупь, медленное изменение того или иного свойства в том или ином направлении. Оптимизация биологической структуры возможна только тогда, когда при неизменных условиях в окружающей среде развитие этой структуры совершается достаточно медленно. В противном случае оптимизация невозможна. Таким образом, не каждое приспособление является совершенным, т. е. оптимальным.
Очевидно, такие величины, как вес, стоимость, допустимая нагрузка и т. д., характеризующие конструкцию моста, могут быть выражены количественно и оптимизированы математическими методами. А можно ли осуществить математическую "обработку" формы живого организма? Да, конечно, только не будем забегать вперед! Такой обработке должна предшествовать определенная "математизация". Иными словами, сначала надо ответить на вопрос: как с помощью цифр описать внешний вид организма?
Для этого есть несколько способов различной степени сложности. Простейший и самый наглядный из них — сравнение числовых характеристик: количества ног, крыльев, глаз, члеников на усиках и т. п. у животных или количества тычинок, цветков, лепестков, компонентов листа и т. п. у растений. Существуют двуногие, шестиногие (насекомые), десятиногие (высшие ракообразные), двукрылые (мухи и комары). Вспомним божьих коровок, их обычные виды — семиточечная (
Coccinella septempunctata) и двухточечная (
Coccinella bipunctata). Живые организмы можно сравнивать и по геометрическим размерам. Так, в справочниках растений мы часто встречаем такое сопоставление: растение маленькое, не больше 10-15 см в высоту, и растение выше 20 см. Можно проводить сравнения и в относительных единицах: например, у лани хвост длиннее уха, а у благородного оленя хвост по длине равен уху. Анализируя форму живых организмов, мы можем осуществлять различные измерения и получать числовые значения, пригодные для дальнейшей математической обработки.
С чего же начать обработку полученных чисел? Математики уже давно научились объединять множество измеряемых величин, или, проще говоря, чисел, таким образом, чтобы получалась некая цельная картина. Если какой-то предмет характеризуется двумя параметрами, то в соответствии с их значениями его можно представить точкой на плоскости. Возьмем пример из биологии: маргаритку, в частности, можно охарактеризовать количеством цветолистиков и высотой соцветия, измеренной в сантиметрах. Обе величины могут изменяться, но только в определенных границах. Итак, мы срываем маргаритку (кому это действие кажется слишком простым, тот пусть попытается отделить соцветие вида Bellis perennis L. от розетки листьев), считаем цветолистики, измеряем длину цветоноса и отмечаем оба значения в системе координат, по осям которой откладываются соответствующие единицы. На том же лугу растет множество других маргариток; мы многократно повторяем наш опыт, и каждый раз на бумаге появляется еще одна точка. При некотором усердии мы получим целое облачко точек.
 Числа (А), размеры (Б) и математические функции (В) — основа для точного сопоставления биологических форм
Числа (А), размеры (Б) и математические функции (В) — основа для точного сопоставления биологических форм
Почему мы выбрали именно эти два параметра? Может быть, они наиболее характерны? Ничего подобного! С таким же успехом мы могли бы взять число листьев в розетке, длину листьев, диаметр головки соцветия и т. д. После того как нам удалось свести два параметра к одной точке, многообразие живой природы снова озадачивает нас, Если к двум параметрам добавить третий, то для каждой конкретной маргаритки мы получим точку уже не на плоскости, а в пространстве, и для всех маргариток, растущих на этом лугу, — объемное облако точек, которое характеризует целый вид. А что дальше?
 Выбранные параметры маргаритки отложены по осям системы координат. В результате измерений этих параметров для различных растений исследуемого вида мы получаем в фазовом пространстве облачко точек, характерное для данного вида. Каждому из растений соответствует одна точка в этом облачке
Выбранные параметры маргаритки отложены по осям системы координат. В результате измерений этих параметров для различных растений исследуемого вида мы получаем в фазовом пространстве облачко точек, характерное для данного вида. Каждому из растений соответствует одна точка в этом облачке
Такой вопрос может возникнуть только у человека, далекого от математики, который убежден в том, что все существующее можно представить зрительно. Математик думает иначе, он живет в мире абстрактного. Если трех измерений не хватает, то пусть будет 4, 5, 6, 10 или, наконец, n измерений, то есть сколь угодно много. И хотя с каждым новым измерением расчеты усложняются, в принципе можно рассчитать любое многомерное пространство. Но теперь слово "пространство" обозначает нечто совершенно отличное от того, что мы обычно изображаем в виде трехмерной структуры. Многомерные пространства используют в своих расчетах также физики-теоретики и называют их фазовыми пространствами. У нас нет причин искать какое-либо другое название. Итак, биологический вид есть облако точек в n-мерном фазовом пространстве.
Математическое обоснование тривиального — что это? Опять игра? Отнюдь нет! То, чем мы здесь занимаемся, относится к области бурно развивающейся сейчас нумерической таксономии. Облако точек вытягивается в длину и разделяется — возникает новый вид. Облако плывет — вид изменяется. Облака находятся близко или далеко друг от друга — это характеризует степень родства различных видов. В принципе все эти процессы можно рассчитать. Но здесь не обойтись, конечно, без быстродействующих электронных вычислительных машин.
Измерение плюс вычисление — это только один из возможных путей, ведущих к пониманию формы живого. Сколько нужно цифр чтобы правильно отобразить форму листа, контуры лягушки или панцирь рака? Конечно, с помощью числового метода мы можем охарактеризовать эти формы с большей или меньшей точностью, в зависимости от числа измеряемых параметров. Но это недостойно истинного математика; к тому же каждое новое измерение увеличивает степень многомерности фазового пространства, что делает расчеты неоправданно сложными. Значительно удобнее выявлять формы не по цифрам, а по аналогии. Это значит, что мы ищем математическую кривую, которая соответствует интересующей нас форме, т. е. аналогична ей, и может быть выражена формулой с возможно меньшим количеством постоянных величин, или констант.
Математикам известна такая универсальная формула, или, точнее, функция, которая позволяет математически выразить почти любую кривую, — это так называемый полином. Он записывается в виде ряда, который можно продолжать сколь угодно долго, но математик ограничивается лишь действительно необходимым числом членов, ибо с каждым новым членом полином все усложняется. Уравнение этого ряда выглядит так:
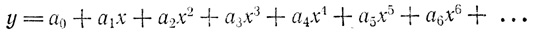
Оно показывает, как изменяется величина у в зависимости от изменения независимо меняющейся величины
x. Обычно говорят, что y есть функция от
x. Если значения
x и
у откладывать по осям системы координат, то мы получим кривую. Буквы
a0,
a1,
a2,
a3,
a4,
a5,
a6,... обозначают константы, они могут быть положительными и отрицательными, большими, малыми и даже равными нулю. Меняя значения этих констант, математик "изгибает" кривую до тех пор, пока она не примет желаемую форму. Для описания простых кривых достаточно ограничиться малым количеством членов такого полинома. Сколько нужно сделать отдельных измерений, чтобы получить изображенный на рисунке полином четвертой степени, т. е. полином, содержащий член
a4x4? Чтобы записать точную формулу, требуется только пять значений, а именно константы
a0,
a1,
a2,
a3 и
a4. Собственно говоря, можно даже обойтись без первого значения, т. е. положить
a0 = 0, тогда ось симметрии листа будет скользить по оси абсцисс. Мы видим, что с каждым новым членом наш полином описывает форму листа несколько точнее. Таким образом, с помощью полинома мы можем описать формы любых объектов независимо от их размеров, а также сравнивать их между собой.
 С помощью полинома, универсальной математической формулы, можно получить почти любую кривую. Чем сложнее кривая, тем большее число членов должен включать соответствующий полином. Можно попытаться подобрать полином, описывающий, например, форму листа
С помощью полинома, универсальной математической формулы, можно получить почти любую кривую. Чем сложнее кривая, тем большее число членов должен включать соответствующий полином. Можно попытаться подобрать полином, описывающий, например, форму листа
 'Машинная улитка'. ЭВМ рассчитала форму улитки, которая лучше всего соответствует реальной
'Машинная улитка'. ЭВМ рассчитала форму улитки, которая лучше всего соответствует реальной
Если мы хотим получить замкнутую кривую, то есть представить лист целиком, то гораздо удобнее записать его форму в так называемых полярных координатах как функцию длины и угла вектора, поворачивающегося вокруг координатной оси.
На следующем рисунке показано, как можно с помощью ЭВМ обсчитать раковину улитки. Структуры аммонитов
[3], так называемые лопастные, или шовные, линии, можно также выразить математически и соответствующие формулы ввести в память ЭВМ, что позволяет детально анализировать форму структур. Это имеет большое значение в палеонтологии и геологии, поскольку аммониты являются одной из самых важных групп "руководящих" ископаемых в некоторых слоях осадочных пород и по малейшим изменениям формы их лопастных линий можно судить о возрасте геологической породы.
 Лопастные линии в раковинах аммонитов. Эти кривые можно выразить математически и ввести в память ЭВМ
Лопастные линии в раковинах аммонитов. Эти кривые можно выразить математически и ввести в память ЭВМ
Итак, форму живого организма можно не только характеризовать размерам, но и описать математически.
Теперь попытаемся с помощью математических формул представить какой-нибудь биологический процесс, например выразить кривую роста. Уму непостижимо: сначала математическое описание формы, а теперь — биологического процесса! Но это кажется трудным только неспециалисту в силу инертности нашего повседневного мышления, привычки воспринимать лишь то, что мы непосредственно ощущаем органами чувств. Форму, то есть три измерения — длину, ширину и высоту, — мы "видим". А изменение этой формы, иначе говоря, изменение этих трех параметров во времени, мы "переживаем". Мы должны запастись терпением и временем и ждать. Для математика время, выраженное в секундах, часах, днях и т. д., такая же счетная величина, как длина и ширина. Если замысловатую форму растения мы выразили с помощью n параметров, представив ее точкой в n — мерном фазовом пространстве, то нам ничего не стоит добавить к ним (n + 1)-й параметр, время, и рассматривать изменение формы растения, т. е. его рост, как ход кривой в (n + 1)-мерном фазовом пространстве.
Итак, из звезды получается комета — звезда с хвостом, летящая по заранее определенному пути. Мы установили, что нумерическая таксономия позволяет представить биологический вид как облачко точек в фазовом пространстве. Если теперь мы добавим время, скажем, продолжительность жизни отдельной особи, то получим уже не облачко точек, а более или менее плотный пучок кривых, представляющих собой кривые роста. Несколько позже мы остановимся на них подробнее, а сейчас хотелось бы высказать еще одно соображение.
Ничто не в силах остановить математика. Не остановился он даже перед авторитетом Декарта, искривив прямоугольную систему координат, названную в честь величайшего математика и мыслителя XVII века "декартовой". Однако в качестве оправдания своих действий математик может сослаться на четвертое правило из "Рассуждений о методе" самого Декарта, которое гласит: "Для познания действительности необходим метод". И в данном случае метод состоит в том, что прямоугольную сетку линий вместе-с нанесенными на нее фигурами непрерывно изгибают в соответствии с формулами. На языке математики этот метод называется преобразованием системы координат.
Более пятидесяти лет назад д'Арси Томпсон написал книгу "Рост и форма" (
On Growth and Form, Cambridge, 1917). Это была одна из первых работ по математической биологии: в ней высказывалась мысль, что преобразование координат удобно применять для описания изменений биологических форм. На рисунке показан пример, приведенный в книге Томпсона, который отражает не рост отдельного организма, а видоизменение, превращение формы в процессе исторического развития. Томпсон пришел к следующему заключению: если рассматривать только внешнюю форму какого-либо организма и задать ее параметры в декартовой системе координат, то форму другого близкородственного организма можно считать результатом непрерывного изменения координат. Следовательно, процесс развития вида в целом можно описать математически соответствующим преобразованием системы координат.
Что это нам дает? Мы нашли способ отобразить природный процесс в формулах и числах. Эти числа вместе с программой мы можем ввести в ЭВМ, и машина с невероятной скоростью и "терпением" проведет самые сложные расчеты. ЭВМ сможет легко рассчитать все возможные промежуточные формы, независимо от того, существовали они или нет на ранних этапах истории Земли. Машина может также экстраполировать, т. е. "мысленно" продолжить путь эволюции. Нам же останется только решить, будет ли "рассчитанный" организм жизнеспособным в реальной действительности или нет. Однако все это справедливо лишь при условии, что с течением времени в соответствии с определенными правилами изменяются только формы, а сами правила, отражающие законы природы, остаются неизменными. Таким образом, буквально на глазах возникает новая проблема. К сожалению, мы не имеем возможности обсуждать ее здесь, хотя в последнее время она получила некоторое развитие.
 На примере формы тела у рыб близких родов [Diodon (А) и Qrthagoriscus (Б)] д' Арси Томпсон показал, что изменение формы можно описать с помощью соответствующего преобразования системы координат
На примере формы тела у рыб близких родов [Diodon (А) и Qrthagoriscus (Б)] д' Арси Томпсон показал, что изменение формы можно описать с помощью соответствующего преобразования системы координат
Мы сделали первый шаг — заключили форму живого организма в систему координат, и не только форму, но и ее изменения. Как мы видели, это не просто, но тем не менее с помощью современной вычислительной техники можно добиться хороших результатов. Итак, в ЭВМ ввели параметры живого организма; она ждет приказа! Что с ними делать? Кое-что мы уже наметили. Машина должна, например, выяснить, насколько интересующий нас организм родствен какому-либо другому. Предположим, что получено число, которое соответствует расстоянию между двумя точками в фазовом пространстве и тем самым позволяет рассчитать степень родства двух организмов. ЭВМ должна установить, к какому виду, к какой расе и с какой вероятностью относится организм, форма которого характеризуется данными параметрами. Она должна высчитать, какую форму должен был бы иметь еще не открытый палеонтологами организм, который по своему геологическому возрасту, с одной стороны, старше, а с другой — моложе уже известных организмов. При этом может выясниться, что форма В произошла не от формы А, а возникла параллельно. Не так уж плохо! Однако в начале главы мы ставили перед собой более смелую задачу — найти ответ на вопрос: почему организм имеет ту или иную форму?
Вернемся к нашей исходной точке зрения, а именно к положению, что организм представляет собой систему, достигшую оптимальности в процессе борьбы за существование, и выясним, насколько применима здесь теория оптимальных процессов. В чем состоит суть этого метода? Это старая и в то же время новая отрасль математики. Старая, потому что данные методы возникли не сегодня, и новая — потому что внедрение и использование их на практике стало возможным только в век совершенных счетных машин. Началом теории оптимальных процессов можно считать формулу, которую свыше двухсот лет назад вывел великий математик Леонард Эйлер. Но предпосылки к созданию этой теории были заложены много раньше.
Излюбленной задачей в курсе дифференциального исчисления в вузах является следующая: "У хозяина есть материал для забора общей длиной l, которым он должен огородить прямоугольный сад со сторонами х и у. Рассчитать, при каком соотношении сторон площадь сада будет наибольшей". Решение этой задачи несложно.
Площадь S прямоугольника рассчитывается по формуле S = xy, где x и y — его стороны. Общую длину забора, т. е. периметр сада, обозначим l, тогда l = 2x + 2y, или х + у — l/2; у = l/2 — х. Заменив в формуле площади y этим выражением, получаем

Эта формула позволяет рассчитать площадь S в зависимости от длины стороны x. Такую зависимость можно представить графически; соответствующая кривая показана на рисунке. Если сторона x очень мала, сторона y, согласно вышеприведенной формуле, должна приблизительно равняться. l/2. Сад превращается в узкое полотно с маленькой площадью. То же получается, когда x велико; x не может быть больше l/2, ибо в этом крайнем случае не хватило бы материала на другие стороны забора и сад состоял бы из двух параллельных заборов, не огораживающих никакой площади. Как подсказывает логика, кривая достигает максимума посередине, а именно в точке, где x принимает значение l/4. Легко подсчитать, что у также должен равняться l/4, следовательно, самую большую площадь имеет квадратный сад. Каждый студент знает, что положение максимума рассчитывается при помощи так называемой первой производной, в данном случае площади S по x. Эта математическая операция позволяет получить новое соотношение, характеризующее наклон функции S в каждой точке x : dS/dx = l/2 — 2x.
 Парабола показывает, как изменяется площадь сада с заданной длиной забора l при изменении длины одной из сторон (например, x). Наклон этой параболы равен ее первой производной dS/dx (прямая линия). Точке пересечения прямой с осью x соответствует максимальное значение площади. Длина стороны x наибольшего по площади (квадратного) сада равна l/4
Парабола показывает, как изменяется площадь сада с заданной длиной забора l при изменении длины одной из сторон (например, x). Наклон этой параболы равен ее первой производной dS/dx (прямая линия). Точке пересечения прямой с осью x соответствует максимальное значение площади. Длина стороны x наибольшего по площади (квадратного) сада равна l/4
Максимум значения S находится в той точке, где наклон кривой равен нулю, т. е. dS/dx = 0. Подставив это значение в предыдущую формулу, получаем l/2 = 2x, или x = l/4. Не правда ли, легкая задача? А вообще говоря, это и есть задача по оптимизации. При заданной длине забора оптимизируется площадь сада.
В начале этой главы мы уже сформулировали типичную задачу по оптимизации. Каков самый короткий путь между двумя точками? Можно ли доказать математически, что это должна быть прямая или, может быть, существует какая-нибудь кривая, которая до сих пор ускользала от нашего внимания?
Здесь школьник уже встанет в тупик: ведь речь пойдет о максимальном или минимальном, т. е. в общем случае об оптимальном значении не какой-то величины, а целой функции. Как решать задачу на максимум-минимум для функции?
Выше мы говорили, что любую функцию, или кривую, можно представить в виде полинома с более или менее большим числом членов и затем охарактеризовать ее с помощью нескольких чисел, а именно констант a
0, a
1, a
2, … a
n. В n — мерном фазовом пространстве такая кривая сводится к точке. Сдвиг точки означает изменение формы кривой, т. е. изменение вида функции.
Рассмотрим следующие уравнения:
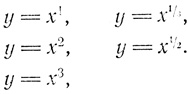
Здесь из полинома выбран только один член. Такое уравнение называют степенной функцией. В общем виде она записывается как у = x
n, т. е. у равен x в n-й степени. С изменением значения n изменяется форма всей кривой.
Предположим, степенная функция описывает форму какого-нибудь биологического объекта и величина n связана с определенным свойством объекта подобно тому, как в предыдущей главе величина тела была связана с теплоотдачей. Этот случай напоминает задачу о длине забора для сада. Выяснив сначала математическую зависимость между величиной n и оптимизируемым свойством объекта, методами диференциального исчисления определяют максимум той математической функции, находят оптимальное значение n и, исходя из него, строят оптимальную кривую.
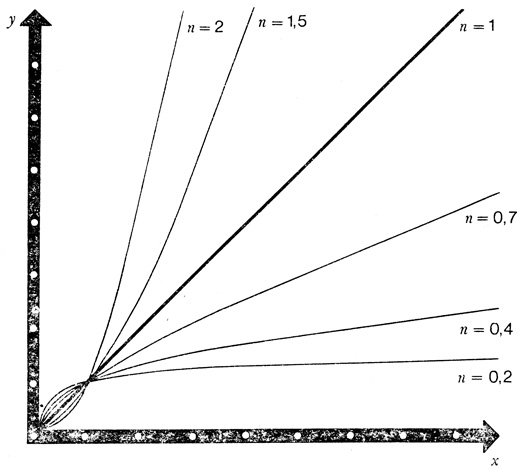 Семейство кривых у = xn. При изменении числа n изменяется форма кривой. Так эволюцию биологических форм в принципе можно представить, изменяя n
Семейство кривых у = xn. При изменении числа n изменяется форма кривой. Так эволюцию биологических форм в принципе можно представить, изменяя n
Однако здесь нам следует остановиться и подвести некоторые итоги. Математическая функция, описывающая какой-либо закон природы, отражает зависимость одной величины от другой и изображается формулой с характеристическими константами, т. е. числами, определяющими ее характер, изменение этих чисел влечет за собой изменение функции.
Характеристические константы могут быть связаны между собой с помощью другой математической функции, которая обусловливает первую функцию; это "суперфункция", или, как ее называют математики, функционал.
Таким образом, поиск оптимального решения сводится к отысканию функционала и определению его максимума (или минимума). Однако насколько просто описать этот процесс словами, настолько сложно выразить его математически, тем более что, как правило, для этого требуется проанализировать связь между многими изменяющимися величинами.
Рассмотрим конкретный пример. Красные кровяные клетки (эритроциты) человека имеют своеобразный вид. Они похожи на резиновые мячики, вдавленные с двух сторон. Для них такая форма оптимальна, потому что она обеспечивает быструю диффузию кислорода к гемоглобину — пигменту крови, содержащемуся в этих клетках. Но более интересна другая проблема, связанная с формой эритроцитов. В гипотонических растворах, т. е. растворах, содержащих меньше солей, чем кровь, кровяные клетки можно "надуть" — в них проникает вода, и они становятся округлыми. Если их осторожно перенести назад в изотонический раствор, восстанавливается прежняя двояковогнутая форма. Чем же она обусловлена? Почему на поверхности эритроцита не образуются другие вмятины? Вопрос интересен еще и потому, что при некоторых болезнях красные кровяные клетки действительно приобретают аномальную форму. Очевидно, нормальная форма красных кровяных клеток человека является следствием оптимального сочетания многих факторов.
Мы начнем анализ этой проблемы с того, что с помощью соответствующей математической формулы представим форму эритроцита в системе координат. Для этого, конечно, можно было бы найти подходящий полином, однако существует более изящный метод. Математикам известна некая кривая, являющаяся геометрическим местом точек, для которых произведение расстояний до двух заданных точек F
1 и F
2 есть величина постоянная. По имени получившего ее ученого она названа кривой Кассини. Итак, для каждой точки на кривой Кассини, изображенной на рисунке, должно быть справедливо уравнение p ⋅ q = а, где р и q — расстояния от этой точки на кривой до двух заданных точек F
1 и F
2. Это уравнение, однако, описывает форму не всей клетки, а только ее поперечного сечения. Форма целой клетки получается вращением ее поперечного сечения вокруг центральной оси.
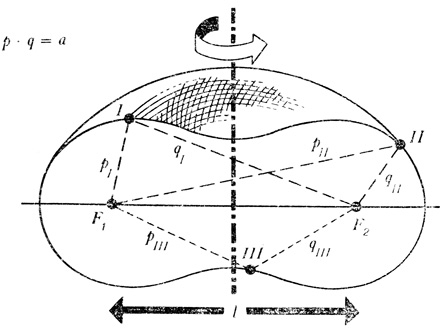 Кривая Кассини — геометрическое место точек, для которых произведение расстояний до двух заданных точек F1 и F2 есть постоянная величина. С помощью этой кривой можно описать форму эритроцитов
Кривая Кассини — геометрическое место точек, для которых произведение расстояний до двух заданных точек F1 и F2 есть постоянная величина. С помощью этой кривой можно описать форму эритроцитов
Таким образом, форму кровяных клеток человека можно охарактеризовать двумя величинами, а именно константой a и расстоянием между точками F
1 и F
2, обозначенным буквой l. Как только отношение а к l изменится, изменится и форма кривой; следовательно, разбухание клеток можно моделировать изменением этого отношения.
Теперь, основываясь на соответствующих физических законах, можно рассчитать энергию, необходимую для деформации эластичной клеточной мембраны при заданных объеме и поверхности клетки. Расчет весьма сложен, но вычислительная машина способна выполнить его с достаточной точностью. В результате мы получим связь между энергией и фактором формы, равным отношению a/l. На рисунке показано, что такую связь, такой функционал можно изобразить в виде кривой, которая имеет один минимум. В точке минимума на деформацию эластичной клеточной мембраны затрачивается наименьшее количество энергии. Как мы видим, в этой точке фактор формы принимает именно такое значение, при котором кривая Кассини соответствует форме эритроцита. Итак, форма наших эритроцитов оптимальна; теперь мы знаем, какие процессы оптимизируются, и в состоянии правильно понять нарушения, связанные с изменением формы эритроцитов у больных. Это может быть одним из путей, ведущих к устранению причин болезни.
Однако подобный анализ объясняет проблему лишь частично. Мы говорили об энергии деформации, обусловленной молекулярным строением клеточной мембраны — пленки толщиной около 1/100 000 мм, окружающей клетку. Для полноты картины следовало бы взглянуть на данную проблему и с позиций молекулярной биофизики, но это увело бы нас далеко от основной темы. Вопросами молекулярной биофизики мы займемся в конце книги. А сейчас вернемся к теме о форме живых организмов в системе координат. Мы видели, что эту форму можно описать математически, т. е. представить в виде чисел и кривых. Более того, с помощью математических выкладок и с учетом физических закономерностей мы можем рассчитать ее изменение, развитие, приспособленность, или, говоря математическим языком, оптимальность. Пример с эритроцитами человека сравнительно прост. Он прост настолько, что хорошая ЭВМ в состоянии его исчерпывающе обсчитать. В большинстве других случаев формы организмов много сложнее и подобные расчеты чрезвычайно затруднительны. Тем не менее такие случаи стоит рассмотреть хотя бы потому, что они позволяют выявить некоторые общие интересные закономерности.
 Эритроцит имеет такую форму, которая при заданных поверхности и объеме клетки соответствует минимальной энергии деформации оболочки. Расчет зависимости между формой и энергией в данном случае достаточно сложен, однако принципиально он не отличается от расчета оптимальной формы сада
Эритроцит имеет такую форму, которая при заданных поверхности и объеме клетки соответствует минимальной энергии деформации оболочки. Расчет зависимости между формой и энергией в данном случае достаточно сложен, однако принципиально он не отличается от расчета оптимальной формы сада
Внешний вид растения, например, можно описать шестью уравнениями, которые отражают как физические условия стабильности, так и законы синтеза и транспорта веществ. С уравнениями такого рода мы уже встречались в первой главе. Если стебель будет слишком высоким, он сломается. То же можно сказать о ветвях, черенках листьев и т. п. С другой стороны, чём длиннее ветви, тем больше поверхность листвы, освещаемая солнцем. Листья обеспечиваются водой, идущей от корней; прежде чем испариться вода должна достигнуть самых удаленных веточек. Каждое из этих требований можно выразить в виде уравнений. Оптимальное решение этих уравнений определяет вид растения. Каков он? Существует одно оптимальное решение или их может быть несколько?
Оптимизация возможна по многим факторам. Вспомним хотя бы пример с постройкой моста. Мост должен быть наиболее дешевым, надежным и легким. Растение тоже стремится к оптимуму по засухоустойчивости, высоте, сопротивляемости ветру, что позволяет ему одержать верх над другими растениями в борьбе за свет. При определенных условиях один из этих факторов может приобрести решающее значение, и тогда оптимизация пойдет по нему; так, кактус обладает оптимальной защитой от засухи, а деревья оптимально используют поверхность листьев. Поэтому и существует не одна форма растений, которая является "особенно" оптимальной, а огромное разнообразие окружающего нас растительного мира.
 Расчет кривой развития биологического вида путем решения дифференциального уравнения дает бесконечное множество кривых, и только выбор начала отсчета а0 отводит одной из кривых роль 'частного решения'. Если в некоторых 'критических' точках кривые расходятся, это означает, что родословная разветвляется. По осям отложены произвольно выбранные параметры организмов a и b
Расчет кривой развития биологического вида путем решения дифференциального уравнения дает бесконечное множество кривых, и только выбор начала отсчета а0 отводит одной из кривых роль 'частного решения'. Если в некоторых 'критических' точках кривые расходятся, это означает, что родословная разветвляется. По осям отложены произвольно выбранные параметры организмов a и b
Остановимся на растениях и попытаемся разобраться в некоторых особенностях их эволюции с точки зрения теории оптимальных процессов. Сущность оптимизации состоит в приспособлении вида к окружающей среде. Как мы уже знаем, развитие формы растений можно проанализировать путем решения задачи по оптимизации, представляя процесс развития в виде математического уравнения. Такого рода уравнения называются дифференциальными. В следующей главе мы рассмотрим математическое описание биологических процессов подробнее, а сейчас отметим лишь, что решением дифференциального уравнения является целое семейство кривых, а не одно или несколько числовых значений.
Итак, мы получим множество кривых развития, различающихся между собой по форме и направлению. И только решив, какую точку на плоскости считать за исходную, мы сможем отбросить все кривые, не проходящие через эту точку, и оставить одну, которая и является действительным, или, как говорят, частным", решением нашей задачи. Следовательно, эволюция нашего растения должна идти вдоль этой кривой в направлении оптимума.
Не исключено, что одна из отброшенных нами кривых в какой-то точке сближается с действительной кривой, а потом они снова расходятся. В такой точке процесс эволюции не стабилен. Путем мелких, случайных изменений, иначе говоря, мутаций,
индивидуум как бы совершает "прыжок в сторону". Он переходит на "боковую колею" и движется уже по другому пути. Процесс эволюции как бы "раздваивается".
Конечно, этот процесс в целом следует рассматривать не на плоскости (мы это сделали исключительно из соображений наглядности), а в n-мерном фазовом пространстве. К сожалению, хотя такие процессы легко описать на словах и выразить математически в общем виде, рассчитать их точно в многомерном пространстве современная вычислительная техника пока не позволяет. Тем не менее подходы к расчету самого сложного явления живой природы — эволюции — в принципе ясны.
Все течет, все изменяется
Заботы натуралиста-коллекционера, или как законсервировать динамическую структуру. Гераклитово "Все течет..." и динамическое равновесие живой природы. Динамические структуры на всех уровнях биологической организации. Можно ли рассчитать динамические структуры? ЭВМ управляет процессами технической микробиологии, рассчитывает дозировку лекарств, следит за работой внутренних органов и биологическим равновесием в жидкостях, содержащихся в нашем теле.
В начале книги мы сравнивали былинку и телевизионную башню, сравнивали формы биологического и технического объектов. Затем мы заинтересовались внешним видом живых организмов и установили, что форма растений и животных тесно связана с их функциями и возможностями. Мы обнаружили, что с помощью чисел и математических формул можно описать внешний вид биологического объекта и его изменение, можно судить о сходстве и различии форм. Как выяснилось уже в первой главе, глубокое понимание формы живых организмов немыслимо без знания различных жизненно важных процессов. Теперь пойдем дальше и постараемся объяснить особенности формы организма, исходя из его функции.
Конечно, зависимость между формой и образом действия, иными словами, между структурой и функцией, характерна и для неживой природы. Часы будут казаться нам бессмысленным предметом из латуни и стали до тех пор, пока мы не поймем, как сцепляются шестеренки, как заводная пружина приводит в действие весь механизм и для чего, собственно говоря, движутся стрелки на циферблате. Если мы определим эту связь, то сможем понять, почему шестеренок именно столько, а не больше или меньше, почему эта шестеренка именно такого размера, а та — другого и т. д. Аналогичным образом в первой главе мы пытались понять связь между структурой и функцией в биологических системах: почему кости имеют определенную толщину, почему мышь не может быть меньше и прочее. Это одна сторона структурно-функциональной связи, принцип которой в равной мере справедлив как для технических, так и для биологических объектов. Если глубже вникнуть в проблему формы живых организмов, то откроется другая сторона, которая имеет решающее значение.
Предположим, мы приобрели исключительно дорогие наручные часы. Это украшение, образец элегантности и точности, символ респектабельности, вещь далеко не повседневная. Мы храним часы в чистом сухом месте с возможно меньшими колебаниями температуры. Мы надеваем их лишь в особых случаях, а затем снова упаковываем. При таком обращении часы сохранят свою работоспособность исключительно долго. Зубчатые колесики не сотрутся, а рубины будут казаться новыми даже нашим далеким потомкам.
Теперь обратимся к биологии. Допустим, коллекционер, поймав редкого жука, бабочку или найдя ценное растение, хотел бы присоединить находку к своей коллекции или передать музею. При этом желательно, чтобы образец как можно больше походил на живой, а по возможности даже был таковым. К сожалению, это исключено. Даже при хорошем уходе животное живет лишь ограниченное время, и нам не удастся сколько-нибудь значительно увеличить время жизни растения, если мы срежем его и поставим в воду. Основная причина наших неудач заключается в том, что структура организма связана с его функцией. Вне функции она мертва. Когда "часовой механизм" живого организма не работает, его "шестеренки" ломаются.
Следует отметить, что современные методы позволяют замораживать и длительное время сохранять жизнеспособные клетки и части органов. Сперма может долго находиться в жидком азоте при температуре -196° С, а после оттаивания снова оказывается способной к оплодотворению. Поговаривают уже о "банке органов", где в замороженном виде будут храниться "запасные части" человеческого тела. Правда, пока еще ни разу не удавалось восстановить жизнедеятельность какого-либо высшего организма после замораживания, но с развитием науки и эта проблема, по-видимому, будет решена. И не исключено, что когда-нибудь мы увидим в жидком гелии Спящую Красавицу.
 В технике статические структуры (А) хорошо сохраняются, если их герметически изолировать от внешних воздействий, таких, как влажность воздуха, колебания температуры, свет и т. д. Динамические биологические структуры (Б) связаны с окружающей средой обменом веществ. Изоляция приводит их к гибели
В технике статические структуры (А) хорошо сохраняются, если их герметически изолировать от внешних воздействий, таких, как влажность воздуха, колебания температуры, свет и т. д. Динамические биологические структуры (Б) связаны с окружающей средой обменом веществ. Изоляция приводит их к гибели
Этот факт ни в коей мере не противоречит нашему постулату о связи между структурой и функцией. Позже мы еще вернемся к разговору о теплоте и температуре. При очень низкой температуре все процессы жизнедеятельности приостанавливаются, в том числе и процессы разложения биологических структур. В отличие от этого механизм часов сохраняется при температуре, которая может быть намного выше или ниже температуры, необходимой для их нормальной заботы.
Принцип сохранения структуры через ее функцию заложен в самом устройстве биологической системы. Почти во всех видах клеток действует механизм, который обеспечивает непрерывную замену старых разрушающихся молекул новыми. И лишь немногие клетки, как, например, уже упоминавшиеся красные кровяные тельца, способны только "изнашиваться". Итак, подведем итоги. Биологические структуры отличаются от технических тем, что при функционировании системы они не изнашиваются, а, наоборот, непрерывно восстанавливаются и поддерживаются, другими словами, функционирование системы является для них условием существования. Как говорил Томас Манн, жизнь колеблется между "ограничивающими ее процессами распада и обновления". Об изменчивости жизни, живых структур люди задумывались еще давно. Древнегреческий философ Гераклит (500 г. до н. э) написал свое знаменитое rcavta pel: "Все течет, все движется и ничто не остается неизменным". Два века спустя эту же мысль высказывал Аристотель.
Мы, как современные естествоиспытатели, будем пользоваться современным языком. Так, в случае шестеренки, неживого, технического изделия, мы говорим о статической структуре, тогда как живую, биологическую систему, например клетку, мы называем динамической структурой. Структура — это нечто видимое и относительно постоянное. Для статической структуры такие свойства очевидны. Шестеренка изготавливается и в процессе работы медленно изнашивается. Ее форма, масса "стационарны", т. е. постоянны, независимы от времени (естественно, в определенных границах).. Что же происходит с динамической структурой? Поскольку эта структура непрерывно разрушается и вновь восстанавливается, ее постоянство заключается только в том, что процессы распада и обновления точно компенсируют друг друга и любое влияние на эти процессы приводит к изменению структуры.
Вспомним надувные резиновые фигуры, на которых с таким удовольствием летом плавают дети.
Если такая игрушка цела и невредима, то достаточно надуть ее один раз, чтобы образовалась статическая структура, форма которой не изменяется в течение некоторого времени. Совсем иное дело, если в ней где-нибудь появится дырка, пропускающая воздух. Обычно в таком случае мы либо заклеиваем дырку либо выбрасываем испорченную игрушку. Но ее можно было бы использовать в качестве модели динамической структуры. Непрерывно подкачивая в игрушку воздух, мы тем самым компенсируем утечку, и резиновый зверь остается более или менее плотным. Выходящий из дырки воздух все время заменяется новым. Утечка воздуха через дырку не постоянна, а зависит от давления. Если надувать игрушку более интенсивно, давление внутри нее повысится и утечка воздуха усилится. Для каждой скорости надувания устанавливается в соответствии с внутренним давлением своя скорость утечки.
Итак, мы видим, что в отличие от статических, динамические структуры регулируются процессами, обусловливающими их разрушение и восстановление. Это очень важная мысль, она объясняет, почему биологические объекты приспосабливаются к изменяющимся условиям много лучше, чем любые технические системы.
Состояние, при котором распад уравновешивается синтезом, называют динамическим равновесием. Этот термин полвека назад предложил Людвиг фон Берталанфи. С тех пор теория динамического равновесия приобрела в биологии большое значение.
Весьма наглядную модель динамического равновесия представляет сосуд со стоком. Если в такой сосуд наливать сверху воду, она будет вытекать снизу. Вначале приток больше, чем сток, и уровень воды постепенно повышается. Но с повышением уровня растет давление, а следовательно, увеличивается скорость вытекания воды. В какой-то момент приток и сток уравновесят друг друга и в сосуде установится некий постоянный уровень воды, т. е. будет достигнуто не зависимое от времени состояние динамического равновесия. Оно поддерживается, конечно, до тех пор, пока приток воды остается неизменным. Если скорость притока увеличить, уровень воды вновь начнет подниматься, пока не установится новое равновесное состояние. И наоборот, если приток воды уменьшить, ее уровень понизится. Аналогичным образом система реагирует и на изменение стока.
 Наполненный водой резервуар с притоком и стоком является хорошей моделью для описания стационарных, т. е. не зависимых от времени, состояний системы. А — статическая структура (оба крана закрыты), которая постоянна из-за отсутствия процессов, зависящих от энергии; Б, В, Г — динамические структуры в различных состояниях равновесиях, которые зависят от положения сливного крана
Наполненный водой резервуар с притоком и стоком является хорошей моделью для описания стационарных, т. е. не зависимых от времени, состояний системы. А — статическая структура (оба крана закрыты), которая постоянна из-за отсутствия процессов, зависящих от энергии; Б, В, Г — динамические структуры в различных состояниях равновесиях, которые зависят от положения сливного крана
Однако оставим нашу модель. Она достаточно наглядна, и возникающие в ней изменения можно легко наблюдать. А встречается ли в живой природе состояние динамического равновесия? Оказывается, это основной принцип функционирования живой системы, который реализуется на всех! уровнях биологической организации.
Начнем с молекул. В последние годы стало возможным получать радиоактивные изотопы всех элементов, в том числе и многих биологически важных. Существуют изотопы, которые химически и биохимически ничем не отличаются от "нормальных" атомов, но их присутствие легко обнаружить по регистрируемому приборами излучению. При помощи этих так называемых радиоактивных изотопов можно "метить" молекулы и следить за индивидуальной заменой одной молекулы другой, а также наблюдать и измерять восстановление отдельных частей молекулы. Эксперименты показали, что все молекулы более или менее быстро заменяются новыми и что постоянство биохимического состава есть не что иное, как состояние динамического равновесия, которое поддерживается на протяжении всего времени существования организма.
 Радиоактивные изотопы позволяют изучать системы, находящиеся в состоянии динамического равновесия, и определять их параметры. В данном случае с помощью изотопа Са45 измеряется кальциевый обмен в крови рыбы. Под действием двух насосов Н раствор циркулирует, в двух замкнутых системах — во внешней среде и кровеносной системе рыбы. Счетчик С регистрирует радиоактивность
Радиоактивные изотопы позволяют изучать системы, находящиеся в состоянии динамического равновесия, и определять их параметры. В данном случае с помощью изотопа Са45 измеряется кальциевый обмен в крови рыбы. Под действием двух насосов Н раствор циркулирует, в двух замкнутых системах — во внешней среде и кровеносной системе рыбы. Счетчик С регистрирует радиоактивность
 Динамическое равновесие на различных уровнях биологической организации. А — молекулярный; Б — субклеточный (митохондрии); В — клеточный; Г — органы и организмы; Д — биоценоз
Динамическое равновесие на различных уровнях биологической организации. А — молекулярный; Б — субклеточный (митохондрии); В — клеточный; Г — органы и организмы; Д — биоценоз
Поскольку состав молекул непрерывно обновляется, то построенные из них клеточные компоненты — митохондрии, хлоропласты, ядро, наружная клеточная мембрана и т. д. — представляют собой не статические элементы, которые можно сравнивать с техническими изделиями, а динамические структуры, распадающиеся и синтезируемые при динамическом равновесии. Динамика заметна и в ионном составе содержимого клетки. В клетке поддерживается ионная среда, по своему составу совершенно отличная от окружающей, хотя в большинстве случаев суммарная концентрация ионов в клетке такая же, как и во внешней среде. По отношению к окружающей среде клетка "изотонична"
[4], ибо в противном случае высокие давления, вызванные процессами осмоса, разрывали бы клетку, тем не менее ионный состав ее иной. Например, в сыворотке крови человека ионов натрия содержится примерно в 10 раз больше, чем ионов калия, а внутри эритроцита соотношение концентраций этих ионов совершенно обратное. В результате возникают сильные градиенты концентраций, благодаря которым ионы калия выходят из клетки, а ионы натрия стремятся внутрь нее. Клетка подобна кораблю, давшему течь, и, чтобы предотвратить гибель этого "корабля", постоянно работают ионные "насосы", удаляя натрий и "накачивая" калий.
 Хотя суммарная концентрация ионов в клетке такая же, как и во внешней среде, концентрации отдельных ионов, особенно ионов калия (К) и натрия (Na), существенно отличаются. Такие жизненно важные для клетки различия поддерживаются специальными ионными насосами, локализованными в клеточной стенке. Однако под действием разности концентраций ионы непрерывно стремятся назад и в результате возникает динамическое равновесие, подобное тому, которое устанавливается в резервуаре с водой
Хотя суммарная концентрация ионов в клетке такая же, как и во внешней среде, концентрации отдельных ионов, особенно ионов калия (К) и натрия (Na), существенно отличаются. Такие жизненно важные для клетки различия поддерживаются специальными ионными насосами, локализованными в клеточной стенке. Однако под действием разности концентраций ионы непрерывно стремятся назад и в результате возникает динамическое равновесие, подобное тому, которое устанавливается в резервуаре с водой
Таким образом, концентрация ионов в клетке поддерживается на достаточно постоянном уровне и находится в динамическом равновесии. Аналогичным образом ведут себя и другие вещества, содержащиеся внутри клетки. Динамическое равновесие другого рода присуще различным органическим молекулам, которые разлагаются в клетке в процессе обмена веществ. Их называют метаболитами. Такие соединения, как, например, разного рода сахара, пассивно проникают (диффундируют) или засасываются клеткой и подвергаются здесь разложению. Следовательно, это не приток и сток, которые взаимно компенсируются, а приток и уничтожение. Такие процессы весьма многочисленны.
 В сообщающихся сосудах обмен водой продолжается и после того, как уровни в них установились. В этом легко убедиться, подкрасив воду в одном из сосудов. Тем не менее такая структура является статической, ибо процесс обмена здесь не требует затрат энергии
В сообщающихся сосудах обмен водой продолжается и после того, как уровни в них установились. В этом легко убедиться, подкрасив воду в одном из сосудов. Тем не менее такая структура является статической, ибо процесс обмена здесь не требует затрат энергии
Далее мы еще будем говорить об энергетическом балансе таких процессов. Здесь же мы только отметим, что не все системы, в которых происходит обмен, относятся к системам динамического равновесия. Представим себе два сообщающихся сосуда. Когда уровни воды в них выравниваются, обмен воды все равно продолжается за счет самодиффузии; но, поскольку этот процесс не связан с преобразованием энергии, подобная система является статической, а не динамической.
Хотя живая клетка как строительный кирпичик животного или растения в целом является динамической структурой, находящейся в состоянии динамического равновесия, некоторые из ее частей имеют статическую природу. Во многих случаях содержание воды в клетке пассивно регулируется ее энергетическим уровнем. В этом отношении клетку можно смело сравнивать с сообщающимися сосудами. Правда, энергетический уровень клетки определяется не силой тяжести, а сложным взаимодействием различного вида энергий. Однако об этом несколько позже.
 В клетке ионы калия (К+) и натрия (Na+) активно 'перекачиваются' (шестеренка и жирные стрелки) и пассивно возвращаются назад (тонкие стрелки), в то же время обмен водой и ионами хлора (Сl-) происходит только пассивно, подобно тому как это осуществляется в сообщающихся сосудах
В клетке ионы калия (К+) и натрия (Na+) активно 'перекачиваются' (шестеренка и жирные стрелки) и пассивно возвращаются назад (тонкие стрелки), в то же время обмен водой и ионами хлора (Сl-) происходит только пассивно, подобно тому как это осуществляется в сообщающихся сосудах
Вначале, обсуждая возможности консервации технических и биологических систем, мы говорили о различии между статическими и динамическими структурами. Теперь же необходимо отметить, что многие компоненты клетки, а следовательно, и всего организма являются структурами статическими. Заглянем в биологический музей. Мы найдем там животных и растения, которые хорошо сохранили свою форму, а частично и окраску. Засушенное растение легко сохраняется десятилетиями, так же как засушенный жук, рак, бабочка, морская звезда или рыба. Отсюда следует, что некоторые части живого организма, в основном те, от которых зависит его форма, т. е. элементы скелета, представляют собой статические структуры, хотя и являются продуктами динамических структур. В определенной степени организм обладает возможностью при необходимости, например в случае повреждений, приводить статические структуры в активное состояние, обеспечивая тем самым их изменение и восстановление.
Мы несколько отклонились от темы, коснувшись проблемы динамического равновесия на клеточном уровне. Теперь мы знаем, что организм в целом представляет собой динамическую структуру, хотя отдельные его части могут иметь статическую природу. Но вернемся к нашей основной темe и попытаемся найти другие, более высокие структурные уровни биологической организации.
Следующую ступень в этой организации составляют биологические системы, образованные совокупностью клеток. К ним можно отнести ткани, органы и целые организмы. Клетки нашего тела постоянно отмирают и заменяются новыми. Исключение составляет только нервная система. Очевидно, специализация нервных клеток слишком высока, чтобы их можно было заменить. Установлено, что из имевшихся вначале 10 млрд. нервных клеток 10 тыс. ежедневно безвозвратно теряются. До сих пор остается загадкой, как наш мозг безболезненно переживает такую потерю.
Клетки других систем восстанавливаются. Типичным примером может служить кровь. Продолжительность жизни эритроцитов у человека в среднем равна 100-120 дням, примерно столько же живут и другие клеточные компоненты крови. Кровь с точки зрения ее состава можно рассматривать как динамическую систему. Если врач установит болезненные изменения в составе крови пациента, то причина их может скрываться как в кроветворении, так и в управляемом процессе разрушения кровяных клеток.
Естественно, что у большинства читателей понятие "управляемый" ассоциируется с кибернетикой. И в самом деле, здесь речь идет в какой-то мере о кибернетической проблеме. О кибернетике, в том числе биологической, написано очень много, поэтому обсуждать этот вопрос мы не будем. Позже мы слегка коснемся кибернетической стороны вопроса,
Заметим, что развитие раковой опухоли можно рассматривать как нарушение динамического равновесия. Внезапно возникающее бурное размножение клеток при прежней скорости их разрушения вызывает увеличение количества клеток подобно тому, как повышается уровень воды в описанном выше резервуаре. Об этом мы также поговорим в дальнейшем.
Пример с клетками крови уже предопределяет переход к биологическим системам следующего, более высокого уровня — биоценозу. Биоценоз — это сообщество одинаковых или различных живых организмов. Деревенский пруд с его водными растениями, водорослями, одноклеточными, червями, дафниями, личинками насекомых и рыбами представляет собой биоценоз; то же можно сказать о луге с его растениями, животными; лесе, болоте и т. д. На горьком опыте человек узнал, что такие экосистемы (биоценозы) являются едиными сложными структурами, узнал после того, как жестоко поплатился за вырубку лесов и истребление некоторых видов животных.
Поскольку мы достаточно говорили о сущности динамического равновесия, теперь нам нетрудно обнаружить динамику в миграции животных, их размножении и гибели. Примечателен тот факт, что в природе взаимосвязь животных и растений, хищника и жертвы играет большую роль. На этом уровне взаимодействие между отдельными элементами системы становится особенно заметным. Очевидно, что в системе, все без исключения элементы которой состоят из отдельных живых объектов, каждая структура должна иметь динамическую природу. Статические структуры, которые мы наблюдаем как на клеточном уровне, так и на уровне отдельных организмов, здесь просто немыслимы. Но, конечно, статические структуры оказывают определенное влияние на динамическое равновесие в живой природе. Правда, эти структуры — характеристика почвы, температура, глубина (для водных биоценозов) и т. д. — представляют собой компоненты неживой природы.
У читателя, возможно, сложилось мнение, что эта глава в отличие от предыдущих не имеет ничего общего с физикой и математикой. Действительно, до сих пор мы довольствовались лишь общими рассуждениями о сущности динамического равновесия. Мы узнали, что биологические системы находятся в состоянии динамического равновесия, т. е. одновременно в статическом и динамическом состояниях. Динамическое равновесие обеспечивает, например, регулярное сокращение клеток сердечной мышцы, готовность к возбуждению у нервных клеток, выделение секрета клетками желез. Динамическое равновесие определяет нормальный качественный и количественный состав клеток крови, "цветение" водорослей в пруду, количество микроорганизмов в промышленных установках по непрерывному производству спирта, пенициллина и т. д.
Из этих примеров видно, как важно знать и заранее рассчитывать условия возникновения динамического равновесия и критерии его устойчивости. Анализ динамического равновесия с помощью "маркированных"
[5] компонентов требует хорошего знания математики и физики. Такие задачи решают сейчас с помощью сложных электронных вычислительных машин. Однако мы попытаемся рассмотреть некоторые принципы теории динамических систем, не углубляясь особенно в математику.
Динамическое равновесие, как мы установили, есть такое изменяющееся состояние, при котором уравновешиваются приток и сток, размножение и гибель, синтез и разрушение. Поэтому условие динамического равновесия мы можем записать в виде простого уравнения
J
AB = J
BA.
Величину J в биофизике называют потоком. С помощью этой величины выражают скорости притока и стока, синтеза и разрушения, частоту размножения и гибели. Иногда это может быть направленная величина, или, как говорят математики, "вектор", иногда просто число — "скаляр". Но в любом случае данный параметр связан со временем: приток в единицу времени, размножение в единицу времени, распад в единицу времени и т. д. Буквы АВ и ВА (индексы при символах потоков) характеризуют направление потока: АВ означает "от А к В", а ВА — "от В к А", т. е. потоки J
АВ и J
ВА противоположны друг другу. В большинстве случаев состояние динамического равновесия определяется не одним, а несколькими потоками, и величины J
АВ и J
ВА представляют тогда сумму потоков в соответствующем направлении.
 Схематическое изображение системы, находящейся в состоянии динамического равновесия. JAB — приток; JBA — cток
Схематическое изображение системы, находящейся в состоянии динамического равновесия. JAB — приток; JBA — cток
Итак, наш разговор уже перекинулся в область биофизики. От общих рассуждений мы перешли теперь к конкретным величинам. Как рассчитывать потоки? Поток — это движение, а каждое движение должно быть обусловлено какой-то силой. Вначале мы просто предполагаем ее наличие и интересуемся возникающими при этом потоками, не задумываясь о происхождении самой силы. Последнее относится к области биоэнергетики, или термодинамики, — науки о преобразовании энергии. Но о ней мы поговорим в следующей главе.
Пусть задана некая "обобщенная" сила: механическая, электрическая или "биологическая". Она является причиной движения, и эту причинную связь мы хотели бы выяснить.
 Динамическое равновесие легче всего рассчитать для резервуара с водой
Динамическое равновесие легче всего рассчитать для резервуара с водой
Проще всего это сделать на примере резервуара с водой. Предположим, приток воды в нем постоянен и зависит только от того, насколько открыт кран. Значительно сложнее обстоит дело со стоком. Даже в том случае, когда на протяжении всего опыта положение крана остается без изменений, скорость вытекания воды через сливную трубу изменяется пропорционально гидростатическому давлению в сосуде. При увеличении давления вдвое через отверстие вытекает вдвое больше воды. Если сосуд цилиндрический, давление прямо пропорционально объему воды. Действительно, в этом случае удвоение объема воды приводит к удвоению высоты водяного столба и, следовательно, к удвоению давления. Мы можем записать

Сток, т. е. поток от А к В, пропорционален объему воды в резервуаре. Знак пропорциональности можно заменить знаком равенства, если ввести некую константу потока, или, как ее иначе называют, константу обмена. Она имеет размерность 1/время (например, 1/мин), а произведение ее на объем дает поток, в данном случае — скорость течения (объем/время). Итак, мы имеем

Это линейное уравнение потока. Линейным оно называется потому, что переменная величина VA входит в уравнение в первой степени, а не в квадрате и не в произведении нескольких переменных. Так как объем VA зависит от времени, т. е. является функцией времени, обозначим объем, занимаемый водой при наступлении равновесия, через V
AРАВН. Как мы уже отмечали, при равновесии J
АВ = J
ВА, следовательно,
Этот простой расчет помог нам выявить важный закон поведения линейной системы — закон "стремления к одному и тому же конечному состоянию".
 Система независимо от ее начального состояния стремится в своем развитии к одному и тому же конечному состоянию. В рассматриваемом случае это равновесный объем VАравн, который определяется только скоростью обмена. Множество кривых, вы-ходящих из разных начальных точек, стремится к этой величине
Система независимо от ее начального состояния стремится в своем развитии к одному и тому же конечному состоянию. В рассматриваемом случае это равновесный объем VАравн, который определяется только скоростью обмена. Множество кривых, вы-ходящих из разных начальных точек, стремится к этой величине
Объем воды в резервуаре V
A полностью определяется скоростью притока J
ВА и константой стока k и совершенно не зависит от того, какое количество воды было в резервуаре в начальный момент времени. Мы можем легко убедиться в этом экспериментально. Подставим сосуд с отверстием под равномерно текущую струю воды. Довольно быстро в сосуде установится динамическое равновесие. Отметим чертой уровень воды и, не убирая сосуда из-под струи, быстро нальем в него воду из другого сосуда. Уровень воды резко поднимется, но не останется таким, а постепенно опустится до отмеченного чертой положения. То же произойдет, если мы искусственно понизим уровень воды. Стремление к одному и тому же конечному состоянию означает, что система рано или поздно вновь приходит к нему независимо от своего исходного состояния. Эта ситуация наглядно представлена на рисунке.
Данный закон применим и к биологической системе. Многие из рассмотренных нами биологических систем, находящихся в состоянии динамического равновесия, при кратковременном воздействии извне вновь возвращаются к своему постоянному положению, определяемому параметрами системы. Позже мы увидим, что нелинейные системы не подчиняются действию этого закона.
Вернемся к соотношению потоков. До сих пор мы рассчитывали, только конечное состояние системы V
AРАВН. Из рисунка видно, что величина V
A может по-разному изменяться во времени. Попытаемся обосновать это теоретически.
Закроем кран, из которого вода поступает в резервуар. Уровень воды в сосуде понизится. Скорость понижения уровня определяется отношением изменения объема к интервалу времени. Сначала уровень воды понижается быстро, потом медленнее. Полученное отношение, таким образом, непрерывно изменяется, и поэтому лучше рассматривать "бесконечно малое" изменение объема в "бесконечно малый" интервал времени. "Бесконечно малое" отличие одной величины от другой дает "бесконечно малую" разницу, или "бесконечно малое" приращение, которое математики называют дифференциалом и обозначают символом d. Следовательно, dV
A есть дифференциал объема и обозначает как раз "бесконечно малое" изменение объема. Соответственно dt — это бесконечно малый интервал времени. Составив отношение dV
A/dt (математики говорят не "dV
A на dt, a dV
A no dt"), мы получим известное каждому старшекласснику отношение дифференциалов, или производную.
Если вода из сосуда не испаряется, то ее объем уменьшается лишь за счет стока. Тогда мы можем записать
-(dV
A/dt) = J
AB
Знак "минус" означает, что положительный сток вызывает отрицательное изменение объема, т. е. его уменьшение. С учетом уравнения потока получим
-(dV
A/dt) = k ⋅ V
A
Это очень важное уравнение, которое справедливо для многих процессов, происходящих как в живой, так и в неживой природе. Оно показывает, что скорость изменения величины, определяемая производной dV
A/dt, прямо пропорциональна самой величине — в данном случае V
А. Чем больше V
A, Т. е. чем дальше кривая удалена от оси абсцисс, тем круче она снижается.
Это уравнение справедливо для случая, когда верхний кран в резервуаре закрыт. Если мы откроем его и обеспечим постоянный приток воды в единицу времени J
ВА, то изменение величины V
А будет определяться другим уравнением:
dV
A/dt = — kV
A + J
BA.
Это уже общее уравнение, которое описывает изменение уровня воды в зависимости от времени в сосуде с двумя открытыми кранами.
Когда система достигает динамического равновесия, V
A больше не изменяется, следовательно, dV
A/dt = 0, а величина V
A в уравнении становится равной V
Aравн. Тогда
0 = — kV
Aравн + J
BA.
Как легко заметить, здесь обнаруживается та же связь между объемом V
Aравн и потоком J
ВА, о которой мы говорили выше.
Однако вернемся к общему уравнению динамической системы. Так как переменная V
А входит в него не только непосредственно, но и через отношение дифференциалов dV
A/dt, такое уравнение называется дифференциальным. Дифференциальные уравнения составляют весьма солидный раздел математики, но здесь мы, разумеется, не имеем возможности его рассматривать. Обсудим только некоторые особенности уравнений такого рода. Решить уравнение — значит найти число, которое, будучи подставленным в уравнение, превращает его в тождество. В отличие от обычных алгебраических уравнений дифференциальное уравнение имеет своим решением не число, а функцию. Поэтому часто говорят не о решении такого уравнения, а о его "интегрировании". Интегрирование есть операция, в известной степени обратная дифференцированию. Поэтому в данном случае интегрирование означает такое преобразование уравнения, при котором производная исчезает.
Учащиеся обычно недолюбливают операцию интегрирования. В то время как производные можно получить, зная общие правила дифференцирования, для интегрирования необходимо знать правильный "прием". И хотя во многих конкретных случаях существуют определенные правила нахождения решения, сформулировать ход решения в общем виде нельзя. Иногда интегрирование вообще невозможно. Поэтому не всякое дифференциальное уравнение можно решить аналитически, иногда приходится довольствоваться приближенными машинными расчетами.
Каждый, кто знаком с основами интегрального исчисления, знает, что в решение интеграла обязательно входят постоянные интегрирования. Необходимость в них отпадает только для так называемых определенных интегралов, в которых пределы изменения функции четко очерчены. Постоянные вводятся и при решении дифференциальных уравнений. Сначала интегрирование дает нам общее решение: бесконечное множество кривых определенного вида. И только после того, как мы находим начальные условия, мы можем "выудить" из них "частное решение".
Это хорошо видно из рисунка, показывающего изменение объема воды в резервуаре. Представленные на нем кривые являются различными частными решениями, которые относятся к разным начальным условиям, но в итоге все кривые стремятся к одному и тому же значению V
Aравн.
Чтобы сделать наше довольно сухое и, казалось бы, столь далекое от биологии изложение более понятным, оглянемся на пройденный путь и подведем некоторые итоги.
Сначала мы установили, что динамическую систему, например обмен веществ в клетке, можно сравнить с резервуаром, в который вода поступает через один кран и вытекает через другой. Затем мы попытались сделать кое-какие расчеты. Для этого мы ввели понятия обобщенной силы и потока, установили соотношения между ними. Оказалось, что в динамической системе изменение величины можно выразить отношением дифференциалов, а поведение всей системы описать дифференциальным уравнением. Решение, или интегрирование, такого уравнения дает нам сначала множество кривых одинакового вида, которые показывают, как будет "развиваться" система из любого исходного положения до тех пор, пока не достигнет стационарного, т. е. не зависимого от времени, состояния динамического равновесия. Особенно важно отметить, что все эти кривые в итоге стремятся к одной точке. Такое поведение простых линейных систем мы назвали "стремлением к одному конечному состоянию". Это свойство и определяет устойчивость биологических систем.
Что же дают нам теоретические представления? Прежде всего они позволяют построить модель любой сложной системы. На рисунке показано, как чисто схематически при помощи гидравлической модели можно представить обмен веществ, протекающий в клетке или в какой-нибудь из ее органелл, например в митохондрии. Если для каждой ячейки (т. е. резервуара) просуммировать потоки по всем направлениям и приравнять их соответствующим отношениям дифференциалов, как мы это делали раньше, то мы снова получим дифференциальные уравнения, решения которых позволяют описать поведение всей системы. Здесь нам опять может пригодиться ЭВМ.
Какое практическое применение имеют подобные расчеты? Допустим, в клинику обращается пациент с заболеванием, причиной которого может быть расстройство функций щитовидной железы. В радиологическом отделении ему дают выпить жидкость, содержащую безвредный для организма быстро распадающийся радиоактивный иод. Счетчик радиоактивности регистрирует интенсивность его излучения в различных частях тела; ЭВМ рассчитывает, исходя из этого, константы скоростей поступления иода в кровь и накопления его в щитовидной железе. На основании этих данных врач может поставить точный диагноз.
 Вещество из внешней среды поступает через клеточную мембрану в клетку К, а затем следует дальше в одну из клеточных структур, 'клеточную органеллу', например митохондрию М (а). Этот процесс можно смоделировать системой из двух наполненных водой резервуаров с насосами Н (б), которые показаны схематически (в). Рисунки расположены в последовательности, соответствующей переходу от конкретной биологической системы к ее все более абстрактному изображению
Вещество из внешней среды поступает через клеточную мембрану в клетку К, а затем следует дальше в одну из клеточных структур, 'клеточную органеллу', например митохондрию М (а). Этот процесс можно смоделировать системой из двух наполненных водой резервуаров с насосами Н (б), которые показаны схематически (в). Рисунки расположены в последовательности, соответствующей переходу от конкретной биологической системы к ее все более абстрактному изображению
В медицине довольно часто возникает необходимость в расчетах такого рода. Как применять лекарство, т. е. в каком количестве и через какие интервалы времени следует принимать или вводить данное вещество, чтобы его концентрация в нужном месте была оптимальной? Лекарство попадает в организм и затем разлагается; от частоты его приема зависит уровень равновесной концентрации. А как влияет интенсивность дыхания на снабжение кислородом определенного органа? Как ускорить выведение из организма токсических веществ?
Однако подобные методы позволяют решать не только медицинские задачи. Везде, где биологические процессы имеют народнохозяйственное значение, в сельском, лесном, водном хозяйствах, в пивоваренной промышленности и виноделии, иными словами, там, где микроорганизмы используются в производстве, оптимальные режимы все чаще и чаще рассчитывают именно таким способом.
 Пример использования системного анализа. Счетчик С регистрирует излучение содержащегося в щитовидной железе радиоактивного иода, который ввели пациенту в целях диагностики. Импульсы регистрируются счетным устройством и записываются в виде кривой изменения радиоактивности во времени. ЭВМ действует по заданной программе и выдает врачу параметры обмена k12, k21..., на основании которых можно поставить точный диагноз
Пример использования системного анализа. Счетчик С регистрирует излучение содержащегося в щитовидной железе радиоактивного иода, который ввели пациенту в целях диагностики. Импульсы регистрируются счетным устройством и записываются в виде кривой изменения радиоактивности во времени. ЭВМ действует по заданной программе и выдает врачу параметры обмена k12, k21..., на основании которых можно поставить точный диагноз
До сих пор мы рассматривали наиболее простые, так называемые линейные зависимости, где скорость изменения какой-либо величины, т. е. поток, связана с самой величиной постоянным коэффициентом. Но как быть, если этот коэффициент изменяется сам? В этом случае возникают нелинейные зависимости, с которыми мы хотели бы познакомить читателя на примере размножения бактерий. Хотя такие нелинейные зависимости весьма характерны для биохимических реакций и процессов транспорта веществ, иначе говоря процессов, происходящих на молекулярном уровне, процесс размножения бактерий в этом отношении наиболее показателен. А поскольку мы уже упоминали, что динамическое равновесие встречается на всех уровнях биологической организации, то будет также полезно и даже необходимо рассмотреть пример из области экологии.
Нальем в пробирку стерильный питательный раствор, который используют микробиологи для выращивания микроорганизмов, и введем туда небольшое количество бактерий или дрожжей. Допустим, в начальный момент времени в растворе находится 10 клеток, живых и способных к делению. Известно, что каждая из них в определенных условиях через определенный промежуток времени делится. Таким образом, через соответствующий интервал времени в растворе окажется 20 клеток. Спустя еще один такой интервал, их будет 40, потом 80, 160, 320, 640, 1280...
Такой процесс размножения можно описать линейным дифференциальным уравнением. Скорость прироста (назовем ее потоком особей J) в этом случае опять пропорциональна общему количеству (клеток), и, как и в прежнем примере, мы можем выразить ее через отношение дифференциалов. Обозначив количество клеток в пробирке буквой n, можно записать
dn / dt = kn
 Так выглядит на графике экспоненциальная фаза роста микроорганизмов: из одной клетки получается 2, из 2-4, из 4-8 г. д. При неограниченном запасе питательных веществ такой ост продолжался бы до бесконечности
Так выглядит на графике экспоненциальная фаза роста микроорганизмов: из одной клетки получается 2, из 2-4, из 4-8 г. д. При неограниченном запасе питательных веществ такой ост продолжался бы до бесконечности
Это уравнение напоминает уже знакомое нам линейное уравнение потока. И действительно, мы опять получили линейную зависимость, но на сей раз без отрицательного знака. Это означает, что в отличие от предыдущего примера, где количество воды в сосуде течением времени уменьшалось, в данном случае число микроорганизмов увеличивается.
Мы не будем заниматься интегрированием этого равнения. Представленная на рисунке формула есть частное решение его для случая, когда количество бактерий в начале опыта, т. е. в момент времени t = 0, было равно n
0. В приведенной формуле появилась новая величина — число е. Это иррациональное число, т. е. число, которое нельзя выразить в виде отношения двух целых чисел; оно равно 2,71828… и является основанием так называемого "натурального" логарифма; им широко пользуются математики. О числе е можно рассказать много интересного, но это уже не имеет никакого отношения к биологии. Вместо е в нашей формуле можно было бы использовать любое другое число, однако в таком случае и величина k стала бы другой.
Как видно, количество клеток n является функцией времени t причем время t в формуле входит в показатель степени. Вообще говоря, прежние наши уравнения также представляют собой экспоненциальные (степенные) функции, в которых k отрицательно, что указывает не на возрастание, а на уменьшение значения функции со временем.
Итак, размножение клеток. Сначала происходящее несколько обескураживает нас. Через каждый промежуток времени концентрация клеток удваивается! К чему же это приведёт в конце концов? Кривая поднимается круче и круче. Это напоминает знаменитую пшенную кашу из сказки о волшебном горшке, которая переполнила дом и вытекла на улицу. Такое может случиться и с маминым тестом и с папиным вином, которое бродит в бутыли. Видимо, наше уравнение в лучшем случае описывает только начало процесса, а затем процесс размножения клеток идет как-то иначе. Что же мы не учли в наших рассуждениях? На самом деле происходит следующее: когда достигается определенная концентрация клеток, питательные вещества среды исчерпываются и бактерии начинают размножаться медленнее, а потом и вовсе прекращают делиться. Как это учесть в уравнении?
Вначале мы приняли, что k — коэффициент пропорциональности, отражающий скорость деления клеток в определенных условиях. Но когда условия ухудшаются, величина k должна уменьшаться. Говоря математическим языком, k теперь уже не постоянная величина, она зависит от числа клеток n. И чтобы идти дальше, нам придется сначала выяснить, какова зависимость k от n. Предположим, что величина k состоит из двух слагаемых. Одно из них пусть будет постоянно; обозначим его k
1 и предположим, к примеру, что с ним связаны некие наследственно закрепленные свойства вида, не зависящие от внешних условий. Второе слагаемое отрицательно, и его абсолютная величина возрастает с увеличением числа клеток; обозначим его k
2n и будем подразумевать, что за ним кроется какой-либо фактор, сдерживающий развитие, например, борьба за существование или, в нашем случае, борьба за пищу.
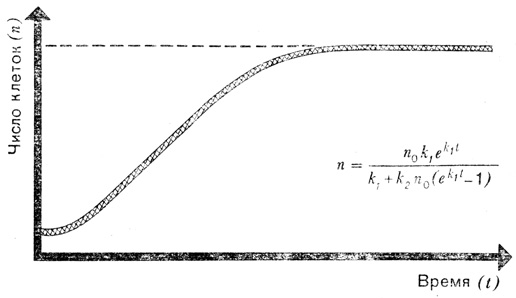 'Логистическая' кривая отражает прекращение размножения клеток после начальной экспоненциальной фазы роста. Уравнение, записанное на рисунке, является наиболее простым из уравнений подобного типа и тем не менее оно намного сложнее приведенного на предыдущем рисунке
'Логистическая' кривая отражает прекращение размножения клеток после начальной экспоненциальной фазы роста. Уравнение, записанное на рисунке, является наиболее простым из уравнений подобного типа и тем не менее оно намного сложнее приведенного на предыдущем рисунке
Итак,

и дифференциальное уравнение принимает вид
Это уже нелинейное дифференциальное уравнение, в которое входит n
2. Чтобы показать, насколько усложняет решение это маленькое дополнение, на рисунке представлено экспоненциальное уравнение, связывающее величины n и t. Но самое важное здесь — новый вид кривой: после быстрого начального подъема кривая стремится к горизонтальной линии, соответствующей равновесному состоянию. Если читатель обладает некоторыми познаниями в алгебре, он может при
желании с помощью этого уравнения провести анализ кривой. Легко показать, что при малых значениях t уравнение превращается в простое уравнение экспоненциального роста. При больших же значениях t нелинейные соотношения между скоростью прироста и общей численностью особей (популяцией) приводят к тому, что кривая загибается.
Такое нелинейное поведение популяции растущих, размножающихся организмов или целого биоценоза, живого сообщества, типично для высших уровней биологической организации. Особенно интересные взаимоотношения возникают там, где один вид животных служит кормом для другого. Зависимость "хищник — жертва" около 50 лет назад рассчитал французский ученый Вольтерра, и с тех пор она неоднократно становилась предметом обсуждения.
Представим себе огороженный участок леса, в котором обитают лисы и зайцы. Предположим, что травы там более чем достаточно, так что зайцы не испытывают недостатка в корме, и нам не придется учитывать эту сторону вопроса. Это упростит решение задачи. Зайцы живут и размножаются. У лис тоже есть пища, и они тоже размножаются. Но когда число лис значительно увеличится, они станут истреблять зайцев в большем количестве, и число тех уменьшится. После этого лисы начнут голодать, некоторые из них погибнут, скорость размножения их понизится. Это приведет к вымиранию лис, и в результате уменьшится число истребляемых ими зайцев. Последних опять станет много, у лис снова появится пища, они будут быстро размножаться, уничтожать зайцев и т. д.
Как показывает этот забавный мысленный эксперимент, динамическое равновесие в данном сообществе так и не наступает: в нем происходят лишь периодические колебания. Это можно доказать математически или, что значительно легче, смоделировать и рассчитать на ЭВМ. Не вдаваясь в детали сложных математических выкладок, рассмотрим результаты таких расчетов. Их можно изобразить графически двумя способами. Если представить количество тех и других животных в зависимости от времени, то мы получим синусоидные кривые; если же на график нанести число зайцев в зависимости от численности лис, то получится так называемый годограф. Если много лис и мало зайцев, точка на годографе скользит направо вниз; зайцы размножились — точка передвинулась налево вверх. Так как синусоиды на верхнем рисунке сдвинуты относительно друг друга, точка на годографе описывает замкнутую кривую. Такой график в честь уже упоминавшегося нами ученого назван циклом Вольтерра. Разумеется, цикл Вольтерра никогда не может касаться оси координат, ибо это значило бы, что вид вымер. В таком случае игра пришла бы к естественному концу. В некоторых случаях кривая представляет собой не замкнутый цикл, а спираль. Это значит, что происходит сдвиг соотношения в пользу одного из видов.
 Численность каждого вида в системе хищник — жертва не устанавливается на каком-либо постоянном уровне, а периодически колеблется. Это можно изобразить либо в виде функции n(t), либо в виде годографа n1(n2) (здесь так называемый "цикл Вольтерра"). Отмеченные цифрами точки цикла соответствуют моментам времени, обозначенным на верхнем рисунке
Численность каждого вида в системе хищник — жертва не устанавливается на каком-либо постоянном уровне, а периодически колеблется. Это можно изобразить либо в виде функции n(t), либо в виде годографа n1(n2) (здесь так называемый "цикл Вольтерра"). Отмеченные цифрами точки цикла соответствуют моментам времени, обозначенным на верхнем рисунке
Мы снова приблизились к той границе, где сегодня кончаются наши возможности разумного использования математики в биологии. Расчеты наземных и водных сообществ живых организмов имеют огромное значение для народного хозяйства, но они чрезвычайно сложны. Сейчас такие расчеты пытаются производить только в микробиологии и гидробиологии, где они связаны с проблемами рыбного хозяйства, очистки вод и охраны окружающей среды.
Теоретическое обоснование колебаний, возникающих в экологических системах, появилось лишь после работ Вольтерра, хотя о существовании таких колебаний было известно уже давно. Еще в средние века люди знали, что эпидемии чумы и холеры, против которых они были бессильны, вспыхивают и затухают периодически, волнами. Такие колебания издавна известны и в экологии. Вероятно, большую роль играют здесь сезонные изменения климата.
Чтобы рассчитать подобные явления, происходящие в окружающем нас мире, необходимо осилить чрезвычайно сложные системы уравнений, правильное решение которых далеко не всегда определяется лишь возможностями вычислительной техники. Чем сложнее уравнения, тем большая неопределенность появляется в результатах расчетов, так как даже небольшие изменения каждого из многочисленных коэффициентов приводят к сильной деформации кривой на графике. Для того чтобы произвести оценки, которые могут оказаться полезными для практики, необходимо решить еще множество теоретических вопросов.
Колебательные процессы, которые мы для наглядности рассмотрели на примере из области экологии, можно встретить также на молекулярном и клеточном уровнях. В особенности это касается ферментативных реакций с характерными для них нелинейными зависимостями. Спектр таких экспериментально обнаруженных и теоретически рассчитанных явлений чрезвычайно широк. Так, регулярные отклонения в клеточных процессах, например периодические изменения концентраций, вызывают колебания, сходные с качанием маятника часов. Во многих случаях такие колебания служат индикатором времени для биологических ритмов. Так называемые "биологические часы", т. е. суточные и сезонные колебания биологической активности, несомненно, объясняются теми же явлениями.
Выше мы уже затрагивали проблему устойчивости. Не все динамические системы подчиняются закону, определенному нами как "стремление к одному и тому же конечному состоянию". Некоторые системы не обладают способностью приходить к одному и тому же конечному состоянию из разных исходных точек. От малейшего изменения в начальном состоянии такой системы зависит, каким путем пойдет ее дальнейшее развитие, подобно тому как одна капля дождя, упавшая всего лишь в нескольких метрах от второй, но по ту сторону водораздела, проследует по Дунаю к Черному морю, между тем как другая попадет вместе с водами Эльбы в Северное море.
Заголовок этой главы гласит: "Все течет, все изменяется". Мы рассмотрели здесь биологические системы в их динамике, динамическое равновесие на разных уровнях биологической организации, переходные состояния, колебания. Мы узнали, что под биологическими структурами следует понимать не только клеточные структуры, которые можно наблюдать с помощью оптического или электронного микроскопов, но и распределение концентраций различных веществ или клеток, распределение организмов в популяциях и многое другое. Такие структуры постоянны только благодаря непрерывному процессу распада и синтеза. В этом смысле они менее постоянны, чем статические структуры в технике. С другой стороны, они не так изнашиваются, потому что непрерывно сами себя "ремонтируют". Таким образом, динамика — основа для функционирования живых систем.
Анализируя линейные и нелинейные уравнения, описывающие различные биологические процессы, мы могли оценить степень сложности этих процессов. Говоря, о "стремлении к одному и тому же конечному состоянию" линейных систем и неоднозначности нелинейных, мы тем самым, хотя бы мимоходом, коснулись вопросов развития этих систем.
Читатель, возможно, заметил, что в наших рассуждениях мы совсем не пользовались понятием "кибернетика". Сейчас появилось много книг по биологической кибернетике, так что нам нет необходимости рассматривать здесь это весьма модное научное направление. Но в принципе рассмотренные нами вопросы можно было бы связать с кибернетикой. Известно, что многие биологические процессы управляются посредством обратной связи, и очень трудно определить границу, отделяющую теорию динамического равновесия от кибернетики. Многое из того, о чем мы говорили, можно без всяких оговорок отнести к кибернетике. Во всяком случае, следует знать, что кибернетика это не игра, оперирующая некими более или менее высокими понятиями, как это, к сожалению, часто бывает, а строгая математическая дисциплина, построенная на основе дифференциальных уравнений.
Беглый взгляд на механизм живого
Часовой механизм живого. Кто его "заводит"? Как он останавливается? В чем заключена движущая сила биологических процессов? Какое отношение к фотосинтезу имеет паровая машина Карно? Термодинамика — всемогущее учение об энергии. От солнечной энергии к химической, электрической, механической. Аденозинфосфаты — химические аккумуляторы. Электрические потенциалы и энергия обмена веществ. Шестеренки сцепляются друг с другом.
Наши размышления заставляют нас все глубже проникать в суть проблемы. Каждый вопрос рождает сотню новых. Начав с формы, мы пришли к движению и установили связь между ними. Сначала это было только наблюдение: нечто двигалось под воздействием какой-то силы, природой которой мы не интересовались. Образно говоря, мы установили, что стрелки часов движутся и это движение подчиняется определенным закономерностям. А теперь сделаем еще шаг вперед: в чем причина движения? Что приводит "шестеренки" живого в движение? Откуда берется движущая сила? Как происходит преобразование энергии?
Мы уже неоднократно обращались к часам как к примеру точной технической системы. Энергетический баланс часов рассчитать нетрудно. Механическая энергия запасается в пружине и затем освобождается опять же в виде механической энергии, приводя в действие часовой механизм. В биологической системе процессы преобразования энергии несравнимо сложнee. Это видно хотя бы из того множества видов "биологического движения", о которых мы говорили в предыдущей главе.
Чтобы подойти к пониманию биологически важных процессов преобразования энергии, нам придется обратиться к истории. В прошлом веке в науке возникло новое направление, которое сначала ставило своей целью лишь одно — улучшение рабочих качеств паровых машин. Это наука об энергии, которую тогда же окрестили термодинамикой — учением о тепловом движении. Достаточно ли высок коэффициент полезного действия паровой машины и как его можно повысить? Можно ли создать вечный двигатель? Почему материал, к примеру тот, из которого изготовляют пушечный ствол, при сверлении нагревается? Таковы были первые вопросы, которые пыталась разрешить эта наука. Со времени своего возникновения термодинамика пережила бурный подъем и продолжает стремительно развиваться в наши дни. Сейчас она служит теоретической основой для расчета всех энергетических процессов, протекающих в природе.
Термодинамику неоднократно пытались использовать для анализа сложных механизмов жизни. Но из-за трудности проблемы все попытки терпели неудачу. Только в последние десятилетия термодинамика достигла такого состояния, которое позволило ей внести важный вклад и в биологию. Сначала идеи термодинамики прочно утвердились в области клеточной и молекулярной биологии, а теперь говорят уже о термодинамике роста, эволюции и даже процесса мышления.
Термодинамика — это выдающееся создание человеческого ума. В ее основе лежат несколько "само собой разумеющихся", эмпирически установленных истин, на которых воздвигнуто логически стройное, надежное и устойчивое здание высшей абстракции. С высоты этого здания, построенного из "математических блоков" на фундаменте точного эксперимента, нам открываются далекие горизонты познания.
Здесь мы не можем останавливаться на частных разделах термодинамики, их развитии и современном состоянии. Однако для понимания общих закономерностей этого и не нужно. Вполне достаточно общего обзора, но нам придется начать издалека.
Сначала о "само собой разумеющемся", о фундаменте термодинамики, о том, что называют "основными законами". Два из них, известные как первое и второе начала термодинамики, особенно важны. Мы попытаемся передать смысл этих законов, не прибегая к математически строгим определениям. Их можно сформулировать следующим образом:
1. Существует нечто, называемое нами "энергией". Энергия обладает следующими свойствами: она не может возникать из ничего, равно как и бесследно исчезать; она существует в различных формах и может переходить из одной формы в другую.
2. Все процессы в окружающей нас природе развиваются в определенном направлении. Самопроизвольные (спонтанные) процессы в замкнутых, или изолированных, системах всегда протекают так, что из всех возможных состояний достигается состояние, характеризующееся максимальной неупорядоченностью.
Это не строгие формулировки, а попытка изложить основные законы термодинамики в наиболее доступной для понимания форме.
Оба начала термодинамики — первое, которое называют также законом сохранения энергии, и второе, позволяющее говорить о возможном направлении самопроизвольных процессов, — послужили исключительно плодотворной почвой для проведения многих расчетов. Первое начало исключает возможность существования вечного двигателя первого рода, т. е. провозглашает бессмысленной давнишнюю мечту людей о машине, которая сама себя непрерывно приводила бы в движение и производила бы при этом полезную работу.
Второе начало также чрезвычайно важно, а сфера его действия столь велика, что это трудно себе представить. Каждый знает, что порядок, если его постоянно не поддерживать, непременно нарушается, — система приходит в неупорядоченное состояние. А что при этом происходит с энергией? Если, например, высокая башня, сложенная из кирпичей, обрушится, то в результате, конечно, освободится энергия. Но такая упорядоченная система, как башня, вряд ли может возникнуть сама по себе из груды набросанных кирпичей даже в том случае, если средняя потенциальная энергия кирпичей в груде и в башне одинакова.
 Вечный двигатель второго рода. Электростанция работает за счет охлаждения речной воды, превращая отобранное тепло в электроэнергию. Такой электростанции не может быть, поскольку самопроизвольное охлаждение тела ниже температуры окружающей среды исключается вторым началом термодинамики. Чтобы использовать тепловую энергию воды, необходимо иметь горячий источник
Вечный двигатель второго рода. Электростанция работает за счет охлаждения речной воды, превращая отобранное тепло в электроэнергию. Такой электростанции не может быть, поскольку самопроизвольное охлаждение тела ниже температуры окружающей среды исключается вторым началом термодинамики. Чтобы использовать тепловую энергию воды, необходимо иметь горячий источник
Взаимосвязь упорядоченности и энергии, о чем мы говорили в предыдущем абзаце, была установлена Людвигом Больцманом лишь через много лет после того, как было открыто второе начало термодинамики. Этот закон играет в биофизике исключительно важную роль, на чем мы остановимся подробнее в дальнейшем. Сейчас же обсудим энергетическую сторону этого закона. Второе начало термодинамики исключает возможность существования вечного двигателя второго рода, т. е. машины, которая отнимает у какого-либо тела тепло и превращает его в полезную энергию, например электрическую.
Покажем это на примере. Температура воды в реке летом около 20° С, в то время как летний воздух несколько теплее. А нельзя ли построить на реке электростанцию, в которой вода охлаждалась бы до 10° С, а полученная тепловая энергия превращалась бы в электрическую? Вблизи такой электростанции приятно было бы отдохнуть от летней жары, а через несколько километров ниже по течению вода в реке снова приобретала бы нормальную температуру. Первое начало термодинамики здесь удовлетворяется. Электроэнергия получается не из ничего, а из тепловой энергии речной воды. Казалось бы, все в полном порядке! Тем не менее такой электростанции нет и быть не может. Второе начало термодинамики запрещает подобное направление протекания самопроизвольного процесса.
Следует отметить, что тепловая энергия занимает особое место среди всех прочих видов энергии. Энергия любого вида — будь то электрическая, химическая, механическая и т. п. — может целиком превращаться в тепловую энергию. Тепловая же энергия превращается в другие виды энергии только тогда, когда существует разница температур. Но даже и в этом случае возможно лишь частичное преобразование энергии. Впрочем, это вещь очевидная, однако и она отражена во втором начале термодинамики. Каждый из нас замечал, что чай в стакане охлаждается, то есть его тепло передается окружающей среде. Однако никому не приходилось наблюдать, чтобы чай в стакане нагрелся за счет окружающего тепла до температуры выше комнатной!
 Температура тела всем известного воробья ТТ = 41 °С. Она выше температуры внешней среды, равной, скажем, ТВ = 10 °С. Такой перепад температур вызывает постоянную потерю тепла, или, говоря языком термодинамики, рассеяние энергии
Температура тела всем известного воробья ТТ = 41 °С. Она выше температуры внешней среды, равной, скажем, ТВ = 10 °С. Такой перепад температур вызывает постоянную потерю тепла, или, говоря языком термодинамики, рассеяние энергии
Можно привести множество подобных примеров. Но естествознание не сводится к одним только рассуждениям; его задача — найти точные математические решения тех или иных вопросов, а для этого необходимы точные определения, величины, зависимости. Выдающимся достижением физики нашего века следует считать то, что ей удалось выразить в формулах основные положения термодинамики и обеспечить тем самым возможность проведения точных расчетов. Но прежде чем перейти к формулам, нужно найти параметры, которыми можно оперировать и которые можно надежно измерять.
Проблема начинается уже с определения понятия энергии. Что это такое? Как ее измерять? Какие явления в окружающем нас мире можно охарактеризовать этим словом? Чтобы ответить на эти вопросы, следует сначала ясно представить себе, какими свойствами должна обладать энергия. Мы говорили, что существуют различные формы энергии и их необходимо измерять, суммировать, сопоставлять друг с другом. Энергия должна быть параметром, определяющим состояние системы независимо от того, как это состояние достигнуто. Энергия должна быть такой же характеристикой состояния системы, как, например, высота расположения тела над уровнем моря, температура, электрический заряд, внутреннее давление и т. д.
Однако мы рискуем слишком уклониться от наших целей и потому постараемся быть краткими.
Итак, поверим на слово, что физикам действительно удалось установить такие энергетические параметры системы, которые можно измерять и с которыми можно проводить различные расчеты.
К сожалению, мы еще не покончили с теорией. Для того чтобы второе начало термодинамики сформулировать количественно, наряду с энергетическими параметрами, характеризующими состояние системы, необходимо еще одно важное понятие. Около 100 лет назад Клаузиус ввел величину, которая могла бы служить качественной мерой тепла. Он назвал ее энтропией. Позже Людвиг Больцман связал ее с понятием вероятности и тем самым с упорядоченностью. Термин "энтропия" часто понимают и используют неправильно. Поэтому мы будем употреблять его как можно реже и только там, где его можно наглядно представить. Просто отметим, что энтропия — это величина, изменение которой определяет направление любого самопроизвольного процесса в замкнутой системе. Эти процессы протекают всегда так, что энтропия системы увеличивается. Данное положение можно рассматривать как еще одну формулировку второго начала термодинамики.
Размерность энтропии — калория/градус (температура измеряется по шкале Кельвина), отсюда ясно, что произведение энтропия X температура = тепло. Это тепло — своего рода дань, которую система отдает окружающей среде при каждом акте преобразования энергии. Любой электромотор при работе становится теплым, бензиновые моторы требуют охлаждения, тормоза нагреваются. Каждый процесс преобразования энергии в системе сопровождается выделением в окружающую среду тепла, которое система теряет безвозвратно.
Но не пора ли, наконец, перейти от физики к биологии? Ведь читатель уже может с полным правом спросить: "Что же все это означает?" Наш собственный повседневный опыт поможет нам сделать этот переход. Кто из нас не чувствовал, как тело выделяет тепло, когда, потея, влезал на гору, колол дрова или копал землю? Именно такие наблюдения заставили ученых пристальнее взглянуть на потерю энергии живым организмом и изучить этот вопрос экспериментально.
 С помощью такого ледяного калориметра Лавуазье и Лаплас в 1780 г. определяли образование тепла у морской свинки. Тепло, выделяемое животным, вызывает таяние льда во внутреннем сосуде. Внешняя ледяная рубашка предохраняет калориметр от нагревания извне
С помощью такого ледяного калориметра Лавуазье и Лаплас в 1780 г. определяли образование тепла у морской свинки. Тепло, выделяемое животным, вызывает таяние льда во внутреннем сосуде. Внешняя ледяная рубашка предохраняет калориметр от нагревания извне
Вообще говоря, попытки провести подобные исследования предпринимались задолго до открытия первого начала термодинамики. Еще в 1777 г. французский ученый Лавуазье попытался исследовать связь между образованием тепла у животного и его дыханием. Спустя три года к этим опытам присоединился его соотечественник Лаплас. Исследователи помещали морских свинок в ледяной калориметр и установили, что за 10 часов животное "производит" тепло, достаточное для того, чтобы растопить 341 г льда. Предположив, что теплота образуется в результате физических процессов сгорания, и определив, сколько двуокиси углерода выделяет морская свинка за то же время, Лавуазье рассчитал количество тепла, выделяющееся при сгорании; оно оказалось достаточно близким к измеренной величине.
 Современный калориметр позволяет точно измерять теплоотдачу различных объектов, подопытных животных и даже людей
Современный калориметр позволяет точно измерять теплоотдачу различных объектов, подопытных животных и даже людей
Этот первый опыт, положивший начало термодинамическим исследованиям в биологии, позже повторили другие естествоиспытатели, которые уточнили результаты Лавуазье и Лапласа. При помощи усовершенствованных приборов они изучили тепловой баланс у различных животных и человека и определили чрезвычайно важные энергетические параметры, характеризующие обмен веществ.
В последнее время такие измерения вновь стали актуальными. Выяснилось, что потеря тепла на единицу массы тела в ходе развития организма не постоянна. Так, при развитии лягушки из икринки сначала наблюдается резкий подъем кривой потери тепла, а затем ее спад до минимального значения. Подобные эксперименты заложили основу термодинамики процессов развития биологических систем. Живой организм представляет собой открытую систему, которая обменивается веществом и энергией с окружающей средой. В термодинамике существует теоретически обоснованный закон, согласно которому открытая система развивается всегда так, что потеря энергии, или скорость выделения тепла, другими словами, прирост энтропии стремится к минимуму. Применим ли этот закон к эмбриональному или даже видовому развитию организмов? Вторая половина кривой, описывающей развитие лягушки, говорит в пользу этого положения. Кривые подобного типа были получены и во многих других исследованиях. Однако не исключено, что наблюдаемые явления в этих случаях лишь внешне похожи на только что описанное, а на самом деле они обусловлены совсем другими причинами.
 Рассеяние энергии, или, говоря иначе, прирост энтропии, в процессе эмбрионального развития организма сначала увеличивается, а затем стремится к минимуму. В настоящее время идут горячие споры о справедливости применения к биологическим объектам закона, согласно которому любая открытая система стремится к состоянию, при котором прирост энтропии минимален
Рассеяние энергии, или, говоря иначе, прирост энтропии, в процессе эмбрионального развития организма сначала увеличивается, а затем стремится к минимуму. В настоящее время идут горячие споры о справедливости применения к биологическим объектам закона, согласно которому любая открытая система стремится к состоянию, при котором прирост энтропии минимален
Здесь действительно проходит сейчас граница применимости принципов термодинамики в биологии. Поэтому будем осторожны в своих выводах и подождем результатов дальнейшего развития этой науки. В настоящее время вырисовывается новое направление термодинамики, так называемая нелинейная термодинамика, которая обещает много интересного. Будущее покажет, оправдает ли она наши надежды. Сложные биологические процессы, такие, как процессы эмбрионального и эволюционного развития, пока лишь в очень малой степени поддаются расчетам. Поэтому мы остановимся сначала на самых элементарных процессах, протекающих в живом организме.
До сих пор мы говорили только об одной стороне энергетического баланса тела — теплоотдаче, или, как мы еще раньше ее назвали, рассеянии энергии. Эта сторона энергетического обмена веществ по праву вызывает наибольший интерес. Ведь если "тепловая дань" платится при каждом процессе преобразования энергии, то по образованию тепла или по соответствующей ему потере энергии можно судить о протекании энергетических процессов, подобно тому как по тиканью часов мы убеждаемся в действии часового механизма. Однако если нас интересует полный энергетический баланс, то нам следует не только учесть выделение тепла организмом, но и выяснить происхождение энергии в живом организме. Тиканье часов свидетельствует лишь о том, что часы идут, но ничего не говорит об их механизме.
Начнем с энергетического баланса у животных. На вопрос об источнике энергии в данном случае ответить легко. Организм "сжигает" потребляемые в виде пищи органические вещества, как примус, керосинка или автомобильный двигатель внутреннего сгорания, и использует освободившуюся энергию. Это сравнение, правда, касается только принципа использования энергии, но не механизма процесса ее "производства". В отличие от названных технических устройств в организме "сгорание" происходит без пламени. Путем сложного чередования ферментов, то есть биологических катализаторов, энергия постепенно отнимается у "сжигаемого" вещества и передается специальному химическому "аккумулятору". В результате образуются богатые энергией молекулы, так называемые "макроэргические соединения", которые диффундируют в ту часть организма, где они необходимы. Здесь присутствуют специальные ферменты, которые способны разряжать эти химические аккумуляторы и использовать освободившуюся энергию в каком-нибудь специфическом процессе.
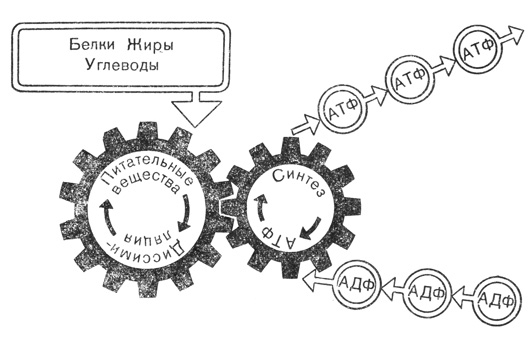 Таким образом очень схематически можно представить процесс образования энергии в теле животного. Богатые энергией питательные вещества разрушаются 'цепью' ферментов. Освобождающаяся при этом химическая энергия идет на зарядку АДФ — АТФ — аккумулятора
Таким образом очень схематически можно представить процесс образования энергии в теле животного. Богатые энергией питательные вещества разрушаются 'цепью' ферментов. Освобождающаяся при этом химическая энергия идет на зарядку АДФ — АТФ — аккумулятора
Существует много веществ, которые могут служить внутренними аккумуляторами энергии. Самые известные среди них — аденозинфосфаты. Наиболее богат энергией аденозинтрифосфат, или сокращенно АТФ, самый бедный — аденозинмонофосфат (АМФ), промежуточное положение занимает аденозиндифосфат (АДФ). Насыщенность этих соединений энергией увеличивается пропорционально количеству связанных фосфатных групп. Одна (АМФ), две (АДФ) или три (АТФ) связанные фосфатные группы определяют высоту энергетического уровня этих соединений. Обычно АТФ — аккумулятор разряжается только до уровня АДФ.
Наступило время сказать несколько слов о природе химической энергии. Этот вопрос не относится к теме нашей книги. Однако пришлось же нам говорить о тепловых электростанциях, хотя они тоже не имеют ничего общего с биологией. Попробуем разъяснить суть этой формы энергии в двух-трех словах. Совершенно очевидно, что химическая энергия связана с молекулами, атомами, ионами. Общая энергия складывается из энергий отдельных молекулы, атома, иона, умноженных на количество таких частиц. Следовательно, химическая энергия зависит от концентрации! Это важный вывод. Но откуда же берется энергия отдельной частицы? Точно ответить на этот вопрос можно лишь с помощью квантовой механики. И опять мы вынуждены подчеркнуть, что изложение принципов квантовой механики выходит за рамки нашей книги. Но следует набросать хотя бы грубую картину явлений, происходящих в мире частиц. Для этого нам придется обратиться к модели атома, созданной Бором.
Согласно этой модели, атом состоит из ядра и вращающихся вокруг него по разным орбитам электронов. Отрицательно заряженные электроны притягиваются к положительно заряженному ядру, и ядро захватило бы их, если бы притяжение не уравновешивалось центробежной силой. Уже такая простая система обладает энергией, подобно привязанному за нить камню, который мы вращаем над головой. Если теперь два атома соединяются в молекулу, то есть химически взаимодействуют, орбиты внешних, так называемых валентных, электронов изменяются. Теперь электроны движутся вместе по сильно измененным орбитам, иногда вокруг обоих ядер сразу. Легко понять, что при этом энергия, запасенная в такой системе, меняется. Молекулы биологических систем состоят из огромного множества атомов. Современная квантовая механика пока не позволяет рассчитать энергию таких молекул. Но мы хорошо умеем измерять выделение или затрату энергии, которые происходят при превращении одного вещества в другое. Часто энергию выражают не в абсолютных значениях, а в относительных величинах, которые определяются относительно какой-либо произвольно выбранной стандартной величины.
Что же разумнее всего выбрать в качестве такой величины при измерении химической энергии? За энергию химического соединения проще всего принять ту энергию, которая необходима для построения этого соединения из отдельных элементов. Однако большинство соединений невозможно получить экспериментально из отдельных элементов: соответствующие реакции не протекают. Но пусть это нас не огорчает. Здесь нам пригодится то положение, которое мы включили в определение понятия энергии, а именно что энергия — величина, характеризующая состояние системы; она не зависит от того, каким путем это состояние достигнуто. Совершенно безразлично, как пройден путь от элемента до соединения, прямо или запутанно, кружной дорогой, в том или ином направлении. Если мы будем тщательно учитывать энергетический баланс каждого этапа, то есть суммировать выделяющуюся энергию и вычитать затраченную, то общий баланс энергии всегда окажется одним и тем же. Если вещество нельзя синтезировать из элементов, то его всегда можно разложить. При этом освободится ровно столько энергии, сколько затратив лось бы при его синтезе. Лучший путь разложения вещества — сжигание. В процессе сжигания вещество разлагается если не до отдельных элементов, то все же до весьма простых соединений. При "сжигании" органических молекул чаще всего выделяются вода, двуокись углерода и окись азота. А эти неорганические соединения уже легко синтезировать из отдельных элементов. Таким образом удается проследить путь от элемента до молекулы. Круг замкнулся.
Учитывая сказанное, можно проделать следующие расчеты. Если результаты калориметрических измерений, полученные в опытах с животными, дополнить данными о потреблении пищи и кислорода и о выделении углекислого газа, то, зная энтальпию (так называют соответствующую измеряемую величину) сгорания пищи и энтальпию образования молекулярного кислорода и углекислого газа, можно вывести энергетический баланс для всего организма, выяснить, соответствует ли ожидаемое реальному, и вообще справедливо ли первое начало термодинамики. Оказывается, справедливо! Конечно, в пределах точности проводимых измерений.
 Для определения теплоты, или энтальпии,сгорания применяют специальный калориметр. Исследуемую пробу помещают в платиновый тигель ПТ, который ставят в закрытый сосуд с кислородной атмосферой. Затем пробу поджигают с помощью электрической искры и она сгорает. Повышение температуры в водяной бане регистрируют с помощью чувствительного термометра Т, Мешалка М обеспечивает перемешивание воды
Для определения теплоты, или энтальпии,сгорания применяют специальный калориметр. Исследуемую пробу помещают в платиновый тигель ПТ, который ставят в закрытый сосуд с кислородной атмосферой. Затем пробу поджигают с помощью электрической искры и она сгорает. Повышение температуры в водяной бане регистрируют с помощью чувствительного термометра Т, Мешалка М обеспечивает перемешивание воды
Итак, мы познакомились с заводной "пружиной" биологического механизма — разложением органических молекул. Теперь пора заняться ведомыми "шестеренками", то есть отдельными процессами преобразования энергии. Прежде всего возникает вопрос: откуда вообще берутся органические молекулы? Как известно, существуют автотрофные ("самопитающиеся") и гетеротрофные организмы. Первые синтезируют для себя питательные вещества сами, вторые используют в качестве пищи другие организмы
[6]. Автотрофные организмы — это зеленые растения, которые мы выращиваем на окнах; как правило, они не нуждаются для своего роста в органических веществах и вполне довольствуются неорганическими солями.
Откуда же в этом случае берется необходимая для обмена веществ энергия? Мы знаем, что растения развиваются только на свету. Свет представляет собой электромагнитные колебания с широким спектром длин волн или частот. Каждый участок видимой области электромагнитного спектра мы воспринимаем как тот или иной цвет. Видимый спектр охватывает область от темно-красного до фиолетового света. Красному свету соответствует наибольшая длина волны (около 0,8 мкм) и наименьшая частота колебаний, фиолетовому — самая короткая длина волны (0,4 мкм) и самая высокая частота колебаний. Существует также инфракрасное, или тепловое, излучение, лежащее в более длинноволновой области за видимым красным светом, и невидимое ультрафиолетовое излучение, расположенное за фиолетовой областью спектра.
Согласно современным физическим представлениям, электромагнитное излучение можно рассматривать не только как волну, но и как поток мельчайших частиц — фотонов. С позиций квантовой теории длина волны (или частота колебаний), от которой зависит окраска света, обусловлена кинетической энергией фотонов. У синего света, обладающего самой короткой длиной волны, фотоны наиболее богаты энергией, у красного света, имеющего наибольшую длину волны, энергия фотонов самая низкая. Свет и темнота, то есть интенсивность света, которую мы измеряем фотоэлектрическим измерителем освещенности, определяются суммарным потоком энергии, равным произведению энергии кванта на поток квантов.
 Видимый свет состоит из качественно различных компонентов, соответствующих разным цветам спектра. Каждый компонент характеризуется своей длиной волны, частотой колебаний или энергией кванта
Видимый свет состоит из качественно различных компонентов, соответствующих разным цветам спектра. Каждый компонент характеризуется своей длиной волны, частотой колебаний или энергией кванта
Какое отношение имеют эти рассуждения к фотосинтезу? Они помогают нам установить связь с термодинамикой, о которой мы говорили в этой главе.
Мы уже знаем, что тепловую энергию можно использовать только в том случае, если существует разность температур. Солнечные батареи, применяющиеся в технике, работают в основном с использованием теплового компонента света, т. е. области, наименее богатой энергией. Солнечные лучи фокусируются в одной точке, в результате выделяется большое количество тепла, которое можно использовать непосредственно или преобразовать в иные виды энергии. Однако в технике существуют и другие преобразователи, солнечной энергии, которые превращают энергию света в электрический ток, минуя теплоту. К ним, например, относятся уже упоминавшиеся фотоэлектрические измерители освещенности.
Растения также используют отнюдь не тепловой компонент солнечного света. Такой принцип преобразования энергии не только рациональнее теплового, он целесообразен еще и потому, что биологические объекты могут нагреваться выше температуры окружающей среды лишь в очень ограниченных пределах. А ведь для эффективного использования тепла необходимы большие разности температур, что, конечно, приведет к разрушению клеток организма. В этом отношении свет с короткой длиной волны, т. е. с высокой энергией квантов, обладает неоспоримыми преимуществами. Такая энергия уже не действует как теплота, не действует просто потому, что температура вещества отражает тепловое движение его молекул и атомов, а кванты с высокой энергией не могут заставить колебаться достаточно быстро эти относительно большие частицы.
Высокочастотный видимый свет способен индуцировать колебания лишь отдельных частей молекул и атомов, например электронов внешней оболочки. Несмотря на это, излучение с большей энергией формально связывают с более высокой температурой. Иногда говорят даже о температуре излучения, подразумевая под этим специфический спектр излучения нагретых тел. Например, в электрических лампочках температура излучения изменяется от температуры красного до температуры белого каления металлической нити.
 Хлорофилл не растворен однородно в цитоплазме растительной клетки, а упорядоченно расположен в гранах — субмикроскопических структурах хлоропластов
Хлорофилл не растворен однородно в цитоплазме растительной клетки, а упорядоченно расположен в гранах — субмикроскопических структурах хлоропластов
Живая природа сделала важное изобретение. Она создала хлорофилл, магнийсодержащий хромопротеид, то есть окрашенный белок, который в большое количестве содержится в растениях и обусловливает зеленый цвет листьев. Он поглощает свет в двух интервалах длин волн — наиболее активно в красной области, длина волны 0,68 мкм, и немного хуже — в синей, длина волны 0,44 мкм. Второй пик поглощения света имеет для фотосинтеза решающее значение. Здесь заключена энергия очень высокого "качества", растение отфильтровывает из всего солнечного спектра кванты с такой энергией, которые легче всего использовать.
Как в данном случае происходит процесс преобразования энергии? Мы уже говорили, что свет с высокой энергией вызывает колебания не столько самих молекул, сколько отдельных их частей. Высокая энергия оказывает воздействие только на электроны. Вспомним наши рассуждения о химической энергии и модели атома Бора с его электронными орбитами: каждой электронной орбите соответствует определенный энергетический уровень. При поглощении фотона энергия электрона увеличивается, в результате он переходит на более удаленную от ядра орбиту.
Обычно электрон быстро возвращается в исходное состояние, отдавая при этом "лишнюю энергию" в виде флуоресценции
[7]. Особенность молекулы хлорофилла и ее белкового окружения в хлоропласте растительной клетки состоит только в том, что свой обратный путь электрон проходит через "машину" обмена веществ, которая забирает и использует энергию, освобождающуюся при переходе электрона в прежнее состояние. Помимо всего прочего, при этом возникает АТФ, аденозинтрифосфат, который уже знаком нам как химический аккумулятор. Синтез глюкозы из углекислого газа воздуха и воды, обычно называемый фотосинтезом, является вторичным процессом, в ходе которого используется химическая энергия образовавшихся молекул АТФ.
На первый взгляд все достаточно просто. Но это лишь кажущаяся простота. На самом деле точный расчет энергетического уровня различных возбужденных состояний атомов и молекул в квантовой биофизике и квантовой механике является исключительно сложной задачей. Трудно даже сказать, можно ли вообще точно рассчитать возбужденные состояния и изменения сложных биологических молекул на современном уровне развития квантовой механики. Поэтому оставим этот вопрос до будущих времен и продолжим наше знакомство с часовым механизмом живого.
До сих пор мы обращали внимание только на две формы энергии — тепловую и химическую — и осветили этот вопрос довольно односторонне. Тепловую энергию мы рассматривали лишь как неизбежное зло, своего рода потерю энергии. Однако теплота — это, конечно, не неизбежное зло, а жизненно важная необходимость. Для функционирования живых систем требуется оптимальная температура, которая у теплокровных животных поддерживается строго постоянной независимо от температуры окружающей среды. Отдача тепла строго регулируется: оно не должно рассеиваться ни слишком сильно, ни слишком слабо. Это мы уже неоднократно подчеркивали.
 Различные способы регуляции рассеяния тепла у животных. Вращением крыльев летающие насекомые разогревают тело до необходимой температуры. Высунутый язык собаки способствует рассеянию излишнего тепла, то есть охлаждению ее тела
Различные способы регуляции рассеяния тепла у животных. Вращением крыльев летающие насекомые разогревают тело до необходимой температуры. Высунутый язык собаки способствует рассеянию излишнего тепла, то есть охлаждению ее тела
О химической энергии мы также рассказали слишком кратко. У читателя могло сложиться впечатление, будто растения синтезируют органические молекулы только для того, чтобы обеспечить пищей животных. Подобный принцип целесообразности, принцип "для того... чтобы" присущ телеологии и, конечно, совершенно неприемлем для естествоиспытателя. И если он где-то и выплыл в нашей книге, то лишь по причине удобства и простоты. Но, даже если исключить эту методологическую ошибку, такие представления были бы неверны. Сложные органические молекулы — это не только метаболиты, но и одновременно строительный материал для живого организма.
 Примерно таким образом в сильно упрощенном виде можно представить энергетический баланс у растений и животных. В прямоугольниках указаны различные формы энергии. Каждая стрелка соответствует цепи сложнейших реакций
Примерно таким образом в сильно упрощенном виде можно представить энергетический баланс у растений и животных. В прямоугольниках указаны различные формы энергии. Каждая стрелка соответствует цепи сложнейших реакций
Таким образом, следует добавить: химическая энергия необходима организму в первую очередь для того, чтобы построить часовой механизм живого. Шестеренки "живых" часов должны сами себя синтезировать и поддерживать в динамическом равновесии. , как мы установили еще в предыдущей главе, чтобы поддерживать состояние динамического равновесия, нужна энергия.
Теперь, сделав все необходимые дополнения и уточнения, касающиеся химической и тепловой энергий, мы рассмотрим другие формы энергии, имеющие значение для функционирования живого организма. И растения, и животные совершают механические движения, которые также требуют затрат энергии. Механизмы таких движений у растений и животных совершенно, различны. О специфике многих движений у растений мы еще будем говорить. В настоящее время наиболее изучен процесс образования механической энергии в скелетных мышцах. Остановимся вкратце на современном состоянии вопроса.
 Физические методы позволяют измерять самые разные формы энергии, которыми обладают растения, животные и микроорганизмы. А — за счет работы осмотических сил растение создает гидростатическое давление Δp, которое гонит воду вверх по стволу; Б — все живые организмы производят механическую работу. Белка увеличивает свою потенциальную энергию Eпот, поднимая свое тело массой m вверх по стволу на высоту h и преодолевая при этом ускорение силы тяжести g; В — электрические рыбы (на рисунке изображен электрический угорь) создают напряжение сотен вольт; Г — в светлячке "спрятаны микроорганизмы, испускающие свет, энергия квантов которого равна произведению постоянной Планка h на частоту ν
Физические методы позволяют измерять самые разные формы энергии, которыми обладают растения, животные и микроорганизмы. А — за счет работы осмотических сил растение создает гидростатическое давление Δp, которое гонит воду вверх по стволу; Б — все живые организмы производят механическую работу. Белка увеличивает свою потенциальную энергию Eпот, поднимая свое тело массой m вверх по стволу на высоту h и преодолевая при этом ускорение силы тяжести g; В — электрические рыбы (на рисунке изображен электрический угорь) создают напряжение сотен вольт; Г — в светлячке "спрятаны микроорганизмы, испускающие свет, энергия квантов которого равна произведению постоянной Планка h на частоту ν
Анатомическое изучение скелетной мышцы показывает, что она состоит из многих длинных мышечных волокон, так называемых миофибрилл. Под микроскопом, особенно в поляризованном свете, эта структура представляет собой чередующиеся темные и светлые полосы. На основании электронно-микроскопических исследований разработана следующая модель молекулярного строения мышцы: к прочным поперечным стенкам, так называемым пластинкам Z, в строгом порядке прикреплены нитевидные белковые молекулы. Эта структура напоминает щетку с гексагонально расположенными щетинками. Расстояние между
отдельными "щетинками" намного больше их диаметра. Теперь представим себе две щетки, размещенные так, что их щетинки находятся друг против друга. Щетинки не должны касаться друг друга, но расстояние между их кончиками должно быть меньше их длины. Теперь между щетинками, параллельно им, поместим спички так, чтобы их концы оказались на разных щетках. Модель готова. Щетинки изображают тонкие актиновые нити, а спички — лежащие между ними толстые нити молекул миозина. Щетки могут сдвигаться относительно друг друга, при этом спички будут больше или меньше" углубляться в область между щетинками.
В мышце происходит интенсивное взаимодействие между молекулами актина и миозина, которое регулируется обменом веществ. Между этими параллельно расположенными молекулами образуются мостики — поперечные связи. Эти связи имеют химическую природу и возникают только между совершенно определенными участками молекул, так называемыми активными центрами. Под действием энергии, вырабатываемой в процессе обмена веществ, например при участии уже неоднократно упоминавшегося аккумулятора химической энергии — аденозинтрифосфата (АТФ), места соединений мостиков с молекулами непрерывно меняются; миозиновая молекула тянет молекулу актина, подобно тому как группа рыбаков вытягивает сеть. Актиновые нити, увлекаемые поперечными связями, скользят между нитями миозина — мышца сокращается. Эта гипотеза скользящих нитей в настоящее время рассматривается как наиболее правдоподобное объяснение механизма мышечного сокращения.
 Строение скелетной мышцы и ее модель. Мышца состоит из миофибрилл (А), которые под микроскопом выглядят как чередующиеся светлые и темные полосы (Б). Модель миофибриллы можно представить следующим образом: между тонкими нитями молекул актина, расположенными в строгом геометрическом порядке, лежат толстые нити молекул миозина (В). Порядок расположения нитей показан на поперечном сечении миофибриллы (Г). Сокращение мышцы можно объяснить скольжением этих нитей относительно друг друга, которое приводит к сближению пластинок Z
Строение скелетной мышцы и ее модель. Мышца состоит из миофибрилл (А), которые под микроскопом выглядят как чередующиеся светлые и темные полосы (Б). Модель миофибриллы можно представить следующим образом: между тонкими нитями молекул актина, расположенными в строгом геометрическом порядке, лежат толстые нити молекул миозина (В). Порядок расположения нитей показан на поперечном сечении миофибриллы (Г). Сокращение мышцы можно объяснить скольжением этих нитей относительно друг друга, которое приводит к сближению пластинок Z
Однако не будем задерживаться на этом очень: специфическом виде работы биологической системы. Пойдем дальше. Об осмотических силах, действующих в живом организме, известно несколько меньше, чем об энергии механического движения. У растений; оба эти явления тесно связаны между собой. Движение листьев мимозы, поворот головки подсолнечника к свету, закрытие цветка вечером — все это механические движения, которые управляются гидравлически, то есть путем изменения содержания воды в некоторых тканях. Основой такого рода гидравлических процессов является движение воды под действием осмотического давления. Классический пример работы, производимой осмотическими силами, — подъем воды от корня растения к вершине. Осмотическую работу клетки можно наблюдать под микроскопом. Инфузории туфельки и другие одноклеточные, обитающие в пресных водах, обладают насосом, подобно аварийной помпе на корабле, предохраняющим их от переполнения водой. Но в отличие от корабля, который в нормальном состоянии водонепроницаем, инфузория наполняется водой непрерывно, так как внутри нее осмотическое давление ниже, чем в окружающей среде. Поступающая вода также непрерывно выкачивается звездчатой сократительной вакуолью. Кстати, это прекрасный пример динамического равновесия на клеточном уровне.
 Работа осмотических сил у одноклеточных. Под действием градиента между внешней (πвнеш) и внутренней (πвнутр) средой вода непрерывно поступает через цитоплазматическую мембрану внутрь инфузории туфельки. Сократительные вакуоли собирают ее, а затем выкачивают
Работа осмотических сил у одноклеточных. Под действием градиента между внешней (πвнеш) и внутренней (πвнутр) средой вода непрерывно поступает через цитоплазматическую мембрану внутрь инфузории туфельки. Сократительные вакуоли собирают ее, а затем выкачивают
Не будем перечислять явления, связанные с процессами преобразования энергии. Их много, а лежащие в их основе механизмы большей частью неизвестны. Однако мы рассматриваем лишь основные принципы преобразования энергии в живом организме. В связи с этим нельзя не упомянуть об электрической энергии, которая наряду с химической играет совершенно особую и очень важную роль в биологических процессах. Здесь в первую очередь, естественно, вспоминаются электрические рыбы: электрический угорь, электрический сом или электрический скат, которые могут наносить чувствительные удары током и тем самым защищаться от врагов. Но каждый из нас имел возможность убедиться в существовании электрических полей и в своем собственном теле. Достаточно назвать ЭКГ — электрокардиограмму, описывающую изменение разности электрических Потенциалов между отдельными частями нашего тела, вызванное деятельностью сердца. Аналогичные электрические поля создают все работающие мышцы тела, а также мозг и органы чувств.
 Электрические рыбы способны вырабатывать импульсы напряжением до нескольких сотен вольт. Такие напряжения они используют в первую очередь для защиты от врагов, поражая тех ударами током. Недавно стало также известно, что короткие электрические импульсы помогают рыбам ориентироваться в мутной воде
Электрические рыбы способны вырабатывать импульсы напряжением до нескольких сотен вольт. Такие напряжения они используют в первую очередь для защиты от врагов, поражая тех ударами током. Недавно стало также известно, что короткие электрические импульсы помогают рыбам ориентироваться в мутной воде
С помощью чувствительных электродов, толщина которых меньше микрометра (мкм), т. е. тысячной доли миллиметра, можно установить, что каждая живая клетка по отношению к внешней среде обладает определенным электрическим потенциалом. Он имеет отрицательное значение и по абсолютной величине оставляет 0,01-0,08 В. Изолятором между этими двумя полюсами (клеткой и внешней средой) служит клеточная мембрана. Это тончайшая, толщиной около 100 Å (0,00001 мм), похожая на мыльный пузырь структура, которая окружает каждую клетку. При возбуждении нервные клетки могут резко изменять величину или даже полярность электрического потенциала мембраны
[8]. В результате возникают нервные импульсы — знаки своеобразной азбуки Морзе, с помощью которых осуществляется связь между органами тела. Однако этим еще далеко не исчерпывается значение электрического потенциала клетки. Под действием разности электрических потенциалов в чрезвычайно тонкой клеточной мембране, как внутри тонкого пластинчатого конденсатора, возникает сильное электрическое поле. Оно определяет свойства мембраны: ее проницаемость для различных веществ, работу ферментов внутри и в непосредственной близости от нее, а также многое другое.
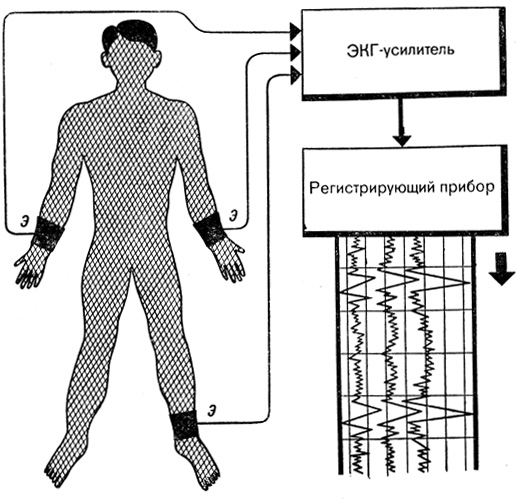 Работающая сердечная мышца создает электрическое поле. Возникающие на поверхности тела электрические потенциалы можно обнаружить с помощью усилителя, чувствительность которого достигает 0,00001 В, и записать в виде электрокардиограммы. Такая электрокардиограмма позволяет врачу судить о работе нашего сердца
Работающая сердечная мышца создает электрическое поле. Возникающие на поверхности тела электрические потенциалы можно обнаружить с помощью усилителя, чувствительность которого достигает 0,00001 В, и записать в виде электрокардиограммы. Такая электрокардиограмма позволяет врачу судить о работе нашего сердца
Таким образом, электрическая энергия в живых огранизмах имеет большое значение как носитель информации, необходимой для работы различных механизмов регуляции и управления. И лишь очень немногие организмы, например электрические рыбы, в процессе многовекового отбора выработали некий механизм, использующий энергию электрического импульса как оружие. В этом случае электрический разряд происходит в результате параллельного и последовательного включения многих элементарных "батарей". Конечно, заманчиво поближе познакомиться с природой таких электрических процессов. Каковы они? Как они возникают? Что ими управляет? Однако мы остановимся только на некоторых основных аспектах этой обширной темы.
 Микроэлектрод, введенный в нервное волокно, дает возможность измерять электрический потенциал клетки. Такой 'потенциал покоя' равен примерно -50 мВ (1 мВ = 0,001 В). При возбуждении нервных клеток мгновенно возникает 'потенциал действия' и потенциал клетки становится положительным
Микроэлектрод, введенный в нервное волокно, дает возможность измерять электрический потенциал клетки. Такой 'потенциал покоя' равен примерно -50 мВ (1 мВ = 0,001 В). При возбуждении нервных клеток мгновенно возникает 'потенциал действия' и потенциал клетки становится положительным
Невольно напрашивается мысль снова провести аналогию с техникой. Мы уже упоминали об "азбуке Морзе биологической связи", "пластинчатом конденсаторе", "электронной обработке данных". Сейчас инженеры действительно очень интересуются этими вопросами, надеясь почерпнуть в живой природе новые идеи для усовершенствования электронных приборов. Следует, однако, указать на очень важное различие между электрическими явлениями в живых организмах и процессами, протекающими в электронных приборах. В электронике в качестве проводников электрического тока используют металлы или кристаллические полупроводники. Ток в них представляет собой поток электронов, т. е. мельчайших, почти не обладающих массой частиц. Биологические системы лишены металлических участков, у них обнаружены только небольшие области, где могут возникать такие же эффекты, как в органических полупроводниках. Следовательно, электрический ток в клетке не может быть потоком электронов, это должен быть ток, способный проходить через ионные растворы. Ионы, т. е. положительно или отрицательно заряженные атомы и молекулы, переносят заряды, создавая тем самым ток и электрические потенциалы. Основная роль в этом процессе принадлежит, естественно, самым маленьким, и, следовательно, самым быстрым ионам. Это ионы натрия, калия и хлора. Заметим, кстати, что в электронике тоже сплошь и рядом используют ионную проводимость. Хороший пример тому — электролитический конденсатор. На проводимости электролитических растворов основана специальная отрасль электроники, называемая хемотроникой.
 Окружающая клетку исключительно тонкая мембрана состоит из жироподобных липидов и белков, которые являются хорошими изоляторами, и снабжена порами. Она служит одновременно изолятором, сопротивлением и батареей. По обе стороны мембраны находятся ионные растворы различного состава
Окружающая клетку исключительно тонкая мембрана состоит из жироподобных липидов и белков, которые являются хорошими изоляторами, и снабжена порами. Она служит одновременно изолятором, сопротивлением и батареей. По обе стороны мембраны находятся ионные растворы различного состава
Может ли хемотроника живой клетки служить образцом, которому следует безоговорочно подражать? Не совершили ли инженеры ошибки, взяв за основу развития электроники электронную проводимость? С полной уверенностью можно утверждать, что, хотя хемотроника уже на заре своего развития приобрела большое значение, она никогда не вытеснит традиционную электронику. Только в немногих специальных областях можно надеяться получить с помощью хемотроники более качественные и экономичные электронные устройства.
Развитие жизни на основе водных растворов обусловлено, конечно, не выбором оптимального принципа электропроводимости. Проблемы выбора здесь вообще не существует, так как электронная проводимость предполагает наличие металлических проводников, а живое существо из металла абсурдно с молекулярно-биологической точки зрения. Поэтому развитие жизни неизбежно оказалось связанным с ионной проводимостью. Хемотроника имеет бесспорные недостатки по сравнению с электронной техникой. Они заключаются главным образом в том, что скорость протекания процессов в этом случае ограничена. Нервный импульс распространяется в течение миллисекунды — тысячной доли секунды. Скорость о распространения не может быть выше, поскольку ионы гораздо крупнее и медлительнее электронов. Хемотроника не успевает за электроникой, которая сейчас оперирует уже наносекундами (0,000 000 001 с). И все же наш мозг работает быстрее, экономичнее и надежнее, чем самая совершенная ЭВМ. И пока это остается загадкой, которая больше всего поражает воображение специалистов по бионике.
Как же возникает разность электрических потенциалов на мембране клетки? В предыдущей главе мы уже рассматривали клетку как систему, находящуюся в состоянии динамического равновесия: ионы калия "закачиваются" внутрь клетки из внешней среды вопреки знаку градиента их концентрации и стремятся снова выйти из нее под действием этого градиента. В отношении ионов натрия наблюдается обратная картина: ионный насос постоянно "заботится" о том, чтобы концентрация ионов натрия внутри клетки была меньше, чем во внешней среде. Посмотрим еще раз на рисунок, приведенный в предыдущей главе. Засасываемый внутрь клетки калий стремится выйти наружу, выкачиваемый из нее натрий стремится снова проникнуть внутрь. Несмотря а это, в клетке по сравнению с внешней средой содержится больше калия и меньше натрия — царит динамическое равновесие.
В электростатике существует строгий закон, который гласит, что для предотвращения возникновения огромных электростатических сил содержание носителей положительных и отрицательных зарядов в растворе всегда должно быть одинаково. Это требование электронейтральности. Следовательно, положительный катион может покинуть клетку лишь в том случае, если он заберет с собой и своего партнера — отрицательный анион. (Слова "положительный" и "отрицательный" как характеристики партнеров означают здесь только знак электрического заряда и ничего более.) И все-таки катионы проникают через мембрану клетки намного легче, чем анионы. Если бы отрицательно заряженный ион хлора по праву, данному ему электростатикой, не тянул назад в клетку своего подвижного партнера — положительно заряженный ион калия, — клетка стала бы бедна катионами. Удерживающие силы велики. Они предотвращают самое худшее.
И все же катион, упорно удерживаемый следующим за ним анионом, побеждает. Он оказывается на наружной стороне мембраны чуть раньше аниона. Этой маленькой разницы уже достаточно, чтобы возник некоторый вполне измеримый электрический потенциал. Так как ионы калия выходят из клетки, а ионы натрия, наоборот, входят в нее, оба эти процесса должны были бы взаимно компенсировать друг друга. Однако этого не происходит, потому что в норме выход калия намного больше, чем приток натрия. Клеточная мембрана так задерживает ионы натрия, что они проникают в клетку даже позже ионов хлора. Таким образом, электрический потенциал клетки определяется диффузией ионов калия. При возбуждении, например, нервных или мышечных клеток клеточные мембраны мгновенно перестраиваются: их проницаемость для ионов натрия значительно возрастает. В этом случае ионом, определяющим величину потенциала клетки, является натрий, который в соответствии со своим направлением диффузии меняет знак потенциала клетки на противоположный. Создается так называемый "потенциал действия" возбужденных клеток, который в отличие от "потенциала покоя" имеет положительный знак, и сама клетка при возбуждении заряжена положительно. Этот наглядный пример дает представление о природе биоэлектрических явлений.
Мы видели, что разность электрических потенциалов между внутренней и внешней поверхностями мембраны определяется состоянием динамического равновесия клетки. Остается нерешенным вопрос: как ионы засасываются в клетку? Решение его очень важно как для теоретической биологии, так и для практики сельского хозяйства и медицины. Ученые упорно работают над этой проблемой, и все же многое остается неясным. Установлено, что для регулирования концентраций ионов внутри клетки используется химическая энергия, в частности энергия аденозинтрифосфата (АТФ). Каким-то образом этот химический аккумулятор приводит в действие ионный конвейер, или, как его еще называют, ионный насос. Выяснение механизма работы такого насоса — одна из важнейших задач сегодняшней молекулярной биологии.
Здесь мы подошли к одной из наиболее интересных областей современной биофизики — биофизике мембран. Ионные насосы нужны не только для создания электрического потенциала. Они необходимы и для транспорта питательных веществ, воды, микроэлементов. Они создают осмотические силы, о которых мы уже говорили. Все транспортные потоки в клетке связаны между собой: один поток влияет на другой, управляет им и при необходимости обеспечивает его энергией. Клеточная мембрана, в которой все это происходит, не просто место действия таких процессов-благодаря своим молекулярным свойствам она активно управляет транспортными потоками и катализирует их взаимодействие.
Затронутые нами вопросы представляют интерес для врача, который прописывает средства для улучшения обмена веществ, следит за нарушением функции всасывания (резорбции) или пытается восстановить деятельность почек больного. Поскольку эти вопросы связаны также с использованием удобрения растениями, они касаются и работников сельского хозяйства. Удивительные возможности биологических мембран вызывают огромный интерес и у инженеров. В химической промышленности вещества разделяются преимущественно путем перегонки. Это сложно и требует значительных затрат энергии. Гораздо выгоднее было бы отделять конечный продукт от исходного вещества или побочных продуктов, используя мембраны с большой поверхностью. При опреснении морской воды, т. е. в области, особенно важной для народного хозяйства, такие мембраны уже успешно применяются.
Возможно, новый толчок теоретической разработке этих вопросов даст термодинамика необратимых процессов. Это новое современное направление термодинамики развилось в последнее десятилетие на основе классической термодинамики обратимых процессов, о которой мы уже кратко говорили. Однако задачи и математический аппарат данной области науки слишком специальны, чтобы обсуждать их в нашей книге.
Что принесло нам знакомство с механизмом живого? Это был лишь тот беглый взгляд, который бросает часовых дел мастер на впервые попавший в его руки механизм. Мы старались рассмотреть и обсудить только самое принципиальное. И, конечно, не на все поставленные вопросы смогли дать достаточно полные ответы. Но не стоит огорчаться. Ведь мы не ставили перед собой задачи доктора Вагнера: "Хоть много знаю я, но все хотел бы знать", однако не разделяем мы и пессимизма доктора Фауста
[9]: "... но что значит — знать? Вот в чем затрудненье!" Мы просто хотели возбудить у читателя любопытство, а оно уже может перерасти в творческую любознательность, стремление заняться серьезными научными изысканиями.
В царстве молекул
Макро- и микрофизика. Можно ли видеть молекулы? Каков размер иона? Мембрана клетки в качестве ионного сита. Игра с бильярдными шарами. Что такое статистический клубок? Жизнь — Вселенная в хаосе. Как она могла возникнуть? Жизнь означает упорядоченность; хаос можно рассчитать, а обуздав и направив, — использовать. Энергия хаотического движения молекул + биологическая информация = биологическая функция. Большие числа и малые пространства. Почему клетки не меньше, чем они есть? Треугольник пространства, времени и надежности в молекулярной биологии. Упорядоченные поля.
Мы начали книгу с описания крупных видимых объектов, которые можно наблюдать в повседневной жизни: летящего насекомого, качающегося стебелька, неуклюжего слона. Нам удалось уточнить и расширить наши знания и представления, многое связать воедино и объяснить. Одних вещей мы коснулись лишь вскользь, другие потребовали более серьезного обсуждения; кое-где нам пришлось глубже проникнуть в теорию и от конкретных жизненных понятий перейти к абстракциям.
Теперь нам придется расстаться с привычными представлениями и приготовиться многое осмыслить заново. Мы попытаемся проникнуть в царство молекул. Это означает не просто изменение масштабов, т. е. уменьшение наблюдаемого объекта или мысленное увеличение предмета нашего исследования, это означает прежде всего, что перед нами открывается качественно новый мир — мир микрофизики. Там скрыты корни жизни. Различные особенности живых организмов, которые можно легко заметить, например появление чего-то нового в окраске, форме или манере поведения, связанного с изменением наследственности, возникновение определенных болезней, таких, как рак, заболевания крови и т. д., обусловлены изменением молекул. Это одна из причин того, что молекулярная биология и биофизика приобретают в последнее время все большее значение.
Рассказывают, что один архитектор поручил своим ученикам спроектировать дома и квартиры для людей, совсем не похожих на нас: они ходят на четырех ногах, голова у них сбоку и т. д. Что это, бессмысленная прихоть? Отнюдь нет. Будущие архитекторы должны уметь забывать о привычном, должны обладать способностью отойти от традиции и взглянуть на мир новыми глазами.
Однако тот мир, который теперь перед нами откроется, — это не фантастика, ибо современная физика уже разработала для его описания достаточно точные научные представления и понятия. Но прежде чем вступить в царство молекул, мы должны сдать в архив наш привычный повседневный опыт. Теперь нашими проводниками будут учебники статистической физики и квантовой механики, где изложено все, что известно физикам на сегодняшний день. А они неплохо ориентируются в этой области!
С жадным любопытством вступаем мы в новый мир и нетерпеливо спрашиваем: как выглядит белковая молекула, о которой мы слышали столько удивительного? Правда ли, будто она катализирует процессы преобразования энергии и вещества, переносит электроны, совершает механическую работу и многое другое? Увы, нас ждет разочарование, так как наш первый вопрос лишен смысла: молекула вообще не "выглядит!"
Мы привыкли к тому, что все окружающее можно видеть и как-то описать. У твердых тел мы замечаем цвет, форму и размер, у жидкостей — по меньшей мере, цвет и консистенцию. Мышь серая и имеет хвост длиной примерно 15 см. И хотя есть мыши коричневые и белые, большие и маленькие, окраску и размеры каждой конкретной мыши мы можем установить однозначно. Если вскрыть мышь, то в ее голове мы найдем мозг, который тоже имеет определенные форму, размеры и окраску. Мы обнаружим разные области мозга, а вооружившись лупой, можем отчетливо разглядеть главные нервы. Специальными методами приготовим срезы мозга и рассмотрим их под микроскопом. Мы увидим разветвления нервных волокон и органеллы клеток: ядро, аппарат Гольджи, митохондрии. Чтобы различить последние, нам понадобится сильный микроскоп с 1300-кратным увеличением. Но на этом наши возможности "видеть" кончаются.
 Различные устройства обеспечивают разную степень увеличения биологических объектов. Возможности 'видеть', в буквальном смысле этого слова, мы лишаемся уже на уровне оптического микроскопа. Применительно к электронному микроскопу, обладающему самой высокой разрешающей способностью, более подходит понятие 'отображать'
Различные устройства обеспечивают разную степень увеличения биологических объектов. Возможности 'видеть', в буквальном смысле этого слова, мы лишаемся уже на уровне оптического микроскопа. Применительно к электронному микроскопу, обладающему самой высокой разрешающей способностью, более подходит понятие 'отображать'
И дело здесь не в оптической промышленности, которая не создала микроскопов с большей разрешающей способностью. Причина кроется в свойствах света, который мы используем для наблюдения. У синего, еще видимого, света длина волны 0,4 мкм. Вблизи этой границы и. лежит максимум разрешающей способности оптического микроскопа. Точно так же как нельзя заточить карандаш топором, невозможно "видеть" объекты, размеры которых меньше, чем длина волны используемого света.
Ну и что же? Ведь всем известно, что существуют электронные микроскопы. Так называют приборы, которые показывают на экране из сульфида цинка во много раз увеличенную картину распределения электронной плотности очень малых объектов. Это досадно, потому что слово "микроскоп" мы привыкли связывать со словом "видеть". А в электронном микроскопе, если пучок электронов преобразовать в световой пучок, нам удается только получить увеличенное изображение на освещенном экране. Тогда в лучшем случае можно сказать, что мы при помощи некой сложной методики отобразили какую-то чрезвычайно мелкую структуру в сильно увеличенном виде. Но о том, чтобы "видеть" ее, не может быть и речи!
 Пластмассовая модель клеточной структуры, увеличенной в 10 000 раз. Она наглядна и помогает нам лучше представить реальный объект. Но не следует принимать эту модель за действительную структуру
Пластмассовая модель клеточной структуры, увеличенной в 10 000 раз. Она наглядна и помогает нам лучше представить реальный объект. Но не следует принимать эту модель за действительную структуру
Может быть, мы просто придираемся к словам? К сожалению, нет. Человек склонен переносить знания, добытые им при изучении одного процесса или объекта, на другие еще не известные ему процессы и объекты, которые он хотел бы исследовать. Сам объект заставляет его исправлять прежние неправильные представления и тем самым обогащает его, расширяет круг его знаний. Ученый, занимающийся молекулярной биологией, "видит" картины, увеличенные в 10 000 или 100 000 раз, видит со всей четкостью и, вероятно, строит на основании этого какие-то осязаемые модели, например из пластмассы. Теперь перед ним модель и, забыв о действительных размерах объекта, он пытается применять известные ему законы физики, скажем первый закон Ньютона: тело сохраняет состояние покоя, если на него не действует никакая сила, изменяющая это состояние. Этот закон, безусловно, правильный, и пластмассовая модель перед глазами биолога лежит на столе совершенно спокойно. Реальная структура, размеры которой в десять тысяч раз меньше, тоже лежала бы очень спокойно, если бы на нее не действовали никакие силы. Но материя не может существовать без движения; эту структуру непрерывно "толкают" со всех сторон колеблющиеся, вращающиеся, движущиеся молекулы окружающей среды, в непрерывном движении находятся и молекулы, из которых состоит сама структура. Модель из пластмассы спокойно стоит на столе, а реальная структура колеблется, вращается и ни минуты не остается в покое.
 Используя дифракцию рентгеновских лучей, можно определить внешний вид биологической макромолекулы. Однако этот метод связан с большими трудностями. Пока известны структурные модели только очень немногих молекул
Используя дифракцию рентгеновских лучей, можно определить внешний вид биологической макромолекулы. Однако этот метод связан с большими трудностями. Пока известны структурные модели только очень немногих молекул
То, что справедливо для очень малых, но все же состоящих из многих молекул объектов, в еще большей степени относится к самим молекулам. Строение молекулы бессилен отобразить даже электронный микроскоп. Для того чтобы получить представление о структуре биологических макромолекул, используют метод дифракции рентгеновских лучей (рентгеноструктурный анализ). У рентгеновских лучей длина волны составляет несколько ангстрем (1 Å = 10
-8см), и, следовательно, с их помощью еще можно получить "изображение" молекулярных структур. Узкий пучок рентгеновских лучей, проходя через возможно более тонкий слой макромолекулярного вещества, к примеру белка, дает на фотопластинке картину дифракции, которая непосвященному показалась бы бессмысленной. Места почернения на фотопластинке очень тщательно промеряют и обсчитывают. В результате получают картину распределения плотности электронов в молекуле. Она напоминает карту гористой местности. На основании такой карты физик строит модель молекулы. Здесь речь уже не идет о какой-то зафиксированной, "замороженной" картине, как в случае электронной микроскопии, однако и эта модель не дает нам четкого представления о динамической структуре молекул. И хотя при рентгеноструктурном анализе движение молекул не "замораживается", то, что мы видим, можно считать неким средним значением для очень многих молекул. Таким образом, модель молекулы своей неподвижностью напоминает манекен, застывший в витрине магазина.
 Модель молекулы, составленная из полушарий. Такие модели таят в себе опасность вступить на позиции механицизма
Модель молекулы, составленная из полушарий. Такие модели таят в себе опасность вступить на позиции механицизма
Мы должны твердо усвоить: "видеть" молекулу нельзя, о "внешности" молекулы не может быть и речи. Вместо "портрета" молекулы мы получаем ее научно обоснованное описание, которое отнюдь не идентично знакомым нам картинам из видимого и привычного мира макрофизики.
Итак, молекула представляет собой динамическую структуру. Но можно ли, основываясь на модели, полученной в результате рентгеноструктурного анализа, определить хотя бы усредненные размеры и. форму молекулы? Да, конечно, однако и здесь существуют свои трудности. Проще всего их продемонстрировать на примере атомов или ионов, т. е. атомов, которые, потеряв или присоединив то или иное число электронов, несут электрический заряд. Ранее мы уже обсуждали значение этих носителей заряда, в частности ионов натрия и калия, которые имеют положительный заряд и называются катионами. Каков размер иона?
Мы привыкли к тому, что размеры окружающих нас предметов и расстояния между ними можно измерять различными способами. Например, длину спортивной площадки можно либо измерить рулеткой, либо определить с помощью оптического дальномера. В принципе мы можем также заметить время, за которое звук пройдет измеряемый отрезок, а затем, зная скорость звука, рассчитать длину отрезка (примерно так, засекая момент вспышки молнии, а затем раската грома, на основании разницы во времени определяют в грозу отдаленность разрядов). И хотя в зависимости от применяемых методов точность измерения длины площадки будет различной, результат измерений всегда окажется одинаковым.
Однако сейчас нам следует забыть наш повседневный опыт. Он не подтверждается на молекулярном и атомном уровнях. Если мы определим радиус иона разными методами, то получим совершенно различные результаты. Рассмотрим следующий пример. В кристалле поваренной соли ионы натрия и хлора "упакованы" в строгом порядке, как елочные шары в коробке. С помощью рентгеноструктурного анализа можно очень точно установить расстояние между центрами ионов. В результате мы получаем так называемый кристаллический радиус иона; например, у натрия он составляет 0,95 Å.
Теперь попытаемся измерить размеры иона другим способом. Когда шар проходит через жидкость, его скорость зависит от величины действующей на него силы, вязкости жидкости и радиуса шара. Если ионы поместить в раствор между двумя электродами, они будут двигаться в электрическом поле. Этот процесс называют электрофорезом. Скорость движения ионов можно определить, зная электрическое сопротивление раствора. Вязкость воды тоже известна. Таким образом, можно рассчитать радиус иона. С удивлением мы обнаруживаем, что для того же иона натрия радиус равен 1,85 Å, т. е. вдвое больше кристаллического радиуса. Что это, ошибка измерения? Ничего подобного. Просто мы измерили совсем другой параметр. Его называют гидратационным радиусом. Ион притягивает воду и движется, облаченный в водяную рубашку. И следовательно, "ошибка" заключается в том, что в этом случае мы как бы измеряем диаметр елочного шара, завернутого в бумагу. В действительности дело обстоит гораздо сложнее, ибо ионы не новогодние шары и у них нет твердой поверхности. Даже если в очень грубом приближении можно представить ядро и электроны как материальные точки, то ион в целом следует рассматривать как область действия сильнейших электрических полей. Что же тогда мы измеряем?
 Какое размер иона? К иону не приложишь штангенциркуль, Здесь можно определить только эффективные радиусы или диаметры, т. е. параметры, измеряемые в совершенно конкретных условиях
Какое размер иона? К иону не приложишь штангенциркуль, Здесь можно определить только эффективные радиусы или диаметры, т. е. параметры, измеряемые в совершенно конкретных условиях
Нам просто пора привыкнуть рассматривать величины как "эффективные" параметры, т. е. значения, измеряемые в совершенно конкретных условиях, как это было показано на примере определения кристаллического и гидратационного радиусов. Для биологических систем важен именно гидратационный радиус. Ведь в живых системах ионы находятся в водной среде и поэтому окружены водяной рубашкой. Но эта водяная рубашка не имеет никакой четкой границы, подобно тому как нигде не обрывается резко атмосфера Земли. И хотя эффективный гидратационный радиус удается определять очень точно, с хорошей воспроизводимостью результатов измерений, тем не менее не стоит рассматривать его как некую абсолютную величину.
Чтобы пояснить эту мысль, рассмотрим следующий пример. До сих пор остается не совсем ясным, каким образом клеточная мембрана обеспечивает избирательную проницаемость для различных ионов. В предыдущей главе мы говорили о том, что калий проходит через мембрану гораздо легче, чем натрий. Для объяснения этого эффекта в свое время была выдвинута так называемая "теория ионного сита", которая сейчас вновь привлекает внимание исследователей. Первоначальная идея выглядела очень простой и даже механистической.
Если представить мембрану в виде плоскости с отверстиями (порами) определенной величины, а ионы как шары соответствующего диаметра, то вся структура в делом будет напоминать сито, с которым дети играют в песочнице. Но ионы — это не обычные отполированные шары, а отверстие диаметром в несколько ангстрем нельзя сравнить с тем, которое оставляет сверло в куске фанеры. Мы уже отмечали, что размеры ионов характеризуются эффективными величинами; то же справедливо и для пор. И если попытаться нарисовать мир молекул более реально, то картину диффузии ионов через мембрану можно описать следующим образом: мелкие частицы проходят через маленькое отверстие, конечно, легче, чем более крупные, но в отличие от твердого шара диаметром, скажем, 4,3 см, который при всем желании не протолкнуть через отверстие диаметром 4,1 см, ион с гидратационным диаметром 4,3 Å может пройти через пору с эффективным диаметром 4,1 Å, если приложенная к нему сила достаточно велика. На основе этих представлений вместо теории избирательной проницаемости (по принципу "все или ничего"), оказавшейся ошибочной, была разработана теория частичной проницаемости (по принципу "больше или меньше").
 Так схематически можно изобразить избирательную проницаемость мембраны для ионов с позиций 'теории ионного сита' в ее механистическом представлении. Малые по размерам ионы калия проходят через отверстия, а ионы натрия в толстой водяной рубашке — нет. Знакомство с физическим смыслом понятия 'размер иона' показывает ограниченность этой теории
Так схематически можно изобразить избирательную проницаемость мембраны для ионов с позиций 'теории ионного сита' в ее механистическом представлении. Малые по размерам ионы калия проходят через отверстия, а ионы натрия в толстой водяной рубашке — нет. Знакомство с физическим смыслом понятия 'размер иона' показывает ограниченность этой теории
Итак, нет таких нелепых вопросов, которые не научили бы нас чему-нибудь. Если теперь мы поняли, почему нельзя видеть молекулы и ионы, если мы хоть что-то узнали об этих мельчайших частицах вещества, составляющих все живое, то первую нашу экскурсию в таинственный мир молекул можно считать успешной. Правда, наши познания, которые можно было бы применить к биологии, еще так малы, что нам придется продолжить путешествие в микромир.
Теперь мы займемся новой проблемой — движением материи. Как мы уже говорили, хотя и можно построить модель молекулы, которая будет спокойно лежать перед нами на столе, реальная молекула находится в непрерывном движении. Как оно происходит? Каково его значение для наших рассуждений?
Начнем опять с самого наглядного — модели, взятой из повседневной жизни. Ввинтим в потолок крючки в произвольном порядке, но не слишком далеко друг от друга и подвесим к ним на тонких нитях бильярдные шары так, чтобы они висели на одинаковой высоте достаточно близко друг к другу, но не соприкасаясь. Закроем двери и окна, чтобы никакое движение воздуха нам не мешало, и подождем, пока шары не перестанут раскачиваться. Вот теперь и начнем эксперимент. Сильно толкнем один из шаров в произвольном направлении. Шар не улетит далеко, вскоре он столкнется с соседним шаром. Бильярдные шары изготовляют из качественной древесины, поэтому их соударение будет упругим. Под упругим в физике понимают такой удар, когда импульс полностью передается от одного тела к другому:
 При упругом ударе импульс одного тела передается другому. Это легко увидеть на примере бильярдных шаров. Шар, который первоначально двигался и произвел удар, останавливается, а шар, получивший удар, приходит в движение
При упругом ударе импульс одного тела передается другому. Это легко увидеть на примере бильярдных шаров. Шар, который первоначально двигался и произвел удар, останавливается, а шар, получивший удар, приходит в движение
Если бы первый шар не висел на нити, подобно маятнику, он сразу бы остановился. То, что он продолжает раскачиваться, безусловно, некоторый недостаток нашей модели, но им можно пренебречь. В любом случае второй шар, приобретя тот же импульс, что имел первый, будет двигаться до тех пор, пока не столкнется со следующим шаром. Итак, первоначальный импульс передается в нашей системе от одного шара к другому. Но не всегда происходит прямое попадание. Часто два шара сталкиваются так, что импульс передается от одного к другому лишь частично. Легко представить, что очень скоро в движение окажутся вовлеченными все шары нашей системы.
Как долго оно будет продолжаться? Это зависит от массы шаров и толщины нитей. Однако в какой-то момент обязательно наступит покой: шары перестанут раскачиваться и неподвижно повиснут на нитях. Но куда же пропала энергия? Ведь она не может бесследно исчезнуть! В то же время ее нет. Точный термометр покажет нам, что температура в комнате (конечно, при условии ее идеальной термоизоляции) слегка повысилась. Механическая энергия превратилась в тепло. Это произошло из-за движения воздуха, а также потому, что шары не являются абсолютно упругими, а обладают некоторой пластичностью.
Теперь настало время, задуматься, в чем смысл этого эксперимента. Что он означает? Какие выводы отсюда можно сделать? Молекулы, атомы, ионы, из которых состоит воздух в комнате, подвижны. Они движутся, сталкиваются, обмениваются друг с другом импульсами. Таким образом, мы получили грубую модель молекулярной системы, правда двумерную, в плоскости, параллельной потолку комнаты. Подобно тому как маятники двигались в плоскости, молекулы могут двигаться в трехмерном пространстве. Однако нам следует твердо усвоить, что молекулы не миниатюрные бильярдные шары, не тела с твердой и гладкой, четко очерченной поверхностью. И все-таки при соударении молекулы ведут себя как обычные макроскопические тела, хотя в случае молекул "сталкиваются" не твердые поверхности, а электрические поля, которые и вызывают силы отталкивания.
 Если в механической системе один шар сталкивается с другими, импульс 'путешествует' по системе. При центральном ударе импульс передается полностью, при боковом — возникает рикошет: ударяющий шар, передав партнеру часть импульса, отклоняется от первоначального направления движения. Через небольшой промежуток времени все шары системы приходят в движение
Если в механической системе один шар сталкивается с другими, импульс 'путешествует' по системе. При центральном ударе импульс передается полностью, при боковом — возникает рикошет: ударяющий шар, передав партнеру часть импульса, отклоняется от первоначального направления движения. Через небольшой промежуток времени все шары системы приходят в движение
Остановимся на этом вопросе поподробнее. Итак, хотя молекулы и атомы нельзя уподобить бильярдным шарам, они соударяются аналогично последним и передают импульс другим молекулам. Но, чтобы не попасть впросак, мы должны быть предельно осторожными, пытаясь перенести макрофизические явления в микромир. Прежде всего интересно выяснить, приходят ли через некоторое время молекулы, подобно бильярдным шарам, в состояние покоя? Как мы уже говорили, часть энергии бильярдных шаров в описанном ранее эксперименте затрачивается на то, чтобы привести в движение воздух. Когда речь идет о молекулах, это движение не играет роли, ибо молекулы — мельчайшие частички вещества, и они колеблются не в воздухе, а в вакууме. Если же молекулы находятся в растворе, то их можно рассматривать как некую "смесь" шаров различного типа, одни из которых представляют растворенное вещество, а другие — растворитель. В растворе молекулы растворенного вещества обмениваются импульсами как между собой, так и с молекулами растворителя; никакого препятствия, подобного воздуху между бильярдными шарами, для них не существует.
А как обстоит дело с потерей энергии, обусловленной пластичностью? Хотя в буквальном смысле нельзя говорить о пластичности молекул, определенное изменение состояния молекулы при ударе в принципе возможно. Молекула может двигаться в том или ином направлении, вращаться вокруг собственной оси, могут также вращаться ее отдельные части или внутри нее возникать колебания. В худшем случае часть молекулы может даже отделиться. С каждым из этих движений связаны соответствующие формы энергии, которые можно рассматривать как составные части общей кинетической энергии молекулы.
Почему в отличие от бильярдных шаров молекулы не нагреваются? Просто потому, что понятие теплоты здесь не совсем применимо. Теплота, температура — это понятия макрофизики. Теплота есть проявление кинетической энергии системы, включающей в себя множество молекул. Горячее тело состоит из чрезвычайно энергично движущихся и сталкивающихся частиц, а в холодном теле движение таких частиц замедлено. В системе с бильярдными шарами энергия движения шаров за счет трения и неупругих соударений превращалась в тепловое движение молекул, а в мире молекул энергия их движения и есть, собственно, теплота. И если теплоту не отводить от молекулярной системы, молекулы будут двигаться бесконечно.
С поступательным, т. е. тепловым, движением молекул, как и с другими видами их движения, связан целый ряд физических вопросов. Эти вопросы интересны и с биофизической точки зрения, однако мы коснемся только некоторых из них. Могут ли скорости поступательного движения, вращения или колебания молекулы меняться непрерывно, подобно тому как меняются скорости движения макротел? Конечно, нет! Это также одна из особенностей микрофизики. Благодаря открытию Макса Планка стало известно, что в микромире энергия может изменяться только отдельными порциями, квантами, которые неделимы. Можно, например, взвесить 1 кг сахара; 1 г; 1/100 г, или, что то же самое, 10
-2 г; 10
-4 г; 10
-8 г и т. д. Однако это "и т. д." имеет свой предел. Если бы мы захотели взвесить 10
-22 г, т. е. 0,0000000000000000000001 г, нам пришлось бы дробить молекулу сахара, и из этого ничего бы не вышло, ибо разрушенная молекула сахара — уже не сахар.
Аналогично обстоит дело и с энергией. Когда речь идет о движении макроскопического тела, его энергия выражается таким большим числом квантов, что уменьшение или увеличение этой энергии на один квант измерить нельзя. Следовательно, величина энергии большого тела изменяется практически непрерывно. С достаточно хорошим приближением сказанное справедливо и для поступательного движения отдельных молекул. Однако при других видах движения молекул, таких, как вращение и колебание, непрерывное изменение энергии невозможно. В этих случаях изменение энергии на один квант уже приводит к значительному изменению скорости движения. С изучением таких процессов связано целое направление биофизики, так называемая квантовая биофизика, которая охватывает вопросы фотосинтеза, природы зрения, действия излучений на вещество и т. д.
А теперь рассмотрим еще один вопрос. Длинную молекулу можно сравнить с цепочкой, каждое звено которой подвижно. Что произойдет с такой цепочкой при хаотическом тепловом движении? Если на молекулу не действуют никакие дополнительные силы, она закручивается в беспорядочный "статистический клубок". Каковы размеры такого клубка при данной длине цепочки и насколько ее концы удалены друг от друга? Как ведет себя такой клубок при движении?
 Представим себе длинную линейную молекулу, состоящую из химически связанных между собой атомов. Углы между отдельными связями остаются постоянными, хотя сама связующая ось искривлена. В результате хаотического тепловогодвижения эта молекулярная цепочка может достаточно сильно перекрутиться (А). Наиболее вероятное состояние молекулы при таком случайном движении — 'статистический клубок' (Б)
Представим себе длинную линейную молекулу, состоящую из химически связанных между собой атомов. Углы между отдельными связями остаются постоянными, хотя сама связующая ось искривлена. В результате хаотического тепловогодвижения эта молекулярная цепочка может достаточно сильно перекрутиться (А). Наиболее вероятное состояние молекулы при таком случайном движении — 'статистический клубок' (Б)
Опять новые вопросы! Пожалуй, нам грозит опасность безнадежно запутаться в лабиринте науки. А этого бы не хотелось. Так будем верны нашему правилу — рассматривать не все, а лишь самое главное, самое интересное. Мы, хотя и не без некоторых усилий, усвоили основные законы микрофизики, тонкости, присущие миру молекул. Мы узнали много удивительного, трудно постижимого.
Какая же картина предстала перед нами? Отсутствие четких границ у молекул и ионов, их непрерывное движение и взаимное превращение, сравнимое с постоянно волнующимся морем, — какой-то хаос! В этом хаосе возник и существует "космос" жизни — микрокосм биологической организации! Как это произошло? Этот вопрос зародился столь же давно, как и способность людей сознательно мыслить, но до сих пор его нельзя считать окончательно решенным. Как же возникла жизнь из хаоса, как могла она сохраниться?
 В молекуле белка строительные кирпичики (аминокислоты) следуют друг за другом в определенном порядке. Эту последовательность аминокислот называют 'первичной структурой' белка. (Символами обозначены отдельные аминокислоты: например, Гли — глицин.)
В молекуле белка строительные кирпичики (аминокислоты) следуют друг за другом в определенном порядке. Эту последовательность аминокислот называют 'первичной структурой' белка. (Символами обозначены отдельные аминокислоты: например, Гли — глицин.)
Мы знаем, что жизнь в высшей степени связана с упорядоченностью и организацией. В строгом порядке выстроены аминокислоты — кирпичики, из которых состоит белок. Их порядок, или, выражаясь языком молекулярной биологии, последовательность, определяет функцию белка, например его высокоспецифическое действие как фермента, т. е. катализатора биохимических реакций совершенно определенного типа. Если последовательность таких кирпичиков нарушится, фермент будет так же мало пригоден к работе, как телевизор, собранный обезьяной.
Итак, аминокислоты связаны между собой настолько прочно, что тепловое движение молекул не в состоянии их перемешать. Но ведь должны же они где-то соединяться друг с другом? "Сборочным цехом", где создаются белковые молекулы, являются рибосомы. Здесь по твердому плану — точной копии с наследственного вещества, хранящегося в кладовой каждой клетки — клеточном ядре, одна аминокислота "сшивается" с другой. На деталях этого процесса мы не будем подробно останавливаться. При желании читатель может в этом разобраться, обратившись за помощью к какой-либо из множества книг по молекулярной биологии. Здесь же для нас важно уяснить, что упорядоченность рождается упорядоченностью. В определенных случаях биологическая организация способна защититься от хаоса, сохранить упорядоченность и передать ее по наследству.
Невольно напрашивается вопрос, как возникла такая упорядоченность. Конечно, умение скопировать план конструкции и построить ее в полном соответствии с этим планом — большое мастерство. Но насколько сложнее разработать такие планы!
 Синтез белковой молекулы Б осуществляется на рибосомах. Порядок расположения элементов наследственного вещества, представленный его копией — молекулой мРНК, определяет последовательность аминокислот в белке. Различные аминокислоты показаны символическими значками. Маленькие молекулы тРНК доставляют к соответствующему коду на мРНК нужные аминокислоты и 'пришивают' их на рибосоме к образующейся белковой молекуле
Синтез белковой молекулы Б осуществляется на рибосомах. Порядок расположения элементов наследственного вещества, представленный его копией — молекулой мРНК, определяет последовательность аминокислот в белке. Различные аминокислоты показаны символическими значками. Маленькие молекулы тРНК доставляют к соответствующему коду на мРНК нужные аминокислоты и 'пришивают' их на рибосоме к образующейся белковой молекуле
В настоящее время у нас уже сформировались достаточно обоснованные представления о том, как благодаря изменчивости и отбору мог появиться план анатомически-морфологического строения организма, как возник первый строительный план.
Путь перехода от неорганической молекулы к органической без участия живых организмов нам сейчас уже в основном ясен. Благодаря космическим полетам удалось обнаружить следы органических соединений и за пределами Земли. Это ознаменовало первый успех в научном объяснении возникновения жизни. Но даже если представить себе целое море органических молекул, например аминокислот, остается непонятным, как появился первый белок, первая молекула, способная к размножению. На карте наших знаний здесь до сих пор простирается огромное белое пятно. По-видимому, нам еще не известны какие-то важные закономерности, ибо создание "разумной" последовательности аминокислот вряд ли могло быть случайностью. Жизнь тогда была бы в высшей степени невероятным явлением.
Теперь остановимся на борьбе жизни с хаосом, борьбе биологической упорядоченности с тепловым движением молекул. Всегда ли тепловое движение ведет к разрушению? В принципе — да, ибо случай творит то, что наиболее вероятно, а самое вероятное — это неупорядоченность. Каждый может в этом убедиться. Достаточно набрать горсть кубиков, а затем бросить их. Очень редко случается так, чтобы один кубик аккуратно лег около другого! А упадут ли хоть раз рядом друг с другом три кубика? Такая упорядоченность, добиться которой в состоянии даже двухлетний ребенок, если и может возникнуть случайно, то лишь с ничтожнейшей вероятностью. Впрочем, это уже положение второго начала термодинамики, о котором мы говорили в предыдущей главе.
Но всегда ли хаотическое движение молекул действительно враждебно жизни? Дальше мы убедимся, что такое движение отнюдь не играет роли лишь разрушающего фактора в различных проявлениях жизни — без теплового движения молекул жизнь вообще была бы невозможна. А затем мы попытаемся решить вопрос: можно ли обуздать случай?
 Броуновское движение мельчайших частиц, видимых под микроскопом. Если мы проследим за такой частицей, то обнаружим, что она движется абсолютно хаотически
Броуновское движение мельчайших частиц, видимых под микроскопом. Если мы проследим за такой частицей, то обнаружим, что она движется абсолютно хаотически
Начнем с наглядного примера. Правда, наглядно только то, что можно видеть, а, как мы уже знаем, само понятие "видеть" на микромир не распространяется. Однако некоторые микрофизические явления можно наблюдать даже с помощью оптического микроскопа.
Полтора столетия назад английский естествоиспытатель Роберт Броун заметил, что частички пыльцы движутся в жидкости очень своеобразно. Даже в препаратах, где жидкость находится в покое, можно наблюдать непрерывное хаотическое движение отдельных частиц во всех направлениях: вперед — назад, вверх — вниз. Легко убедиться, что это явление, позднее названное броуновским движением, вовсе не связано с какими-то особыми свойствами биологического объекта. Мельчайшие частицы грязи, оказавшиеся среди живой пыльцы, также свободно взвешены в жидкости и движутся столь же беспорядочно. В начале нашего столетия Альберт Эйнштейн дал теоретическое объяснение этому явлению и разработал метод точного его расчета.
Мельчайшая частица — спора гриба, пыльцевое зерно, крохотная бактерия или частица пыли — находится в жидкой среде; Со всех сторон она подвергается бомбардировке молекулами жидкости, которые находятся в состоянии теплового поступательного движения (так мы его назвали). В какой-то момент времени сумма ударов, или, как говорят в физике, импульсов, получаемых нашей малой частицей, не равна нулю, они не полностью взаимно уничтожаются. В результате наша частица движется поступательно в некотором направлении. Однако в следующий момент векторная сумма импульсов выглядит уже совершенно иначе. Тогда частица движется в другом направлении. Так направление ее движения непрерывно меняется — то туда, то сюда. В итоге частица остается на одном и том же месте, хотя и совершает колебания в пределах определенного радиуса действия. Как установил Эйнштейн, этот радиус действия тем больше, чем меньше частица, чем выше температура и чем меньше вязкость среды, в которой находится частица. Большие частицы получают такое количество толчков, что последние взаимно уничтожают друг друга, и частица почти не смещается. Как говорится, у семи нянек дитя без глазу.
 Причиной броуновского движения являются толчки, вызванные тепловым поступательным движением молекул. Отдельные импульсы складываются, как показано на векторной диаграмме. Суммарный вектор обозначен жирной стрелкой
Причиной броуновского движения являются толчки, вызванные тепловым поступательным движением молекул. Отдельные импульсы складываются, как показано на векторной диаграмме. Суммарный вектор обозначен жирной стрелкой
Остановимся на явлении броуновского движения молекул. Очевидно, не только отдельные молекулы, но и подталкиваемые ими мельчайшие составные части клеток находятся в непрерывном движении. Теперь попытаемся исследовать роль этого движения в жизни клетки.
Обратимся к реальной жизни. Прекрасным воскресным утром яхтсмен поднимает парус, отталкивается от причала и, воспользовавшись свежим попутным ветром, отплывает в море. Сила и направление ветра в течение дня изменяются, тем не менее благодаря своей ловкости и морской удаче яхтсмен к вечеру снова возвращается к причалу. Как это ему удалось при совершенно случайно меняющемся ветре? Ведь мяч, брошенный в то же время на волю ветру и волнам, уже давно скрылся где-то вдали! Совсем не обязательно быть яхтсменом, чтобы ответить на этот вопрос. У яхты есть руль, киль и подвижный парус. Она может идти под различными углами к направлению ветра и даже против него. Но для этого уже требуется некоторое умение. Это умение, или, говоря научно, хранящаяся в голове яхтсмена информация, способно обуздать слепой произвол ветра; яхтсмен может направить лодку в любом направлении и даже против ветра, используя при этом его же силу.
Попробуем найти параллель этому в живой клетке. Клетки кишечника поглощают питательные вещества. Мелкие молекулы этих веществ без труда проникают сквозь клеточную мембрану, а крупные попадают внутрь за счет того, что мембрана выгибается и образует вокруг молекулы крохотный пузырек. Клетка как бы "пьет"; этот процесс называют пиноцитозом. Пузырьки путешествуют по клетке и, дойдя до ее противоположной стороны, исчезают — вещество поглотилось клеткой. Какая же сила толкает эти "продуктовые посылки"?
Сейчас не без оснований предполагают, что частицы путешествуют в клетке, подгоняемые "ветром броуновского движения молекул". Правда, это движение не так изящно и свободно, как скольжение парусника по водной глади. Вероятно, в клетке проложены своего рода "рельсы" — внутренние ограничивающие плоскости, которые препятствуют движению пузырьков в нежелательном направлении. Пока это не известно точно, но, по-видимому, в тех местах, где питательные вещества должны использоваться, существуют специальные рецепторы, захватывающие частицы. Если же пузырек "приплывает" в неподходящее место, он не задерживается здесь, а отправляется дальше. Такое бесцельное странствование продолжается до тех пор, пока пузырек наконец не попадет к нужному адресату — рецептору, который схватывает и перерабатывает содержимое пузырька. Это, конечно, не самый быстрый метод доставки, но он надежен и "недорог", ибо "оплата" производится за счет теплового движения.
 Так схематически можно представить механизм движения пиноцитозного пузырька по клетке. От места возникновения MB он странствует по клетке, участвуя в хаотическом броуновском движении, до тех пор пока не попадет к рецептору Р и не растворится
Так схематически можно представить механизм движения пиноцитозного пузырька по клетке. От места возникновения MB он странствует по клетке, участвуя в хаотическом броуновском движении, до тех пор пока не попадет к рецептору Р и не растворится
Но что же тогда означает "искусство плавания", "управляющая информация"? Это "адрес", по которому направлена "посылка", и умение распознать нужного адресата. Соответствующая информация заложена в пузырьке, и она представляет собой не что иное, как специфическую структуру белка, т. е. обусловлена опять-таки исходной наследственной информацией.
Можно было бы привести еще много примеров действия подобных механизмов в клетке. Но нам важен принцип. Итак, биологическая функция = энергия хаотического теплового движения + биологическая информация!
До сих пор не удается подсчитать и выразить эту информацию численно. Долгое время полагали, что "бит", взятый из теории информации, мог бы служить наиболее пригодной мерой информации. Однако это ожидание не оправдалось. Предлагаются различные пути решения этой проблемы, но каких-либо удовлетворительных результатов пока получить не удалось.
Таким образом, бесцельное поступательное движение молекул, хаос теплового движения, не обязательно вредит жизни, он включен в сам принцип функционирования живого и даже является необходимой предпосылкой для протекания большинства жизненно важных процессов.
Особенно показательна в этом отношении эволюционная теория Дарвина. Мы достаточно слышали о мутациях, обусловленных наследственными повреждениями. Мутации — это резкие скачкообразные изменения наследственного кода. К ним относятся замена, потеря или изменение основания в цепочке молекулы ДНК, возможно даже изменение целых хромосом. Как правило, большинство мутантов нежизнеспособны и не обнаруживаются в популяции в целом. Но многие мутации, проявляющиеся в нарушении строения тела или функции, очень заметны.
Причиной мутации, как правило, служат случайные события. И хотя при этом роль теплового движения не велика и изменение структуры молекул происходит в основном за счет действия квантов высокой энергии, например космического излучения, тем не менее и здесь в биологический порядок вклинивается случайность. Нельзя позволить этому явлению восторжествовать, ибо оно уничтожит любую биологическую организацию. Поэтому побеждают организмы, обладающие различными механизмами защиты, которые предохраняют организм от воздействия подобных случайностей. Однако со времен Дарвина известно, что без постоянной изменчивости и последующего отбора биологическая эволюция вплоть до человека была бы невозможна.
 Мутант: вторжение случайности микрофизического хаоса в генетическую упорядоченность жизни. Изменения, происходящие на молекулярном уровне, проявляются во внешнем виде растения. Дарвиновская теория эволюции была бы немыслима без мутаций
Мутант: вторжение случайности микрофизического хаоса в генетическую упорядоченность жизни. Изменения, происходящие на молекулярном уровне, проявляются во внешнем виде растения. Дарвиновская теория эволюции была бы немыслима без мутаций
Вернемся снова к тепловому движению молекул. Приведенный выше пример броуновского движения слишком специален и кажется притянутым за уши. Мы воспользовались им лишь потому, что он достаточно нагляден и, кроме того, показывает, что тепловое движение присуще не только молекулам, но и телам, видимым даже в обычный оптический микроскоп. Броуновское движение представляет собой хорошую модель любого молекулярного движения.
Вообще говоря, само движение молекул много важнее и интересней. Оно является причиной переноса вещества в самом широком смысле слова. Вещество может транспортироваться, возникать из другого вещества или превращаться в другое вещество.
Рассмотрим такой пример. Каждому известно, что в горячем чае сахар растворяется гораздо быстрее, чем в холодном. Любая химическая реакция при более высокой температуре протекает быстрее. Почему? Потому что любое преобразование вещества, любой процесс растворения является результатом столкновения молекул. Чем выше температура, тем чаще происходят столкновения, тем быстрее протекает реакция.
Остановимся на процессе растворения сахара в чае. Огромное множество молекул сахара, произвольно расположенных в куске, непрерывно атакуют молекулы воды. Непрерывно, ибо процесс растворения продолжается без остановки вплоть до последней молекулы сахара. Даже когда растворение идет медленно, каждую секунду чрезвычайно большое число молекул переходит из кристалла в раствор.
Что же это за числа, которые мы называем "чрезвычайно" большими? Известно, что один моль (грамм-молекула) вещества содержит 6⋅10
23 молекул — это 6 с 23 нулями. Чему равен моль? Молем принято обозначать количество граммов вещества, равное его молекулярной массе, а молекулярная масса — это, грубо говоря, число, показывающее, во сколько раз данная молекула тяжелее атома водорода. Если известна химическая формула вещества, то его молекулярную массу легко рассчитать с помощью таблиц. Молекула воды (Н
2О) в 18 раз тяжелее водорода, следовательно, 18 г воды составляют 1 моль и содержат 6⋅10
23 молекул. Такое же число молекул находится в 342 г сахара. Теперь мы получили представление о том, что подразумевается под "чрезвычайно" большим числом молекул.
Итак, огромное множество молекул воды постоянно атакует сахар и даже при самом медленном растворении колоссальное количество молекул сахара переходит в раствор. Что означает при таких больших числах увеличение или уменьшение на один порядок?
Сейчас на Земле живет порядка 10
9 человек. По сравнению с 10
23 молекул, содержащихся в одном моле, это мало. Однако, если рассчитать, сколько людей рождается и умирает за одну минуту во всем мире, то получится так много, что можно говорить о почти непрерывном рождении или смерти. Если взять большой промежуток времени, то эта величина может составить население большого города.
 Мельчайшие биологические структуры, отделенные от окружающей среды мембранами. Сколько молекул вещества они могут вместить? Можно ли здесь еще применять заимствованные из 'обычной' химии понятия, такие, как концентрация, вязкость, значения рН и т. д
Мельчайшие биологические структуры, отделенные от окружающей среды мембранами. Сколько молекул вещества они могут вместить? Можно ли здесь еще применять заимствованные из 'обычной' химии понятия, такие, как концентрация, вязкость, значения рН и т. д
Теперь попробуем сузить круг лиц, например до 30-40 человек, которые составляют круг наших знакомых; тогда рождение или смерть будет уже исключительным событием. Таким образом при переходе от большого числа к малому мы одновременно перешли от непрерывности к отдельному статистическому результату.
Так и в молекулярной биологии большое число 10
23 становится маленьким, если мало интересующее нас пространство. Встречаются структуры диаметром меньше 1 мкм. Вспомним митохондрию, маленькую клеточную органеллу, отделенную от цитоплазмы клетки двойной мембраной, — энергетическую станцию клетки с очень сложной системой ферментов, и остановимся на ней подробнее. Функционирование ферментов в митохондрии зависит от многих условий. Особую роль играет величина рН, характеризующая кислотность среды. Если внутренняя среда митохондрии нейтральна, количество ионов водорода в ней равно количеству гидроксильных ионов и составляет 10
7 грамм-ионов на 1 л, иначе 10
-7 моль/л. Как нам теперь известно, в 1 моле содержится 6⋅10
23 молекул (атомов или ионов), т. е. в 1 л нейтрального раствора присутствует 6⋅10
16 ионов Н
+.
Для простоты предположим, что объем митохондрии равен 1 мкм
3, или 10
-15 л. В таком случае в этой органелле плавает не больше 6⋅10
16⋅10
-15 = 6⋅10
1 = 60 ионов водорода. Итак, из очень большого числа получилось маленькое! Что такое 60 ионов, если от того, вступит ли хотя бы один из них случайно в химическую реакцию с молекулой фермента в нужный момент и в нужном месте, зависит биологический процесс? Разве может быть гарантирована непрерывность функционирования, "надежность" работы митохондрии? В такой высокоорганизованной системе, как биологическая, отводить большую роль случаю, конечно же, нельзя.
И здесь мы сталкиваемся с фундаментальной проблемой биологии. В первой главе нас интересовало, почему слон не может быть еще больше, а мышь еще меньше. Ответить на эти вопросы можно было с помощью макрофизики и физиологии. Теперь мы можем спросить, почему не могут быть еще мельче клетки, одноклеточные, бактерии?
По-видимому, этот вопрос действительно связан с борьбой за надежность функционирования системы. Если бы клетка была слишком маленькой, то концентрации молекул в ней или отдельных ее участках оказались бы настолько малыми, что вероятность случайного столкновения молекул, вступающих в химическую реакцию, стала бы слишком низкой и соответствующие химические реакции протекали бы очень медленно. Клетка должна быть достаточно велика, чтобы иметь возможность противостоять случайным флуктуациям теплового хаоса. В этом случае за промежуток времени, необходимый для осуществления какого-либо биологического процесса, с достаточной надежностью будет происходить нужное число случайных столкновений молекул.
Итак, в игру вступает время! Его роль мы уже отмечали на примере со статистикой населения. Можно с уверенностью утверждать, что именно в данную минуту, не позже чем секундная стрелка наших часов сделает полный оборот, в мире родится по крайней мере один ребенок. Однако такое утверждение несправедливо, если мы будем рассматривать только одну какую-либо страну. Если же промежуток времени увеличить до одного часа или дня, то и в последнем случае надежность нашего утверждения значительно повысится.
Вернемся к молекулярной биологии. Если вероятность удачного столкновения в маленьком объеме мала, надо выбрать больший интервал времени. Обобщая, можно сказать так: более мелкие клетки должны быть более медлительными. Таким образом, в мире молекул существует определенная взаимосвязь между пространством и временем.
Конечно, было бы любопытно поближе познакомиться с пространственно-временными соотношениями. Как мала и насколько быстро действует та или иная система? Что нам известно о реакциях, которые могут протекать в столь малых пространствах? Химики тщательно изучили реакции
in vitro, т. е. в пробирках. Нерешенные вопросы остаются и здесь, но многое известно уже достаточно хорошо. В растворе находится огромное множество молекул и ионов. Они равномерно перемешаны и хаотически движутся.
Другое дело — клетка. Она практически целиком состоит из перегородок и заключенных между ними мельчайших капиллярных пространств. Эти перегородки очень хорошо "продуманы". Они устроены примерно так же, как клеточные мембраны, о которых мы уже говорили. На большинстве перегородок располагаются белковые молекулы. Они несут заряды. Другие молекулы, находящиеся между ними, тоже электрически заряжены. Эти заряженные молекулы притягивают ионы, которые образуют более или менее плотный слой. Создается так называемый двойной слой положительных и отрицательных зарядов. Между электрическими зарядами возникает электрическое поле огромной силы; полагают, что его напряженность достигает 10 млн. В/см (вольт на сантиметр). Именно такая напряженность необходима для того, чтобы ионизировать молекулы, не разрывая их. Эти поля воздействуют на молекулы с большой силой. Они выравнивают заряженные молекулы и поляризуют незаряженные.
Мы почти ничего не знаем о биологическом действии этих полей, но можем предполагать, что они являются оружием жизни в борьбе с хаосом. Хранящаяся в ДНК информация определяет структуру белков и их расположение относительно друг друга, создает электрическое поле, сдерживающее хаос. Здесь царит порядок, который нельзя видеть даже с помощью электронного микроскопа, но который нам нужно как-то оценить. Чтобы рассчитать электрические поля даже для самых гладких металлических электродов, например сделанных из ртути, мы вынуждены прибегать к помощи ЭВМ. Что касается биологических поверхностей, структура которых в основном не известна, то пока здесь можно лишь высказывать догадки.
Конечно, в микромире для нас представляют загадку не только электрические поля. Многие физические понятия, такие, как вязкость, поверхностное натяжение, упругость и даже температура, имеют смысл только для больших систем. Но они теряют его, когда речь идет о малых и мельчайших системах, неоднородных, или функционирующих неравновесных, системах. Можно ли вообще говорить о температуре биологической системы? Этот вопрос кажется весьма странным, и тем не менее до сих пор он остается без ответа. Температура, как скажет физик, это свойство "канонической (однородной) совокупности" частиц, которая выравнивается благодаря взаимным соударениям между ними. В биологической системе такого выравнивания, вероятно, никогда не происходит, продукты реакции перебрасываются от одной молекулы к другой под влиянием внутренних электрических полей. У каждой частицы своя собственная "температура".
Однако это уже из области предположений. Подобная экскурсия в мир фантастики оправдана только в том случае, если она пробуждает в нас стремление к новым критическим исследованиям. Данный вопрос относится к разряду тех многих нерешенных проблем, которые стоят перед специалистами в области молекулярной биологии, но которые могут быть решены лишь в содружестве с учеными, занимающимися квантовой механикой и статистической физикой.
Вместо заключения
"Занавес опущен — действие продолжается". Является ли вообще биолог собственно биологом? О специалисте, который не сведущ ни в чем. Поток информации. Коллективное исследование как единственное спасение. Можно ли взнуздать лошадь с хвоста, или как стать биологом? Следует ли биологии опасаться своих соседей? Новая картина биологии. И все-таки она есть!
Что, собственно говоря, представляет собой биология? С этого вопроса мы начали разговор. Но ответ на него не был целью нашей книги. Напротив, возможно, что после ее чтения читателю гораздо большее покажется неясным, чем было прежде. Должен ли биолог быть одновременно химиком, специалистом в области кибернетики, квантовой физики, электроники, термодинамики и т. д.? Возможно ли это? Может быть, по этой причине биологов вообще больше не существует, а есть только физики, химики, математики, инженеры, занимающиеся вопросами живого?
Эти вопросы серьезны и важны. И простыми "да" или "нет" на них не ответишь. У нас осталось еще множество нерешенных проблем, которые мы попытаемся как-то упорядочить и обсудить последовательно.
Бесспорно, в наше время дальнейшее развитие биологии невозможно без помощи других естественных, а также математических и технических наук. Современный студент-биолог более половины своего учебного времени посвящает изучению небиологических, смежных дисциплин. Это необходимо ему независимо от того, в какой области биологии он будет работать в будущем. Бели он стремится попробовать свои силы на стыке наук, то он специализируется в соответствии с этим и становится биофизиком или биохимиком.
Наши знания в области естественных наук сейчас настолько велики, что надеяться на успех в основном могут только достаточно узкие специалисты. Теперь уже не встретишь физика или химика, который, хорошо разбираясь во всех тонкостях собственного предмета изучения, был бы в то же время знаком со всеми специальными проблемами и новейшими результатами исследований в других разделах науки. В равной степени это относится и к биологу — ученому, объектом исследования которого являются в высшей степени развитые и сложные живые структуры. Дробление специальных наук продолжается, область знаний, которую в состоянии охватить один человек, становится все уже.
Покажем это на примере. Согласно грубой оценке, ежедневно в мире выходит около десятка научных публикаций, посвященных новейшим исследованиям биологических мембран. Даже если объем каждой из статей пять страниц — а многих из них больше, — специалист, занимающийся изучением этой проблемы, должен ежедневно прочитывать и перерабатывать пятьдесят страниц научного текста. А это отнюдь не роман!Это описание, как правило, весьма лаконичное, опытных установок, вывод математических формул, анализ полученных кривых; кроме того, все это часто написано на иностранном языке. Если бы ученый взялся читать все это, он, конечно, знал бы, что делают другие исследователи в данной области, но у него не осталось бы ни минуты времени для собственной исследовательской работы.
Выход один: из десятка статей в день отбирается одна-две, наиболее тесно связанные с собственной узкой темой исследования. Биология мембран слишком обширна. Ее подразделяют на биохимию активного транспорта через мембрану, электрохимические проявления потенциала действия, транспорт Сахаров через мембрану клеток кишечника и т. д. Если ограничить себя узкими рамками, то можно успеть не только следить за текущей литературой, но и самому заниматься научными изысканиями.
Но не приводит ли такая специализация в конце концов к специалисту-невежде, человеку, который все более сужает область своих научных интересов, пока наконец не приходит к тому, что ничего не знает обо всем другом. Эта опасность сегодня велика!
Характерной особенностью современных исследований является сотрудничество ученых разных специальностей при решении одной общей проблемы. Специалисты по электронике разрабатывают и конструируют сложные измерительные приборы, биологи препарируют биологический объект, химики анализируют его состав, физики и математики обрабатывают полученные данные. И совместная работа всех этих специалистов строится довольно непросто.
Научные исследования — не производство предметов широкого потребления. Любой из предметов нашего обихода, прежде чем попасть к потребителю, проходит через руки многих специалистов. При этом рабочий не обязательно должен представлять себе вид готовой продукции. Например, рабочий, изготовляющий шарниры, не знает, будут ли их встраивать в шкаф, радиолу, футляр для прибора или пианино. Это его не интересует, ему вполне достаточно знать, что шарниры данного размера где-то используются.
В отличие от этого каждый член исследовательского коллектива должен представлять себе проблему в целом. Специалисту по электронике нужно знать электрические параметры изучаемого биологического объекта, чтобы эффективно и безошибочно применять измерительную технику. Физик должен иметь представление о множестве биологических явлений, которые могут повлиять на результаты измерений. Биолог в соответствии с требованиями своих коллег должен уметь выбрать наиболее подходящий объект для изучения. Но главное — всем нужно знать проблему, знать, что сделано в мире в этой области, как наиболее рационально подойти к решению задачи, чтобы достаточно быстро добиться эффективных результатов. Далее необходимо проанализировать результаты и. сформулировать в виде рабочих гипотез или теорий новые задачи. Итак, все заняты биологией! Значит ли это, что все, работающие над этой темой, — биологи?
Если называть биологом исследователя, который изучает живую систему, тогда действительно все сотрудники исследовательской группы — биологи независимо от полученного ими образования. Если же считать биологом ученого, усвоившего известные в настоящее время общие взаимосвязи и закономерности, присущие такому явлению, как жизнь, и не учитывать, в какой области он в настоящее время работает, то на поставленный выше вопрос следует ответить отрицательно.
Здесь мы подошли к следующей проблеме: действительно ли существует собственно биология со своим специфическим образом мышления, типичными закономерностями и т. д. или она разделяется на физику и химию живого? Снова приведем конкретный пример.
Допустим, биолог, слабо знакомый с математикой, узнаёт, что некоторые проблемы можно решить с помощью дифференциальных уравнений. Поскольку эти проблемы его интересуют, он хочет ознакомиться с данным методом ближе. Он покупает учебник по дифференциальным уравнениям и сразу же обнаруживает, что его знания в области дифференциального исчисления недостаточны, чтобы понять эту книгу. Он приобретает .новый учебник. На первых же страницах он сталкивается со значком "lim". Что это такое? Он обращается к исчислению бесконечно малых величин, пределам сходимости рядов. Снова учебники! Затем следуют функции, векторы — очень скоро человек оказывается окруженным горой учебников и приходит к выводу: помочь ему может только систематическое изучение математики в вузе с самого начала!
Подобную же картину мы увидим, если вместо математики обратимся к физике. Для понимания специальных физических проблем также необходим надежный фундамент, который дает только систематическое образование. Вот почему биологи так редко находят путь в физику и математику. У кого хватит времени и терпения начать систематическое обучение с самых азов?
Теперь рассмотрим обратную ситуацию. Физик, химик или математик заинтересовались какой-то специальной проблемой, скажем ростом бактерий. Здесь любому из этих специалистов придется легче, чем биологу, хотя в учебнике по микробиологии каждый из них также встретит множество незнакомых терминов. Об одних ему придется где-то справиться, другие он может просто пропустить. При некотором усердии через относительно короткое время он узнает о бактериях достаточно для того, чтобы применять свои математические, физические или любые другие специальные знания.
Стал ли он при этом биологом? Конечно, он может многое сделать, работая в содружестве с "классическим" биологом, ведь теперь он лучше разбирается в биологической проблематике. Его аналитический, натренированный в другой области знаний ум может оказаться здесь очень полезным. Но может случиться и так, что он поведет себя в биологии как слон в посудной лавке. Наивное невежество будет побуждать его строить самые невероятные гипотезы, которые не выдерживают биологической проверки или попросту представляют собой умствование, лишенное смысла. Подобных примеров великое множество. Нередко пытаются стрелять из пушек по воробьям, которые притом и сидят где-то совсем близко. И если математический аппарат достаточно сложен, биолог не в состоянии сразу раскусить подобный обман.
Биология в отличие от математики и физики не заходит за их границы, которые абсолютно неприступны для любого чужака. Но в биологии, так же как в физике и математике, творческая работа возможна только после ознакомления с основами этой науки. Микробиолог должен знать и клетки животных. В своих изысканиях он должен учитывать эволюцию, а следовательно, ему необходимы некоторые знания о формах. В современном представлении эволюция — это сравнение не только форм, но и функций. Как развивались те или иные ферментные системы? Где и как впервые возникли определенные механизмы биологических реакций?
Итак, биология существует! Внедрение в нее методов и идей смежных дисциплин не ведет к дроблению общей картины этой науки. Напротив, в результате сотрудничества с другими науками современная биология значительно обогатилась возможностями, перспективами и знаниями. Расширился и круг ученых, работающих в биологии. Приведенный нами пример ни в коем случае не должен отпугивать физика от биологии, принижать его успехи в этой области. Огромный вклад, который физики внесли в биологию, подчеркивает и список лауреатов Нобелевской премии за последнее десятилетие. Во многих лабораториях мира физики работают бок о бок с биологами, и многие из них стали биологами в истинном смысле этого слова. То же можно сказать о химиках, математиках, инженерах. Современная биология, как и любая другая наука, не боится богатства идей.
Наша книга, разумеется, не претендует на изложение всех представлений современной биологии. Это всего-навсего попытка взглянуть на биологию с иной точки зрения. Мы лишь мимоходом коснулись здесь различных проблем и суждений. Большинство рассмотренных нами примеров относятся к той области, которую в настоящее время определяют понятием "биофизика", хотя такое определение ограниченно и неточно. Мы хотели обратить внимание читателя на то, что развитие этого специального направления значительно обогатило биологию, что современную общую биологию невозможно представить без учета вклада, сделанного биофизикой. И если затронутые здесь проблемы пробудили у читателя интерес, автор считает свою задачу выполненной.
Примечания
1
Шредингер Э., Что такое жизнь? С точки зрения физика; М., Атомиздат, 1972.
(обратно)
2
Витализм (от латинского vitalis — жизненный, животворный, живой) — идеалистическое течение в биологии, согласно которому во всех живых организмах существует особая нематериальная жизненная сила. Витализм берет начало от первобытного представления об одушевленности всех тел природы. — Прим. ред.
(обратно)
3
Обширный отряд вымерших головоногих моллюсков; они обладали наружной раковиной, завитой в плоскую спираль с плотно налегающими оборотами. — Прим. ред.
(обратно)
4
Изотоническими называются растворы, имеющие одинаковое осмотическое давление. В данном случае имеется в виду, что внутри клетки осмотическое давление такое же, как во внешней среде. — Прим. рeд.
(обратно)
5
Отдельные компоненты динамических систем можно маркировать, т. е. метить, по-разному. Например, при анализе молекулярных систем используют радиоактивные изотопы, при анализе популяций микроорганизмов — генетические метки и т. д. — Прим. ред.
(обратно)
6
Автотрофам для их существования достаточно наличия воды, двуокиси углерода, неорганических солей и подходящего источника энергии. В отличие от автотрофов гетеротрофные организмы не способны синтезировать питательные вещества из неорганических соединений. Поэтому гетеротрофы (сюда относятся животные, грибы, многие бактерии) вынуждены жить за счет автотрофов. — Прим. ред.
(обратно)
7
Флуоресценция (от названия минерала "флюорит", у которого впервые была обнаружена флуоресценция, и латинского escent — суффикс, означающий слабое действие) — световое излучение, очень быстро затухающее после окончания возбуждения (время затухания около 10
-8 с). Флуоресценция возникает в результате самопроизвольных переходов возбужденных молекул (атомов) в нормальное состояние. По спектрам флуоресценции можно судить о свойствах молекул. — Прим. ред.
(обратно)
8
Здесь имеется в виду разность потенциалов между внутренней и внешней поверхностями мембраны. — Прим. ред.
(обратно)
9
Персонажи трагедии И. Гёте "Фауст" (пер. с нем. Н. Холодковского, часть первая, сцена 1). — Прим. ред.
(обратно)
Оглавление
Предисловие редактора перевода
Введение. Тем, кто хочет изучать биологию, хотя и не собирается стать директором зоопарка
Большое и малое в сравнении
Форма живых организмов в системе координат
Все течет, все изменяется
Беглый взгляд на механизм живого
В царстве молекул
Вместо заключения
*** Примечания ***