Дональд Протеро
Отпечатки жизни
25 шагов эволюции и вся история планеты

Посвящаю эту книгу нашим великим
популяризаторам и защитникам науки
Нилу Шубину
Биллу Наю
Нилу Деграссу Тайсону
и покойным Карлу Сагану и Стивену Джею Гулду
Предисловие
История жизни на Земле удивительно сложна. В настоящее время на нашей планете обитают от 5 до 15 миллионов видов живых существ. Поскольку свыше 99% когда-либо существовавших видов уже вымерли, можно сделать вывод, что на Земле успели пожить сотни миллионов биологических видов, если не больше, с тех пор как зародилась жизнь — а это произошло 3,5 миллиарда лет назад или даже раньше.
Поэтому выбрать всего 25 окаменелостей, чтобы на их примере представить сотни миллионов вымерших видов, — непростая задача. Я сосредоточился на окаменелостях, знаковых в истории эволюции. Они демонстрируют ключевые этапы зарождения крупных групп биологических видов либо иллюстрируют эволюционные переходы от одной группы к другой. Кроме того, жизнь — это не просто возникновение новых групп видов, а поразительное проявление разнообразных адаптаций, в ходе которых менялись размеры организмов, занимаемые ими экологические ниши и среда обитания. Поэтому я выбрал несколько «экстремальных» ископаемых форм, демонстрирующих, чего может достичь биология. Вы познакомитесь и с крупнейшим наземным животным, и с самым большим сухопутным хищником, и с некоторыми гигантскими морскими тварями, когда-то бороздившими океан.
Естественно, такой сложный подход вынуждает игнорировать многих существ, поэтому я мучительно выбирал, кого описать, а кого пропустить. Старался подобрать окаменелости, относительно хорошо сохранившиеся и известные. Таким образом, были исключены многие виды, останки которых слишком фрагментарны, чтобы уверенно их интерпретировать. Учитывая интересы широкого круга читателей, я отдавал предпочтение динозаврам и позвоночным. Прошу прощения у всех моих знакомых палеоботаников и микропалеонтологов — за то, что обделил их дисциплины, посвятив обеим всего по главе.
Надеюсь, вы простите мне мои грехи, деяния и недеяния и уделите внимание существам, истории о которых я хочу вам рассказать. Пусть они сделают вашу жизнь ярче!
Благодарности
Благодарю Патрика Фитцджеральда (Patrick Fitzgerald), Кэтрин Шелл (Katherine Schell) и Ирэн Пэвитт (Irene Pavitt) из издательства Columbia University Press — за их помощь в этом проекте. Моя особая признательность Патрику — за то, что предложил мне эту идею и сделал много ценных замечаний. Ещё благодарю Брюса Либермана (Bruce Lieberman) и Дэвида Арчибальда (David Archibald) — за то, что вычитали готовый черновик этой книги, а также Майка Эверхарта (Mike Everhart) и Тома Хольтца (Tom Holtz) — за рецензирование отдельных глав. Особенно признателен Даррену Нэйшу (Darren Naish) — за его энциклопедические знания о наземных позвоночных (тетраподах), что позволило ему тщательно проверить 15 последних глав. Людей, которые любезно предоставили фотографии и иллюстрации для этой книги, я благодарю в соответствующих местах. Спасибо Тамуре Нобумичи (Nobumichi Tamura), Карлу Бьюэллу (Carl Buell) и Мэри Персис Уильямс (Mary Persis Williams) за их изумительные рисунки, украшающие эту книгу.
Кроме того, благодарю моих сыновей Эрика, Закари и Габриэля за любовь и поддержку, которой они меня окружили, пока я писал книгу. Особая благодарность моей восхитительной жене, доктору Терезе Левель (Teresa LeVelle) за её поддержку и мотивацию, а также за то, что помогала мне выкраивать время, чтобы закончить книгу в срок.
01. Планета ила
Первые окаменелости
Криптозоон
Следовательно, если эта теория верна, то не может быть сомнения в том, что прежде, чем отложился самый нижний кембрийский слой, прошли продолжительные периоды... и мир изобиловал живыми существами... На вопрос, почему мы не находим богатых ископаемых отложении, относящихся к этим предполагаемым древнейшим периодам... я не могу дать удовлетворительного ответа.
Чарльз Дарвин. «Происхождение видов путём естественного отбора»
Дилемма Дарвина
Когда Чарльз Дарвин в 1859 году опубликовал свой труд «О происхождении видов», палеонтологическая летопись являлась слабым местом его аргументации. На тот момент не было известно практически ни одно ископаемое, которое можно было бы с уверенностью причислить к переходным звеньям. В частности, ни одной из окаменелостей, о которых пойдёт речь в этой книге. Первой подходящей окаменелостью такого рода стал
археоптерикс, которого предстояло открыть в 1861 году (см. главу 18). Ещё более тревожным представлялось отсутствие каких-либо окаменелостей, датируемых ранее кембрия — самого первого периода палеозойской эры, начавшегося около 550 млн лет назад (см. иллюстрацию на фронтисписе). Разумеется, в середине XIX века палеонтологическая летопись ещё была мало изучена: к тому моменту историю окаменелостей детально отслеживали всего около 60 лет. Кроме того, Дарвина озадачил тот факт, что в немногочисленных докембрийских отложениях, где были найдены древнейшие трилобиты, отсутствовали окаменелости, которые бы отражали переход от примитивных существ к трилобитам и другим организмам кембрия. Дарвин чётко отмечает это во фразе, вынесенной в эпиграф данной главы.
Учёный связывал загадочное отсутствие окаменелостей с «несовершенством геологического разреза» и маловероятной возможностью, что большинство организмов вообще не фоссилизируются. В сущности, он был прав. Дарвин обратился с этим вопросом к коллегам-учёным, и на протяжении следующих ста лет палеонтологи отчаянно пытались отыскать какие-либо окаменелости старше трилобитов.
Многим геологам уже известны проблемы, связанные с окаменелостями докембрийского периода. Большинство докембрийских слоёв настолько древние и находятся на таких глубинах, где под действием высоких температур и сильнейшего давления давным-давно превратились в метаморфические породы. При этом все окаменелости, которые могли в них сохраниться, разрушились. Большинство действительно древних пород подверглись эрозии — ещё одной разрушительной силе. Даже там, где степень сохранности относительно хорошая, древнейшие породы скрыты под наслоениями более молодых, поэтому почти везде на Земле они редко выходят на поверхность. Все названные факторы противоречат идее о том, что мы могли бы извлекать окаменелости из докембрийских слоёв с тем же успехом, что и из кембрийских.
Однако есть и другая проблема. Оказывается, в условиях докембрийской окружающей среды (когда почти не было кислорода и отсутствовал озоновый слой) древнейшие организмы, видимо, долго не могли отращивать раковины и другие жёсткие органы. Действительно, на протяжении 2 млрд лет мир был «населён» бактериальной биоплёнкой и (гораздо позже) водорослями, которые росли на мелководье у побережий и покрывали камни (рис. 1.1). В докембрийских породах
есть окаменелости, но большинство из них микроскопичны. Чтобы увидеть желаемое, надо тщательно перетирать тонкие каменные срезы, а потом рассматривать их с большим увеличением. Практикующий геолог просто не заметит окаменелости в большинстве докембрийских пород.
 Рис. 1.1.
Рис. 1.1. Реконструкция мелких приливных заводей, которые существовали на Земле 80% времени биологической эволюции, с 3,5 млрд до 550 млн лет назад. Единственными заметными останками жизни в них были бугорчатые отложения цианобактериальных сообществ (матов), так называемые строматолиты (рисунок Карла Бьюэлла из работы:
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 7.1)
Тем не менее в этих породах есть множество характерных черт, дискуссии о которых не утихают долгое время. Например, структуру, внешне напоминающую странный узор лучевидных бороздок, в 1848 году описал один из первых канадских геологов — сэр Джон Уильям Доусон, назвавший свою находку
олдхамия (рис. 1.2). Он полагал, что обнаружил ископаемые останки полипа. Но ирландский геолог Джон Джоли, однажды пройдя по оставленной в грязи замёрзшей колее, обнаружил подобный узор, образованный кристалликами льда. В 1884 году он заявил, что олдхамия — просто характерный отпечаток ледяных кристаллов, а не окаменелость. Сравнительно недавно учёные вновь исследовали олдхамию, и теперь считается, что такие бороздки мог оставить какой-то червь, то есть, в конечном итоге, это след живого организма. Однако приведённый пример демонстрирует, как легко люди обманываются, когда изо всех сил пытаются отыскать признаки докембрийской жизни. Другую «тварь» в 1868 году открыл легендарный биолог и защитник Дарвина Томас Генри Гексли, заметивший слизистый «организм» в пробах ила, поднятых с морского дна. Родовое название можно перевести с греческого как «глубокая жизнь», а видовое дали в честь немецкого биолога Эрнста Геккеля. Однако знаменитый британский естествоиспытатель Чарльз Уайвил Томсон не впечатлился. Он изучил образцы и пришёл к выводу, что это лишь продукт разложения неких грибков. Другой биолог, Джордж Чарльз Уоллич, предположил, что «организм» — продукт химического распада органических веществ.
 Рис. 1.2.
Рис. 1.2. Первое изображение олдхамии (перерисовал Э. Протеро)
По этой и иным причинам Уайвил Томсон и многие другие британские исследователи организовали и профинансировали экспедицию «Челленджера», длившуюся с 1872 по 1876 год. «Челленджер» — основательно снаряжённый парусник, оснащённый паровым двигателем, стал первым кораблём, совершившим кругосветное океанографическое путешествие. На тот момент британское научное сообщество понятия не имело, что собой представляет дно океана; предполагалось, что на глубине по-прежнему могут обитать трилобиты. Кроме того, учёные хотели понять, чем на самом деле является батибия Геккеля. «Челленджер» взял более 360 проб ила с океанского дна, но не обнаружил ни одной батибии. Тогда корабельный химик Джон Янг Бьюкенен изучил более старые образцы и нашёл нечто, напоминавшее таинственную «слизь». Проанализировав вещество, он обнаружил, что это лишь продукт реакции между сульфатом кальция и спиртом, в котором законсервировали образец. Уайвил Томсон отправил Гексли вежливое письмо, в котором проинформировал того о результатах «идентификации» организма, проведённой Бьюкененом. Следует отдать Гексли должное: он опубликовал в журнале Nature письмо, где признал свою ошибку. В 1879 году на регулярном собрании Британской ассоциации по содействию научному прогрессу Гексли взял на себя полную ответственность за допущенную ошибку.
Ещё одна ложная тревога случилась в 1858 году, за год до публикации дарвиновского «Происхождения видов». Легендарный канадский геолог Уильям Э. Логан, позднее возглавивший Геологическую службу Канады, обнаружил необычные минералы на берегах реки Оттава близ Монреаля. Логан в течение многих лет показывал образцы учёным, но большинство не были склонны видеть в них доказательство древней жизни. Затем эти образцы попали к Доусону, известному канадскому учёному. В 1865 году Доусон назвал слоистую структуру Логана
Eozoon Canadense (буквальный перевод — канадская заря жизни) (рис. 1.3). Доусон считал, что перед ним — фоссилизированные останки гигантской фораминиферы (одноклеточные существа, напоминающие амёб, живущие в океанах и образующие известковые раковины). Он назвал находку «одним из самых блистательных самоцветов в короне Геологической службы Канады». Однако вскоре после этого объявления другие специалисты внимательнее присмотрелись к геологическому строению образцов. Выяснилось, что эозоон — просто метаморфическое наслоение минералов кальцита и серпентина, а не окаменелость. Последний гвоздь в гроб «зари жизни» забили в 1894 году, когда вблизи Везувия в Италии было открыто, что подобные структуры могут образовываться в породах под действием вулканического тепла.

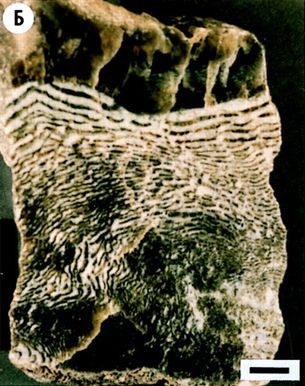 Рис. 1.3.
Рис. 1.3. Eozoon canadense (канадская заря жизни): (
А) иллюстрация из книги Доусона «The Dawn of Life»; (
Б) типовой экземпляр из Смитсоновского института. Масштабные полоски = 1 см ((
А) иллюстрация из книги
J. W. Dawson. The Dawn of Life. — London: Hodder and Stoughton, 1875, предоставлено J. W. Schopf)
Криптозоон: ещё одна ложная тревога?
Олдхамия, батибия, эозоон — лишь некоторые из псевдоокаменелостей, мнимых образцов докембрийской жизни, с которыми когда-то носились как с первопредками всех живых существ, а потом развенчали миф. Сегодня их помнят только специалисты по истории геологии.
Оглядываясь назад, легко понять возникшие заблуждения. Большинство геологов на ранних этапах своей карьеры усваивают, что геологический ландшафт изобилует псевдоокаменелостями и объектами, которые кажутся таковыми, пока к ним не присмотришься (зная, к чему присматриваться). Почти каждый геолог-любитель купится на «растительный узор» дендритов пиролюзита — минеральную форму оксида марганца, которая выглядит точь-в-точь как веточки папоротника. Большинство псевдоокаменелостей — это минеральные включения, представляющие собой крупицы песка или ила, цементирующиеся в виде самых разнообразных форм. Их большая часть напоминает сферы или пузырьки, но встречаются и более причудливые формы, которые неопытный дилетант вполне может принять за «окаменевший мозг» или «окаменевший фаллос». Многие другие очертания также заставляют нас обманываться и видеть «узоры» там, где их нет.
Мы видим «замки» среди облаков и «животных» на звёздном небе. Аналогичным образом человек жёстко запрограммирован выискивать смысл и закономерности практически в любом наборе случайных образов. Этот феномен называется парейдолия. Соответственно, опытные геологи учатся скептически относиться к любым попыткам трактовать камни причудливой формы как окаменелости. Требуются годы, чтобы научиться отличать первые от вторых. Это было особенно актуально на ранних этапах развития геологии, когда большинство осадочных структур и форм, образующихся при раскопках, ещё не были изучены и их с трудом отличали от истинно биологических окаменелостей.
Следующей важной фигурой в данной истории был Чарльз Дулиттл Уолкотт — геолог-самоучка, работавший в Геологической службе США (рис. 1.4).
 Рис. 1.4.
Рис. 1.4. Чарльз Дулиттл Уолкотт в сланцевом карьере Бёрджесс в 1912 году (фотография предоставлена Смитсоновским институтом)
Он получил только десятилетнее школьное образование, не имел учёной степени, но позднее был отмечен многими почётными академическими титулами. Именно Уолкотту предстояло стать одним из крупнейших американских учёных начала XX века. Практически в одиночку он документировал всю историю американского кембрия от штата Нью-Йорк до Большого каньона, также выступив в качестве основателя науки о докембрийских окаменелостях. Затем он прославился умением решать множество задач одновременно, масштаб которого мы сегодня едва ли можем себе представить. Уолкотт был директором Геологической службы США (1894–1907), потом его выдвинули на должность секретаря (директора) Смитсоновского института (1907–1927), причём он совмещал эту работу с должностью президента Национальной академии наук (1917–1923). Также он занимал президентские посты в Американском Философском обществе (как и Доусон) и в Американской ассоциации по содействию научному прогрессу. Несмотря на невероятную управленческую занятость, Уолкотт умудрялся каждое лето выкроить несколько недель на изнурительные полевые исследования, которыми занимался в Скалистых горах и на плато Колорадо. Там он описывал огромные горы, состоящие из кембрийских пород, и собирал обширные коллекции окаменелостей, на описание которых (с последующей публикацией работ) тоже находил время. Именно в ходе одной из таких экспедиций Уолкотт наткнулся на сланцы Бёрджесс — настоящее Эльдорадо среднекембрийских мягкотелых ископаемых (см. главу 5). Он бегло описал эти окаменелости, но, учитывая огромную занятость, не имел времени подробно их исследовать.
Свою научную карьеру Уолкотт начал, работая с легендарным Джеймсом Холлом — первым главным геологом и палеонтологом штата Нью-Йорк. Проводя отпуск в Саратоге, Уолкотт предпринял краткую вылазку в Лестер-Парк, что в пяти километрах к западу от Саратога-Спрингс. Там его впечатлила слоистая структура древнейших докембрийских пород, которые он изучал (рис. 1.5). В 1878 году, в возрасте всего 28 лет, он начал подробно описывать эти слоистые куполовидные структуры, по форме также напоминавшие капусту, которые Холл в 1883 году назвал криптозооном (скрытой жизнью). Они были типичны практически для всех докембрийских пород, поэтому Уолкотт не сомневался, что перед ним — древнейшие из сохранившихся биологических ископаемых.
 Рис. 1.5.
Рис. 1.5. Строматолиты из Лестер-Парк, которые Джеймс Холл и Чарльз Дулиттл Уолкотт назвали криптозооном (фотография автора)
Однако большинство других учёных отнеслись к идее скептически. Слоистые структуры легко образуются естественным путём, без участия живых организмов. Таковы, например, наслоения в метаморфических породах, которые Доусон ошибочно принял за ископаемую «зарю жизни», либо структуры, образующиеся при медленной кристаллизации из раствора или в процессе метаморфического разлистования. Авторитетный критик криптозоона, знаменитый учёный, Альберт Чарльз Сьюард, многие годы считавшийся самым влиятельным в мире специалистом по палеоботанике, справедливо указывал на то, что ранее не удалось найти сохранившиеся органические останки растений и других форм жизни. Поэтому случай криптозоона не вызывал доверия.
Тем не менее многие геологи продолжали описывать эти слоистые структуры, напоминавшие купола или капусту, представлявшие собой единственные характерные детали докембрийских пород, видимые невооружённым глазом, давали им названия. Кроме
Cryptozoon, появился род
Collenia, соответствовавший слоистой структуре чуть иной формы, а название
Conophyton присвоили слоистым конусообразным структурам. Советские учёные, которые могли исследовать огромные массивы неметаморфических докембрийских пород в Сибири, с особым удовольствием подбирали названия для всевозможных форм слоистых образований. Их объединили в более широкую категорию под названием
строматолиты (слоистые камни), хотя большинство геологов сомневались в биологическом происхождении данных образований.
Эврика!
В течение первой половины XX века в сообществе геологов и палеонтологов существовал глубокий раскол по поводу того, что представляют собой строматолиты. Исследования, проводившиеся одно за другим, не выявляли никаких признаков органики или сохранившихся клеток в их слоях, поэтому дело казалось сомнительным. Поскольку ни один из имевшихся экземпляров этих структур не рос и не подавал признаков жизни, отсутствовали серьёзные доводы, которые можно было бы предъявить скептикам.
В 1956 году геолог Брайан У. Логан из Университета Западной Австралии в Перте вместе с коллегами исследовал северо-западное побережье Австралии. Логан и компания добрались до залива Шарк, также известного под названием Акулья бухта, расположенного примерно в 800 км севернее Перта. Во время отлива в районе Хэмлин-Пул на южном берегу залива им открылся пейзаж возрастом в полмиллиарда лет, которого ещё не видел ни один учёный на Земле (рис. 1.6)! Дно бухты было усыпано цилиндрическими «башенками» с округлыми вершинами, каждая высотой от одного до двух метров. Они сильно походили на многие криптозооны и другие докембрийские строматолиты — но были живыми и росли! Более тщательный анализ показал, что эти столбы сложены из тончайших осадочных пород, накапливавшихся миллиметр за миллиметром, как древние строматолиты. На поверхности строматолитов обитали организмы, в результате жизнедеятельности которых и формировались таинственные структуры: вязкие скопления сине-зелёных бактерий, или цианобактерий (ошибочно названных «сине-зелёными водорослями», хотя таковыми не являвшихся — водоросли представляют собой настоящие растения с ядерными эукариотическими клетками). Сине-зелёные бактерии являются не только одной из самых простых, примитивных форм жизни на Земле, но, возможно, и первыми фотосинтезирующими организмами на нашей планете. Большинство учёных полагают, что сине-зелёные бактерии синтезировали первый атмосферный кислород на Земле, благодаря которому впоследствии развились более сложно организованные животные.
 Рис. 1.6.
Рис. 1.6. Цилиндрические строматолиты в заливе Шарк (Австралия) (фотография предоставлена R. N. Ginsburg)
Дальнейшие исследования строматолитов в заливе Шарк позволили выяснить, как образуется их тонкослойная структура. Эти слизистые маты стремительно растут на солнце в светлое время суток, когда накатывает прилив и они оказываются под водой. У свежих матов вязкая поверхность, на которой оседает различный материал — особенно ночью, во время отлива; рост цианобактерий приостанавливается на несколько часов. Затем, с новым приливом и восходом солнца бактерии опять тянутся к свету и отращивают новые нити, полностью накрывая осадок, который скопился за минувшую ночь. Так продолжается день за днём и год за годом. В благоприятной среде растущие маты захватывают сотни отдельных слоёв накапливающегося осадка. В конце концов органические останки бактерий разлагаются, и остаётся лишь слоистый осадок, не содержащий ни органических структур, ни химических следов их существования.
Итак, если этот процесс настолько прост, почему строматолиты не встречаются по всей планете, как в докембрии? Ответ подсказывает залив Шарк. Мелководье Хэмлин-Пула исключительно солёное, так как песчаная отмель, окружающая устье залива, ограничивает приток и отток воды. Кроме того, поскольку залив расположен в зоне субтропических пустынь, здесь очень жарко и солнечно. По мере испарения воды солёность осадка в мелкой бухте лишь возрастает. На самом деле, солёность здесь вдвое выше, чем в прилегающей части океана (более 7% против 3,5%); такие условия могут переносить только цианобактерии. Растительноядные моллюски (например, блюдечки, литорины и морские ушки, обитающие на современных приливных мелководьях), которые могли бы поедать подобные бактериальные маты, не могут жить в такой солёной воде, поэтому нетронутые маты продолжают расти. Эта экосистема очень напоминает докембрийский мир, когда более высокоразвитые растительноядные морские животные (например, улитки) ещё не существовали. Три миллиарда лет назад самой сложной формой жизни были бактериальные маты, и ничто не мешало их росту. Как отмечает мой друг Дж. Уильям Шопф из Калифорнийского университета Лос-Анджелеса, юная Земля была «планетой ила» (см. рис. 1.1).
С тех пор как в 1956 году было сделано открытие в заливе Шарк (первые публикации о нём появились в 1961 году), живые строматолиты удалось найти во многих местах на Земле. У большинства из них есть ключевая общая черта: они растут в среде, неблагоприятной для более развитых организмов (например, для растительноядных моллюсков). Я вплотную рассматривал строматолиты, растущие в солёных лагунах вдоль тихоокеанского побережья Нижней Калифорнии. Они обитают в солёных водах у западной оконечности Персидского залива. Огромные столбы с куполообразными вершинами, как в заливе Шарк, можно встретить и в солёных лагунах Лагоа-Салгада в Бразилии (название этого района так и переводится с португальского — солёные лагуны). Среди немногих строматолитов, выживающих в воде с нормальной солёностью, можно назвать образцы с отмелей Эксумы на Багамских островах: там такие сильные течения, что даже блюдечки и морские ушки не могут закрепиться.
Учёные постоянно находят новые ископаемые строматолиты, некоторые — столь же древние, как сама жизнь. К ним также относятся вероятные строматолиты из свиты Варравуна в Западной Австралии (всего на несколько сотен километров восточнее залива Шарк) — им 3,5 млрд лет, а также древнейшие микроскопические следы клеток цианобактерий. Настоящие строматолиты возрастом 3–4 млрд лет встречаются в группе Фигового Дерева (Fig Tree group) в Южной Африке. Примерно 1,25 млрд лет назад строматолиты достигли наибольшего разнообразия по форме, размеру и области распространения. Они до сих пор остаются единственным видимым свидетельством того, что уже в те времена наша планета была обитаема. Затем начался медленный упадок, а в кембрии их количество сократилось на 80% от исходного. Вероятно, виной тому были многочисленные растительноядные существа, истреблявшие строматолиты повсюду, где они росли в обычной морской воде. Строматолиты Лестер-Парк, показанные на рис. 1.5, относятся к хойтским известнякам — среднекембрийской формации. Это одни из немногих строматолитов, доживших до кембрия. К моменту массового распространения беспозвоночных в ордовике (около 500 млн лет назад) строматолиты на Земле почти исчезли.
При том, насколько они стали редки в последние 500 млн лет, микробные маты всегда готовы возродиться и расцвести махровым цветом, когда настанут тяжёлые времена для их пожирателей. После трёх великих вымираний, случившихся за историю Земли (в конце ордовика, в позднем девоне и самое массовое вымирание в конце перми), строматолиты вновь распространялись в «постапокалиптическом» мире, когда в результате вымирания биоразнообразие животных сильно сокращалось. В каждом случае строматолиты росли как сорняки, заселяя просторные ландшафты вместе с немногими уцелевшими оппортунистическими видами; их расцвет наступал всякий раз, когда вымирали питавшиеся ими животные.
Наконец, стоит учитывать ещё один момент. На протяжении почти 85% истории земной жизни (от 3,5 млрд примерно до 630 млн лет назад) на нашей планете не существовало столь крупных существ, от которых могли остаться заметные окаменелости. Только строматолиты можно рассматривать без микроскопа. Существует масса предположений, почему жизнь не развилась раньше. Многие из них сводятся к тому, что уровень атмосферного кислорода долгое время был недостаточным для существования многоклеточных организмов; ситуация изменилась только в кембрии. Каковы бы ни были причины, на протяжении большей части биологической истории на поверхности Земли существовали только бактериальные маты и куполообразные строматолиты — больше ничего. Если инопланетяне когда-либо высаживались на нашей планете, они могли увидеть это и, разочаровавшись, улететь.
Интересно вспомнить о метеорите ALH 84001, найденном в горах Алан Хиллс в Антарктиде. Сначала он откололся от Марса и лишь много позже оказался на Земле. В 1990-е годы развернулись нешуточные споры о крошечных шарообразных и продолговатых структурах в этом метеорите: не являются ли они окаменелостями марсианских организмов? Вопрос до сих пор остаётся открытым. Однако если на Марсе и
была жизнь, сейчас она почти наверняка замёрзла, поскольку на этой планете слишком холодно для существования жидкой воды.
[1] Земля должна была выглядеть почти так же: вплоть до 600 млн лет назад на ней существовали только одноклеточные организмы. Поэтому любой кусок земных пород и образец с земной поверхности выглядели бы в точности как марсианские породы до замерзания.
Лучше один раз увидеть!
Первые строматолиты, послужившие основой для криптозоона Джеймса Холла и Чарльза Дулиттла Уолкотта, можно увидеть в Лестер-Парке восточнее Саратога-Спрингс, в штате Нью-Йорк. Выезжаете из центра Саратога-Спрингс и отправляетесь на запад по шоссе 9N штата Нью-Йорк. Поверните налево на Миддл-Гроув Роуд, потом снова налево на Лестер-Парк Роуд (также известную под названием «Дорога окаменевших садов»). Проедьте по ней ещё около 150 м. Оказавшись в парке, следуйте по указателям, пока не прибудете в Окаменевшие Сады.
В некоторых музеях строматолиты экспонируются, в других можно встретить диорамы с их докембрийским изображением. Среди последних: Денверский музей природы и науки, Музей естественной истории под открытым небом (Чикаго), Геологический музей висконсинского университета в Мэдисоне, Смитсоновский институт (Вашингтон, округ Колумбия), Музей естественной истории штата Юта, город Солт-Лейк-Сити, Палеонтологический музей Реймонда Альфа в Уэбб-Скулз (Клермонт, Калифорния), Музей естественной истории штата Виргиния, Мартинсвилл и Музей Западной Австралии (Перт).
Дополнительная литература
Grotzinger, John P., and Andrew Н. Knoll. Stromatolites in Precambrian Carbonates: Evolutionary Mileposts or Environmental Dipsticks? //
Annual Review of Earth and Planetary Sciences. — 1999. — Vol. 27. — 313–358.
Knoll, Andrew H. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. — Princeton, N.J.: Princeton University Press, 2003.
Schopf, J. William. Cradle of Life: The Discovery of Earth’s Earliest Fossils. — Princeton, N. J.: Princeton University Press, 1999.
02. Эдиакарский сад
Первые многоклеточные организмы
Чарния
Начинающих палеонтологов обычно привлекают крупные эффектные экземпляры — например, плотоядные динозавры или плейстоценовые млекопитающие. Но чтобы отыскать настоящих монстров, диковинных обитателей утраченных миров, стоит обратиться к палеонтологии беспозвоночных. Бесспорно, самые странные из ископаемых организмов относятся к эдиакарской эпохе.
Марк Макменамин. «The Garden of Ediacara»
От одной клетки ко многим
Как мы узнали из первой главы, отсутствие каких-либо докембрийских окаменелостей длительный период времени представляло проблему для эволюционной биологии. Над ней ломал голову Чарльз Дарвин, и не только он. Так продолжалось до 1954 года, когда были открыты первые бесспорные микроокаменелости, и до того, как в конце 1950-х объяснили происхождение строматолитов, оказавшихся продуктом жизнедеятельности бактериальных сообществ. Эти открытия показали, что жизнь оставалась одноклеточной с 3,5 млрд примерно до 600 млн лет назад. Окаменелости многоклеточных организмов не встречаются вплоть до «кембрийского взрыва». Многие полагали, что в этой загадочной и таинственной эволюционной пропасти, закончившейся с появлением многоклеточных, которые обладали твёрдыми раковинами (например, трилобитов), обнаружить жизнь так и не удастся.
Но затем в породах стали находить любопытные окаменелости. Большинство из них остались от довольно крупных (до метра в поперечнике) мягкотелых существ, не развивших экзоскелета. Эти существа фоссилизировались в виде отпечатков на песчанике или аргиллитах морского дня, то есть среди них не было целиком сохранившихся тел (данная проблема возникает, если существо не обладает раковиной или экзоскелетом). Некоторые подобные образцы были найдены в Намибии в 1930-х и на Эдиакарских холмах в Австралии в 1940-х годах. Но они ещё не были датированы, поэтому предполагалось, что найденные окаменелости относятся к раннему кембрию.
 Рис. 2.1.
Рис. 2.1. Реконструкция чарнии (иллюстрация предоставлена Nobumichi Tamura)
Наконец, в 1956 году пятнадцатилетняя школьница Тина Негус нашла образец в лесу Чарнвуд близ города Грантем, в английском графстве Линкольншир (рис. 2.1). Вот как она его описывала:
Когда я была подростком, мне на глаза попалась монография о геологии чарнвудского леса, которую я нашла в местной библиотеке. Мы часто ездили в Чарнвуд, многие места, упоминаемые в книге, были мне знакомы. Я по максимуму скопировала карты из книги и упрашивала моих долготерпеливых родителей как можно скорее туда наведаться. Мы приехали и пробрались к карьеру. Я знала из книги, что здешние отложения состоят из напластованного вулканического пепла, долго лежавшего под водой, — раньше я о подобном не слышала. В то время карьер был заброшенным, тропинка напоминала овечью. Оказавшись на дне, я стала царапать склон и прямо на уровне головы нашла... окаменелость! Я ничуть не сомневалась, что это окаменелость, удивилась лишь тому, что во всех книгах, которые я читала, докембрий назывался эпохой, в которую ещё не возникла жизнь. Я подумала, что это папоротник, вернее, его веточка, но потом заметила, что у «листьев» нет центрального черенка, а сама чересполосица этих «листьев» постепенно переходит в «стебель».
На следующий день, придя в школу, я обратилась к учительнице географии, ведь я думала, что география — самая близкая к геологии наука. Сказала ей, что нашла окаменелость в докембрийских отложениях в Чарнвудском лесу. Она ответила: «В докембрийских породах нет никаких окаменелостей!». Я сказала, что знаю, но именно поэтому моя находка меня так озадачила. Она даже не сбавила шага, не взглянула на меня, просто сказала: «Значит, это НЕ докембрийские породы». Я уверяла её, что именно докембрийские, на что она повторила, что в докембрийских отложениях нет окаменелостей. Настоящий порочный круг и узколобость. Я отступилась на этот раз, но спросила родителей: а могли бы мы ещё раз съездить туда?
У Негус не было ни нужных инструментов, ни опыта, чтобы извлечь образец из твёрдой породы. Но годом позже это удалось другому местному школьнику по имени Роджер Мейсон, который потом стал профессором геологии. Парень отдал образец Тревору Форду — местному геологу, который в 1958 году официально опубликовал информацию о находке в «Трудах Йоркширского геологического общества». Форд назвал ископаемое
Charnia masoni (родовое название дано в честь Чарнвудского леса, видовое — в честь Роджера Мейсона) и предположил, что эта структура осталась от каких-то водорослей. Позднее геологи утверждали, что экземпляр относился к кораллоподобным организмам, так называемым «морским перьям», которые действительно напоминают мягкие подводные перья. Однако центральный «стебель» у
Charnia не прямой, как у папоротника, морского или обычного пера, а зигзагообразный. Как мы убедимся ниже, до сих пор не ясно, что это был за организм. Тем не менее это первая многоклеточная окаменелость (фактически
вообще первая окаменелость), найденная в докембрийских породах. Вспоминая учительницу географии из школы Негус, можно заключить, что до конца 1950-х годов выражение «докембрийская окаменелость» казалось нонсенсом. Все были уверены, что не сохранилось никаких докембрийских окаменелостей, видимых невооружённым глазом, поэтому образец был кембрийским либо просто не являлся окаменелостью.
Окаменелости хребта Флиндерс
Ещё до того, как была формально описана чарния, геологи открывали окаменелости крупных мягкотелых организмов в других частях света. Но поскольку находки делали в пластах неопределённого возраста, их обычно относили к кембрию. Ещё в 1868 году шотландский геолог Александр Мюррей нашёл в глубоководных морских песчаниках в районе Мистейкн-Пойнт у канадского острова Ньюфаундленд «ветвистые» окаменелости, напоминающие чарнию. Но тогда никто не знал, как их датировать или интерпретировать, поэтому о них просто забыли. В 1933 году немецкий геолог Георг Гюрих, занимаясь разведочными работами для золотодобычи в Намибии (в те годы данная территория являлась западной колонией ЮАР), составил геологическую карту страны. Там он нашёл многочисленные окаменелости интересных мягкотелых существ. Однако и на сей раз никто не мог определить возраст образцов, и их снова сочли кембрийскими.
Самые многочисленные и наиболее изученные образцы этой странной фауны были добыты на Эдиакарских холмах, относящихся к хребту Флиндерс в Южной Австралии, примерно в 336 км к северу от Аделаиды. В 1946 году австралийский геолог Реджинальд Спригг работал на Эдиакарских холмах, составлял геологические карты и оценивал заброшенные прииски, чтобы определить целесообразность возобновления разработок с применением новых технологий. Однажды, решив отдохнуть и пообедать, он заприметил первую из удивительных окаменелостей. Но он не был палеонтологом и сбор окаменелостей не входил в его обязанности, поэтому учёный просто рассказал о своих находках палеонтологу Мартину Глесснеру из Университета Аделаиды.
Глесснер был незаурядным человеком. Он родился 25 декабря 1906 года в северо-западной Богемии (ныне территория Чехии), когда она ещё входила в состав Австро-Венгрии. Получил образование в Венском университете, где к 25 годам стал как дипломированным юристом, так и доктором геологии. В начале научной карьеры Глесснера отправили в Москву, где он организовал микропалеонтологические исследования для Института нефти АН СССР. Соответственно он был одним из первых, кто использовал микроокаменелости для датировки нефтеносных пород и определения глубины древних вод. В Москве Глесснер познакомился с русской балериной Тиной Тупикиной и женился на ней. В данной ситуации ему оставалось принять советское гражданство либо покинуть СССР. После возвращения в Австрию в 1937 году Глесснер был вынужден почти сразу бежать из страны, поскольку страну оккупировали гитлеровцы (по отцу он имел еврейские корни). В итоге Глесснер с женой оказались в Порт-Морсби на Новой Гвинее, где учёному предложили организовать микропалеонтологическую службу для новообразованной Австралийской нефтяной компании. Затем война дошла до Новой Гвинеи, и в 1942 году Глесснер с женой бежали в Австралию, где учёный продолжал работать в нефтяной промышленности до 1950 года. Вплоть до завершения карьеры он занимал пост профессора и декана факультета геологии и палеонтологии в Университете Аделаиды. Именно там Глесснер занялся изучением таинственных окаменелостей, которые ему прислал Спригг, организовав масштабную работу по сбору множества новых экземпляров.
Проделав титаническую работу, он описал окаменелости, которые, на его взгляд, напоминали медуз, морские перья и разнообразных странных «червей» (рис. 2.2). Благодаря открытию чарнии в Англии и Австралии Глесснер смог продемонстрировать, что эдиакарские окаменелости относятся к позднейшему докембрию. Это доказывало, что удивительные крупные мягкотелые организмы отличались большим разнообразием и встречались по всему миру (в частности, в Африке, Австралии, Англии, на Ньюфаундленде и в России близ Белого моря). В 1984 году учёный подвёл итоги своей работы в книге «The Dawn of Animal Life» («Заря животных»), которая до сих пор считается классической. На поздних этапах карьеры Глесснер стал обладателем многих премий за первопроходческие работы по изучению древнейшей многоклеточной жизни.
Глесснер приложил массу усилий, чтобы интерпретировать эти любопытные отпечатки и следы на песчанике Флиндерс в сравнении с современными организмами (рис. 2.3). Круглые пузыри напоминали медуз, ветвистые формы — морские перья. Некоторые были исключительно крупными для древнейшей многоклеточной жизни. Например, широкие листовидные «черви» с перистым узором бороздок (так называемая дикинсония) могли достигать 1,5 м в длину (см. рис. 2.2)!


 Рис. 2.2.
Рис. 2.2. Эдиакарские окаменелости включают следы крупных мягкотелых волнистых организмов, известных только по отпечаткам на океаническом ложе. (
А) крупный ребристый «червь»
Dickinsonia; (
Б) членистый «червь»
Spriggina; (
В) щитовидный организм
Parvancorina, возможно, родственный трилобитам (фотографии предоставлены Смитсоновским институтом)
 Рис. 2.3.
Рис. 2.3. Диорама эдиакарской фауны, на которой реконструированы морские перья, медузы и «черви» (иллюстрация предоставлена Смитсоновским институтом)
Факт исключительной сохранности останков этих существ, которые должны были бы легко разлагаться, позволяет сделать ряд выводов: в позднем докембрии почти не было падальщиков; эдиакарские существа могли быть накрыты матами цианобактерий, способствовавшими их сохранности, либо многие из этих существ (особенно в районе Мистейкн-Пойнт, Ньюфаундленд) оказались похоронены заживо во время оползней ила на прибрежных мелководьях.
Камо грядеши, Эдиакара?
В дальнейшем учёные уже не были так уверены, что эдиакарские окаменелости легко причислить к ныне существующим типам животных — червям, медузам, морским перьям. Отмечалось, что симметрия и строение гипотетических «медуз» не напоминают ни одну из современных медуз. Аналогичным образом стебель «морских перьев» был не прямым, а зигзагообразным, как у чарнии (не наблюдается ни у одного из современных морских перьев). Симметрия и строение большинства «червей» не встречаются ни у каких современных групп червей, не говоря о том, что на окаменелостях нет и следа пищеварительного тракта или других систем, которые есть у всех червей.
Обращая внимание на эти странности строения, палеонтологи предлагают более оригинальные объяснения эдиакарских окаменелостей. Некоторые, например Адольф Зейльахер, работавший в Йельском и Тюбингенском университетах, полагают, что эти окаменелости не сопоставимы с нынешними животными. На самом деле, считал Зейльахер, это были «экспериментальные образцы» многоклеточной жизни, и симметрия их тел не имеет ничего общего со строением современных организмов. Он назвал эту фауну вендозоа, или вендской биотой. В русскоязычных источниках весь поздний докембрий, к которому относятся данные окаменелости, называется вендской эпохой, хотя международные геологические организации сейчас именуют этот период эдиакарским. Зейльахер отмечал, что по строению эти существа напоминали наполненные водой «подушки» волнистой формы; у них не было ни малейших признаков центральной нервной системы или пищеварительного тракта, которые есть даже у самых примитивных червей. Таким образом, эдиакарские существа вполне могли не использовать органы, относимые нами к пищеварительной, дыхательной или нервной системе. Напротив, эти «водяные подушки» обладали максимальной поверхностной площадью относительно собственного объёма, и за счёт «волнистости» эта площадь увеличивалась. Они всасывали все питательные вещества и кислород через складчатую «кожу» и аналогичным образом избавлялись от продуктов обмена веществ.
Марк Макменамин из колледжа Маунт-Холиок предложил гипотезу сада Эдиакары. По его мнению, огромная поверхностная площадь существ (по сравнению с их объёмом) позволяла им удерживать в тканях большое количество цианобактерий или истинных водорослей в качестве симбионтов. Эти фотосинтезирующие симбионты могли производить большое количество кислорода, поглощая углекислый
газ, — именно такую роль в современных коралловых рифах играют водоросли, растущие на гигантских двустворчатых моллюсках и других морских организмах. Грегори Реталлак из Орегонского университета, специализирующийся на изучении ископаемых почв, считает, что эти существа в основном являлись лишайниками и грибами, а не животными или растениями. Недавно он предположил, что эти структуры могут быть просто рельефом почвы.
Итак, существуют разные мнения о природе загадочных тварей, существовавших на заре биологии. Некоторые считают их обычными медузами, червями и морскими перьями, но большинство учёных полагает, что эти ископаемые не похожи ни на какие ныне живущие организмы. Являются они по-настоящему уникальной экспериментальной фауной — так называемой вендской биотой, некими крупными эндосимбиотическими организмами, лишайниками или почвой, — пока вопрос остаётся открытым. В конце концов, это лишь отпечатки на мягком песчаном или илистом морском дне. Мы очень приблизительно представляем себе их объёмное строение, как могли выглядеть поверхностные ткани, уже не говоря о внутренностях или жёстких частях тела. В этом и проблема: без жёстких частей тела существа с трудом могли сохраниться в ископаемой летописи. Останки часто сворачивались, дробились или повреждались именно потому, что принадлежали водянистым существам, напоминающим медуз.
Каковы бы ни были эти существа, важно помнить: они, без сомнения, демонстрируют, что скачок от одноклеточной к многоклеточной жизни произошёл около 630 млн лет назад. Их биоразнообразие увеличивалось по мере того, как наша планета разогревалась, миновав стадию «Земля-снежок», когда огромные ледяные щиты покрывали нашу планету от полюсов до экватора. В течение следующих 90 млн лет они были, пожалуй, единственными формами жизни на Земле, пока к концу их господства не стали появляться крошечные моллюски (см. главу 3). Затем они резко сошли со сцены, проиграв борьбу сначала примитивным моллюскам, а затем трилобитам. Примерно 500 млн лет назад эдиакарская фауна полностью вымерла, оставив биологам одни загадки.
Лучше один раз увидеть!
Первый образец
Charnia и другой окаменелости,
Charniodiscus, очень похожей на
Charnia и описанной Тревором Фордом в 1958 году, экспонируются на почётном месте в Нью-Уолкском музее города Лестер в Англии.
Эдиакарские окаменелости экспонируются в очень немногих музеях, поскольку они не так привлекательны или роскошны, как скелеты динозавров. В США среди таких музеев следует упомянуть Денверский музей природы и науки, Музей естественной истории под открытым небом (Чикаго) и Смитсоновский институт (Вашингтон, округ Колумбия). Во многих австралийских музеях есть экспонаты с хребта Флиндерс, особенно в Музее Южной Австралии (Аделаида) и в Музее Западной Австралии (Перт). Кроме того, они есть в Экологическом резервате Мистейкн-Пойнт на полуострове Авалон (Ньюфаундленд) и в Музее естественной истории Зенкенберг (Франкфурт-на-Майне, Германия).
Дополнительная литература
Attenborough, David, with Matt Kaplan. David Attenborough’s First Life: A Journey Back in Time. — New York: HarperCollins, 2010.
Glaessner, Martin F. The Dawn of Animal Life: A Biohistorical Study. — Cambridge: Cambridge University Press, 1984.
Knoll, Andrew H. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. — Princeton, N. J.: Princeton University Press, 2003.
McMenamin, Mark A. S. The Garden of Ediacara. — New York: Columbia University Press, 1998.
Narbonne, Guy M. The Ediacara Biota: A Terminal Neoproterozoic Experiment in the Evolution of Life //
GSA Today. — 1998. — Vol. 8. — P. 1–6.
Schopf, J. William. Cradle of Life: The Discovery of Earth’s Earliest Fossils. — Princeton, N. J.: Princeton University Press, 1999.
Seilacher, Adolf. Vendobionta and Psammocorallia: Lost Constructions of Precambrian Evolution //
Journal of the Geological Society, London. — 1992. — Vol. 149. — 607–613.
Seilacher, Adolf. Vendozoa: Organismic Construction in the Proterozoic Biosphere //
Lethaia. — 1989. — Vol. 22. — P. 229–239.
Valentine, James W. On the Origin of Phyla. — Chicago: University of Chicago Press, 2004.
03. Мелкие раковинные ископаемые
Первые раковины
Клаудина
Волна открытий, заставляющих пересмотреть историю самого раннего кембрия, началась после Второй мировой войны, когда в бывшем СССР большие группы учёных привлекли к исследованию геологических ресурсов Сибири. Там над сравнительно толстыми сериями докембрийских осадочных пород залегают более тонкие раннекембрийские осадочные формации, нетронутые более поздними процессами горообразования (в отличие от кембрийской складчатости в Уэльсе). Эти породы красиво обнажаются по берегам Лены и Алдана, а также в других частях этого обширного и малонаселённого региона. Группа сотрудников Московского палеонтологического института во главе с Алексеем Розановым из Московского палеонтологического института открыла, что в древнейших известняках кембрийского периода содержатся всевозможные мелкие неизвестные скелеты и фрагменты скелетов, размеры большинства из которых не превышают 1 см. Эти окаменелости получили звучные латинские наименования, а позже были объединены под общим термином «мелкие раковинные ископаемые».
Дж. Джон Сепковски-младший. «Foundations: Life in the Oceans»
Строители раковин
В главе 1 мы узнали, что первым ответом на вопрос Чарльза Дарвина о «кембрийском взрыве» стало открытие бактериальных матов, названных строматолитами и существовавших уже 3,5 млрд лет назад, а в конечном итоге — и микроокаменелостей цианобактерий, и прочих микроорганизмов из пластов той же эпохи. В главе 2 мы узнали, как одноклеточная жизнь породила многоклеточных мягкотелых существ из эдиакарской фауны. Но как же организмы, обладающие раковинами? Откуда они взялись?
Проблема с отращиванием жёсткой оболочки (
биоминерализация) не так проста, как может показаться. Большинству животных очень сложно извлекать из морской воды ионы кальция и карбонатов либо кислород и кремний, а затем выделять их для постройки известняковых или силикатных раковин. Чтобы происходила биоминерализация, требуются специальные биохимические пути; обычно этот процесс очень энергоёмкий.
Толстая раковина двустворчатого или одностворчатого моллюска отращивается из мясистой прослойки, так называемой мантии, которая находится прямо под раковиной и окружает мягкие ткани животного. В этом органе есть специализированные структуры и физиологические механизмы, позволяющие извлекать из морской воды ионы кальция и карбонатов, а затем превращать их в кристаллы карбоната кальция. Моллюски могут выделять это вещество в виде двух минералов: кальцита, широко распространённого в большинстве известняков, и арагонита (также называемого перламутром), которым большинство моллюсков выстилают внутреннюю часть своих раковин. Именно поэтому на внутренней стороне большинства ракушек заметен переливчатый жемчужный блеск — например, у морских ушек. Именно в ходе этого процесса образуются жемчужины, столь ценимые собирателями драгоценностей. По сути жемчужина — слоистое арагонитовое образование, скапливающееся вокруг центрального ядра (например, песчинки), попадающего в мантию некоторых моллюсков. Благодаря арагонитовому покрытию песчинка перестаёт раздражать мантию.
Исходя из длительного существования эдиакарской фауны (более 100 млн лет), мы можем заключить, что крупные мягкотелые организмы очень долго и прекрасно обходились без жёстких раковин. Судя по данным молекулярных часов, позволяющих судить о времени расхождения крупных таксонов, большинство основных типов (губки; стрекающие (медузы), анемоны; черви; кольчатые черви; плеченогие (брахиоподы) и моллюски) существовали в мягкотелой форме ещё в разгар эдиакарского периода, задолго до того, как обзавелись раковинами, после чего тела животных стали ещё более разнообразными. Но если раковина — бремя, зачем её отращивать? Как правило, раковина является защитой от хищников. Многие палеонтологи настаивали, что раковины начали возникать в качестве адаптации к новым хищным обитателям планеты, истребившим всех уязвимых, незащищённых мягкотелых тварей. Некоторым животным раковина служит резервуаром необходимых химических веществ. Отдельные моллюски с её помощью выделяют в окружающую среду избыточные отходы метаболизма.
Важнее всего, что минерализованные раковины способствуют диверсификации строения тела, а значит, повышают экологическое разнообразие и приспосабливаемость. Некоторые сохранившиеся до наших дней безраковинные моллюски (в частности, соленогастры) внешне напоминают червей. Но после обретения раковины среди моллюсков развились такие разнообразные и характерные формы, как хитоны, двухстворчатые, устрицы, гребешки, лопатоногие, блюдечки, морские ушки, улитки, каракатицы, кальмары и наутилусы (кораблики). Среди моллюсков есть тихоходные и примитивные, например блюдечки и морские ушки, ползающие среди камней в приливных заводях и питающиеся водорослями; есть безголовые двухстворчатые моллюски-фильтраторы. К моллюскам же относятся исключительно умные и быстрые хищники — осьминоги, кальмары, каракатицы.
Появление «ракушечек»
Сравнительно позднее возникновение раковин, спустя более 100 млн лет эволюции крупных мягкотелых животных, подсказывает, что отращивание раковины — непростой процесс. Аналогично, стоило ожидать, что большая раковина не могла появиться сразу. Это подтверждает и палеонтологическая летопись.
На протяжении длительного периода времени не было известно о каких-либо животных, более примитивных, чем трилобиты, обитавшие в раннем кембрии (см. главу 4). Некоторым казалось, что «внезапное возникновение» трилобитов с их сложными членистыми раковинами из белкового хитина, армированными кальцитом, свидетельствует: трилобиты (равно как и другие группы многоклеточных панцирных животных) возникли спонтанно, а не произошли от предшественников. Когда-то такое явление именовалось «кембрийским взрывом».
Вскоре после Второй мировой войны СССР бросил значительные силы на геологическую разведку отдалённых регионов, в том числе Сибири. В основном это делалось в экономических целях: искали полезные ископаемые, в частности уголь, нефть, уран и металлы. Одновременно была выполнена базовая работа по геологическому картированию этих областей и сбору окаменелостей. По берегам Лены и Алдана — больших рек, текущих на север и относящихся к бассейну Северного Ледовитого океана, удалось обнаружить гораздо более полные серии кембрийских и эдиакарских пород, чем найденные к тому времени в других частях света. Вскоре советские учёные приступили к описанию раннего отрезка кембрия,
до появления трилобитов, возникших только на третьем этапе кембрийского периода (этот ранний кембрийский ярус в советской науке именовался атдабанским). Два самых ранних кембрийских яруса, находящихся ниже отложений с древнейшими трилобитами, назвали немакит-далдынским и томмотским.
Хотя в этих породах не нашли трилобитов, в них обнаружили окаменелости других распространённых групп раковинных кембрийских существ — в частности губок, похожих на них (ныне вымерших) археоциатов и брахиоподов (плеченогих). Наиболее частыми находками были крошечные (не более 5 мм в диаметре) окаменелости, названные мелкими раковинными ископаемыми или мелкой раковистой фауной. Эти миниатюрные образцы было сложно найти, если только сборщик окаменелостей не знал точно, что именно искать. Неудивительно, что такие ракушки десятилетиями ускользали от внимания геологов, привыкших искать крупных эффектных трилобитов. Как правило, плотная концентрация этих крошечных существ наблюдалась в ракушечных горизонтах (рис. 3.1). Собирать их как цельные образцы в полевых условиях не представлялось возможным. Напротив, было гораздо проще доставить куски фоссилизированной породы в лабораторию и медленно выделить окаменелости из камня с помощью кислоты. В других случаях куски фоссилизированного известняка нарезались тонкими пластами и стачивались в тонкие каменные срезы толщиной не более 30 микрон, после чего наклеивались на предметное стекло микроскопа. Под микроскопом эти известняки оказывались нашпигованы разнообразными мелкими и сложными окаменелостями (рис. 3.2).
 Рис. 3.1.
Рис. 3.1. Типичная собранная вручную проба с крошечными раковинными включениями, заметными на выветренной поверхности тёмной полосы, пролегающей посередине. Образец из формации Вуд-Каньон в Белых горах (Уайт-Маунтинз) близ Лайды, штат Невада (фотография автора)
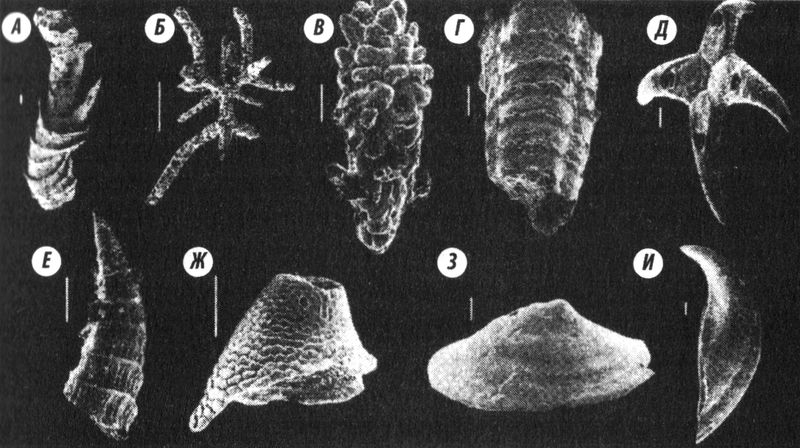 Рис. 3.2.
Рис. 3.2. Породы из самых ранних кембрийских ярусов (немакит-далдынского и томмотского) не содержат останков трилобитов, но наполнены крошечными фосфатосодержащими окаменелостями, так называемой мелкой раковистой фауной. Некоторые из этих раковин могут принадлежать моллюскам (
Д,
З,
И), тогда как другие, очевидно, были спикулами губок или фрагментами сетчатого панциря более крупных существ, в частности червей: (
А) Cloudina hartmannae, одна из древнейших известных скелетных окаменелостей, из тех же отложений, к которым относятся эдиакарские окаменелости, найденные в Китае; (
Б) спикула известковой губки; (
В) спикула, возможно, принадлежавшая кораллу; (
Г) Anabarites sexalox, трубчатое животное с трёхлучевой симметрией тела; (
Д) спикула, возможно, принадлежавшая древнему моллюску; (
Е) Lapworthella, конусообразный организм неизвестной систематики; (
Ж) скелетная пластина Stoibostromus crenulatus, организма неизвестной систематики; (
З) скелетная пластина Mobergella, возможно, относившегося к моллюскам; (
И) капсулообразная раковина Cyrtochites, возможно, относившегося к моллюскам. Масштабные полоски = 1 мм (фотографии предоставлены S. Bengston)
Когда открыли мельчайшие окаменелости, было неясно, к каким группам известных животных они относятся. Некоторые явно напоминали раковины двустворчатых или одностворчатых моллюсков. Другие казались фрагментами сетчатого панциря, покрывавшего тела более крупных существ. Встречалось множество игольчатых или стержнеобразных элементов, так называемых спикул, которые тесно переплетались между собой, образуя единственные жёсткие органы, имеющиеся у губок.
Важно, что многие из них состоят из фосфата кальция (минерала апатита), а не карбоната кальция, из которого строят раковины морские животные. Наряду с примером древнейших брахиопод (лингулоидных), использовавших фосфат кальция для строительства раковин, это позволяет предположить, почему потребовалось так много времени для развития крупных раковинных животных, в частности трилобитов. Видимо, пришлось преодолеть немало препятствий и сложностей, прежде чем в раннем кембрии был запущен процесс минерализации раковин. Во-первых, ни одно из этих крошечных существ не могло секретировать более нескольких десятков тонких слоёв раковины — они не были готовы обзавестись столь крупными раковинами, как трилобиты. Во-вторых, ряд соображений химического характера, наряду с изобилием раковин, состоящих из фосфата кальция (а не карбоната кальция), свидетельствует, что на тот момент содержание кислорода в атмосфере и океанах ещё не достигло уровня 21%, наблюдаемого на нашей планете сегодня. Предполагается, что уровень кислорода был гораздо ниже, что осложняло работу геохимических и физиологических механизмов, которые позволили бы моллюскам выделять минералы для своих раковин.
Прогнозы Престона Клауда
Дисциплина докембрийской геологии и палеонтологии практически не существовала до 1954 года, когда Стэнли Тайлер и Эльсо Баргхорн открыли первые факты о микроскопических докембрийских окаменелостях и опубликовали информацию о них. Один человек стал выдающимся первопроходцем и авторитетом в сфере докембрийской биологии и геологии, начиная с 1950–1960-х годов и вплоть до своей смерти. Это Престон И. Клауд. За свою научную карьеру я несколько раз встречался с Престоном, и, как вспоминается не только мне, но и Уильяму Дж. Шопфу в его книге «Cradle of Life» («Колыбель жизни»), Клауд возвышался над местностью как башня, хоть и был ростом всего метр шестьдесят семь, обладал сверкающей лысиной и щетинистой бородой. Но он (цитируя Шопфа) был «гигантом, неутомимым чудом, полным энергии, идей, мнений и трудолюбия. Возможно, он был величайшим учёным, работавшим на стыке биологии и геологии, какого дали миру Соединённые Штаты... Клауд не любил пустой болтовни и поражал коллег некоторым высокомерием (кое-кто за глаза называл его “маленьким генералом”, но никогда не говорил этого в лицо). Но одно достоинство Клауда перевешивало любые его недостатки: он был превосходен».
Клауд сделал хорошую карьеру в научной сфере, особенно преподавая в Калифорнийском университете в Санта-Барбаре и работая в Геологической службе США, где превратил палеонтологию в настоящий локомотив. Новаторское и масштабное мышление Клауда позволило ему стать экспертом во многих сферах, от исследования брахиопод и добычи бокситов до океанографии, изучения коралловых рифов и петрологии карбонатов. В 1974 году он стал писать книги, в которых предостерегал о будущем нашей планеты, исчерпаемости ресурсов и пике нефтедобычи, об антропогенных экологических катастрофах на Земле. Две главные книги на эту тему («Cosmos, Earth and Man: A Short History of the Universe» (1978) и «Oasis in Space: Earth History from the Beginning» (1988)) впервые объединили его масштабные представления о 4,5-миллиардной истории Земли с прогнозами о том, как люди рискуют уничтожить свою планету.
Задолго до того, когда кто-либо стал разрабатывать факты существования древнейшей жизни, Клауд активно изучал докембрийские микроокаменелости и строматолиты, а также искал новые эдиакарские ископаемые. Ещё важнее, что он создал парадигму для понимания докембрийской Земли — эпохи длиной в 3 млрд лет, когда уровень кислорода был низок, медленно эволюционировала одноклеточная жизнь, а затем последовало взрывное распространение эукариотических клеток, вызванное «кислородной катастрофой» между 2 и 1,8 млрд лет назад. Клауд выдвинул много инновационных идей о докембрийской геохимии, составе докембрийской атмосферы и океанов. Его знаменитая работа «A Working Model of the Primitive Earth» (1972) стала основой почти для всех исследований докембрия, проведённых в течение следующих сорока с лишним лет.
Клаудина
Клауду, как и многим другим геологам, не давала покоя большая разница между крупными, но мягкотелыми эдиакарскими созданиями и панцирными трилобитами. В конце жизни он очень обрадовался открытию и описанию мелких раковинных ископаемых раннего кембрия, которые в значительной степени восполнили этот пробел. Но всё-таки почему в докембрии не встречаются раковинные окаменелости? Как мог возникнуть такой эволюционный разрыв между эдиакарской и мелкой раковистой фауной?
В 1972 году Герард Гермс описал окаменелости из свиты Нама в Намибии (на тот момент данная территория являлась колонией ЮАР и называлась Юго-Западной Африкой), относящиеся к позднему докембрию. Он сообщил о странном известковом ископаемом около 6 мм в поперечнике и около 150 мм в длину. Ископаемое состояло из нескольких вложенных друг в друга конических оболочек, а внутри была трубчатая полость (рис. 3.3). До сих пор нет единого мнения о том, к какой группе животных они относятся (может, это были черви, выделявшие особые вещества для постройки трубчатого скелета) и можно ли их причислить к какой-то современной группе. Обычно усматривается связь между этими организмами и строматолитами: они должны были тяготеть к мелководной среде обитания, где имелись бактериальные маты. Кроме того, есть свидетельства о существовании других организмов, охотившихся на них, — первых настоящих хищников.
 Рис. 3.3.
Рис. 3.3. Реконструкция клаудины. Показана внешняя структура, напоминающая вложенные друг в друга конусы. Кроме того, изображена цилиндрическая камера, которую занимал мягкотелый строитель этой раковины (рисунок Mary P. Williams на основе нескольких источников)
Кем бы ни были эти таинственные существа, они стали первыми жителями Земли, которые обзавелись раковинами (наряду с китайскими трубчатыми окаменелостями,
Sinotubulites). Они появились в позднем докембрии, причём не только на территории Намибии, но и в Антарктике, Аргентине, Бразилии, Калифорнии, Канаде, Китае, Мексике, Неваде, Омане, Испании, Уругвае и особенно в России. В 1972 году Герме назвал их
Cloudina, отметив огромный вклад Престона Клауда в докембрийскую биогеологию. В последующие годы неоднократно возобновлялись споры об этих невероятно примитивных и фрагментарных окаменелостях, появлялись новые интерпретации. Однако кажется правильным, что древнейшие раковинные животные на Земле носят имя в честь Престона Клауда.
Тлеющий фитиль
«Кембрийский взрыв» скорее напоминал тлеющий фитиль (рис. 3.4). В период с 600 до 545 млн лет назад единственными многоклеточными существами на нашей планете были крупные мягкотелые эдиакарцы, лишённые раковин. Очевидно, геохимические условия (особенно низкий уровень кислорода) не давали возможности для эволюции крупных раковинных организмов. Наряду с таинственными эдиакарцами среди строматолитных матов обитали предки мелких раковинных ископаемых, в частности клаудины и синотубулиты.
 Рис. 3.4.
Рис. 3.4. Подробная схема стратиграфической истории окаменелостей, охватывающая поздний докембрий и кембрий, демонстрирует, что взрывного возникновения жизни в кембрии не было. В тот период жизнь развивалась поэтапно, в течение около 100 млн лет. Окаменелости первых крупных мягкотелых эдиакарских существ впервые появляются 600 млн лет назад, на вендском этапе позднего докембрия (см. рис. 2.2). К концу их господства мы находим первые мелкие раковинные организмы, в том числе примитивные конические клаудины и синотубулиты. На немакит-далдынском и томмотском этапах кембрия доминировали мелкие раковинные ископаемые, а также древнейшие брахиоподы, конические археоциаты, напоминающие губок. В этот же период наблюдается множество вертикальных следов, указывающих на широкое распространение червеобразных существ, не имевших жёсткого скелета. Наконец, на атдабанском этапе, около 520 млн лет назад, мы видим распространение трилобитов, а также значительную диверсификацию общего числа родов. Это известно благодаря минерализованным раковинам трилобитов, которые очень хорошо сохраняются (см. гистограммы в правой части схемы). Соответственно, «кембрийский взрыв» протекал на протяжении более 80 млн лет и не был внезапным событием даже по геологическим меркам (скопировано из работы
Donald R. Prothero and Robert H. Dott Jr. Evolution of the Earth, 7th ed. — Dubuque, Iowa: McGraw-Hill, 2004. — Fig. 9.14)
Затем, между 545 и 520 млн лет назад (в немакит-далдынском и томмотском периодах) крупнейшими существами на планете были мягкотелые организмы с крошечными минеральными включениями в коже или губки, свитые из мелких спикул, а также мелкие раковинные моллюски и брахиоподы. 520 млн лет назад, минимум через 80 млн лет после появления крупных многоклеточных организмов,
наконец появляются животные с большими кальцифицированными раковинами — трилобиты. Соответственно, не было никакого «кембрийского взрыва». Конечно, если не считать взрывом процесс длиной в 80 млн лет (от начала эдиакарского периода до атдабанского) или 25 млн лет (продолжительность двух первых этапов раннего кембрия).
Креационисты и прочие упорно игнорируют эти факты и искажают ископаемую летопись в собственных целях, пропагандируя ложную трактовку «кембрийского взрыва». Вот как описывает ситуацию Эндрю Кнолл:
«Был ли на самом деле кембрийский взрыв? Некоторые интерпретируют эту проблему сугубо в семантическом ключе — процесс, разворачивающийся на протяжении десятков миллионов лет, нельзя считать взрывным, а если кембрийская фауна не “взрывалась”, то, возможно, с ней вообще не происходило ничего экстраординарного. Определённо кембрийская эволюция не была карикатурно быстрой... Должны ли мы постулировать некий уникальный, но малопонятный эволюционный процесс, чтобы объяснить появление современных животных? Я так не думаю. Кембрийский период длился достаточно, чтобы достижения протерозоя могли совершиться без участия процессов, не известных популяционной генетике. 20 млн лет — долгий срок для животных, поколения которых сменяются каждые один-два года».
Дополнительная литература
Attenborough, David, with Matt Kaplan. David Attenborough’s First Life: A Journey Back In Time. — New York: HarperCollins, 2010.
Conway Morris, Simon. The Cambrian “Explosion”: Slow-fuse or Megatonnage? //
Proceedings of the National Academy of Sciences. — 2000. — Vol. 97. — 4426–4429.
Conway Morris, Simon. The Crucible of Creation: The Burgess Shale and the Rise of Animals. — Oxford: Oxford University Press, 1998.
Erwin, Douglas H., and James W. Valentine. The Cambrian Explosion: The Construction of Animal Biodiversity. — Greenwood Village, Colo.: Roberts, 2013.
Foster, John H. Cambrian Ocean World: Ancient Sea Life of North America. — Bloomington: Indiana University Press, 2014.
Grotzinger, John P., Samuel A. Bowring, Beverly Z. Saylor, and Alan J. Kaufman. Biostratigraphic and Geochronologic Constraints on Early Animal Evolution //
Science. — 1995. — October 27. — 598–604.
Knoll, Andrew H. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. — Princeton, N. J.: Princeton University Press, 2003.
Knoll, Andrew H., and Sean B. Carroll. Early Animal Evolution: Emerging Views from Comparative Biology and Geology //
Science. — 1999. — June 25. — 2129–2137.
Runnegar, Bruce. Evolution of the Earliest Animals //
J. William Schopf, ed. Major Events in the History of Life. — Boston: Jones and Bartlett, 1992. — P. 65-93.
Schopf, J. William. Cradle of Life: The Discovery of Earth’s Earliest Fossils. — Princeton, N. J.: Princeton University Press, 1999.
Schopf, J. William, and Cornelis Klein, eds. The Proterozoic Biosphere; A Multidisciplinary Study. — Cambridge: Cambridge University Press, 1992.
Valentine, James W. On the Origin of Phyla. — Chicago: University of Chicago Press, 2004.
04. Хочу я туда, где трилобитов стада...
Первые крупные панцирные животные
Оленеллус
Трилобиты напоминают о берегах древних морей, кишевших нарождающейся жизнью, когда тишину нарушали только вой ветра, шум прибоя и гром грозовых раскатов или вулканических извержений. Борьба за жизнь уже брала своё на просторах морей, но лишь законы природы и естественные события определяли судьбу развивающихся живых форм. Никаких следов было не найти на этих берегах, поскольку жизнь ещё не завоевала сушу. Геноцид ещё не был изобретён, жизни на Земле угрожали лишь кометы и астероиды. Все окаменелости — это своеобразные капсулы, уносящие нас в нашем воображении на невиданные берега, затерянные в море эпох, предшествовавших нашей. Время трилобитов невероятно удалено от нас, но всё-таки относительно небольшими усилиями мы можем выкопать его посланцев и подержать их в руке. А если мы изучим их язык, то поймём, что они могут нам рассказать.
Рикардо Леви-Сетти. «Trilobites»
Посланцы далёких времён
Одни из самых популярных окаменелостей как среди коллекционеров-любителей, так и у профессиональных палеонтологов — это трилобиты. Они существовали в период с 550 до 250 млн лет назад и в течение этих 300 млн лет разделились более чем на 5000 родов и 15 000 видов, ныне полностью вымерших (рис. 4.1). Они были разнообразны: от крошечных
Acanthopleurella (всего 1 мм в длину) до гигантских
Isotelus rex (более 70 см в длину). Поскольку трилобитов относительно просто собирать во многих местах, а также по причине исключительного изобилия их остатков во всех залежах верхнего палеозоя (особенно кембрия), они часто составляют ядро многих любительских коллекций. Изумительно сложные контуры, изысканные орнаменты, причудливая анатомия глаз и прочих частей тела, а также удивительные черты — благодаря этому трилобиты так притягательны для большинства коллекционеров ископаемых.
 Рис. 4.1.
Рис. 4.1. Реконструкция двух трилобитов: так они могли бы выглядеть при жизни (иллюстрация предоставлена Nobumichi Tomura)
Это увлечение характерно не только для нашего времени. Силурийский трилобит, вставленный в амулет, обнаружили в пещере, где 15 000 лет назад жили люди. Кембрийский трилобит, найденный австралийскими аборигенами в кремнистом сланце, был унесён на большое расстояние и стал украшением для первобытного орудия труда. Индейцы юта любили инкрустировать свои амулеты окаменелостями трилобитов
Elrathia kingi, часто встречающимися в породах хребта Хауз-Рейндж на территории штата Юта. Они называли это существо тимпе ханитца пачави (водяной жучок в каменном доме). Окаменелости
Elrathia kingi попадались в этом регионе настолько часто, что их даже промышленно добывают экскаваторами и продают почти во всех минералогических магазинах и сувенирных магазинах с окаменелостями.
Ещё важнее, что трилобиты были первыми крупными панцирными организмами на Земле. Данные о времени генетической дивергенции ближайших родственников этих животных убедительно свидетельствуют, что в раннем кембрии встречались мягкораковинные трилобиты, а уже в атдабанском периоде, на третьем этапе раннего кембрия, у них развились минерализованные панцири (см. рис. 3.4). Возможно, причина в том, что в атмосфере наконец скопилось достаточно кислорода, чтобы трилобиты могли кристаллизовать кальцит в своих раковинах. Большинство организмов, существовавших ранее, были мягкотелыми, без жёстких органов, либо имели крошечные зачаточные раковины (см. главы 2 и 3). Следовательно, они фоссилизировались лишь в среде, которая способствовала консервации, а не разложению (см. главу 5). Мало того, что трилобиты обладали большими сложными панцирями, состоявшими из хитина (как крабы, омары, креветки, насекомые, пауки, скорпионы и прочие членистоногие), эта относительно мягкая и нестойкая оболочка была армирована слоями минерального кальцита. Соответственно, трилобиты фоссилизировались гораздо легче, чем другие обитатели кембрия, поскольку принадлежали к одной из немногих групп, имевших минерализованные раковины. Поскольку трилобиты с твёрдым панцирем появились в атдабане, они в избытке присутствуют в ископаемой летописи, и создаётся ложное впечатление, что в истории жизни на Земле между атдабанским и томмотским периодом произошёл «кембрийский взрыв» (см. рис. 3.4). На самом деле это было взрывное возникновение животных с минерализованными скелетами.
Благодаря изобилию фоссилизировавшихся трилобитов в залежах позднего кембрия удалось описать более 300 их родов из 65 семейств, что многократно превышает количество находок прочих групп окаменелостей, относящихся к тому времени. Практически в любых кембрийских отложениях большинство окаменелостей — трилобиты, поэтому палеонтологи отсчитывают кембрийское время по этапам их эволюции.
Кто такие трилобиты
Трилобиты — это древнейшие известные людям ископаемые членистоногие. К данному типу также относятся насекомые, пауки, скорпионы, ракообразные и другие существа (см. главу 5). Трилобиты обладают всеми характерными чертами этого типа. Подобно другим членистоногим, они имеют сегментированный экзоскелет, разлагающийся при линьке. Поэтому ископаемые часто представляют собой разные фрагменты перелинявшего экзоскелета, а не цельные останки животных, которые могли бы сохраниться достаточно долго, чтобы снова перелинять. Однако, в отличие от покрова большинства других членистоногих, хитиновый экзоскелет трилобитов армирован минерализованным кальцитом, поэтому они фоссилизировались лучше, чем насекомые, пауки, скорпионы и большинство ракообразных.
Голова трилобитов, вернее головной щит, называется
цефалоном (в переводе с греческого — голова; рис. 4.2,
А). Обычно это широкая структура с двумя «щеками» по бокам и центральной долей («носом») — так называемой
глабелью. По обе стороны глабели, как правило, располагались два глаза. Некоторые трилобиты имели крошечные глазки или вообще были их лишены (слепые или полуслепые). У других имелись огромные вращающиеся глаза, обеспечивавшие угол обзора в 360 градусов и позволявшие заметить любых хищников. У наиболее высокоразвитых трилобитов были линзы, состоявшие из двух кристаллов кальцита. Такая линза получалась двояковыпуклой и давала поправку на сферические аберрации, возникавшие в толстых увеличительных стёклах. Спустя 400 млн лет после того, как у трилобитов развились эти «устройства», их повторно изобрёл Христиан Гюйгенс — великий голландский учёный XVII века. Ещё важнее, что трилобиты, пожалуй, были первыми существами на Земле, которые имели настоящие глаза и полагались на зрение, отыскивая пищу и ускользая от хищников.
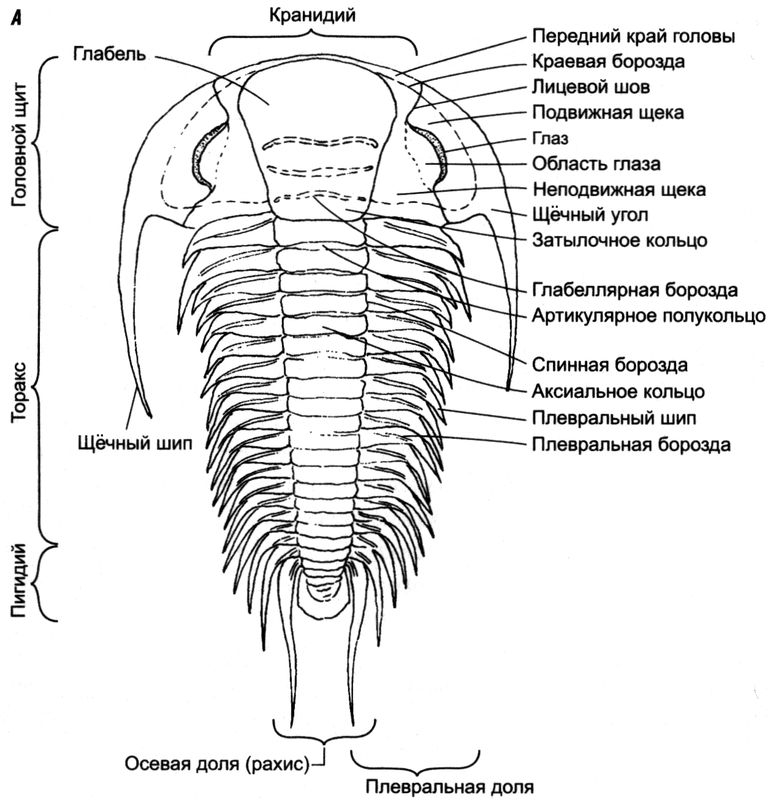

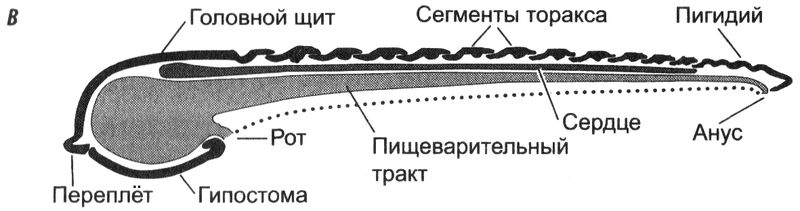 Рис. 4.2.
Рис. 4.2. Общая анатомия трилобита. (
А) цельный экзоскелет, вид сверху; (
Б) головной щит, вид снизу; (
В) поперечное осевое сечение, элементы скелета отмечены чёрным цветом (адаптировано из нескольких источников)
Щёчные области при линьке отходили от центральной части головы (кранидия), поэтому в большинстве окаменелостей трилобитов сохранилась только середина головного щита. Опытный специалист по трилобитам часто может определить вид этого животного по одному кранидию. Элементы глаз, глабели, контуры щёк, а также шипы по краям головного щита трилобита отличались большой изменчивостью. На передней части хорошо сохранившихся экземпляров находится пара «антенн», с помощью которых трилобиты находили путь в тёмной мутной воде. В нижней части тела трилобита имелась пластина, частично прикрывавшая рот; из-под неё выходили ротовые органы, позволявшие животному засасывать питательный ил и переваривать его (рис. 4.2,
Б). Большинство трилобитов питались наносами или фильтруемой грязью.
Средняя часть тела трилобита называется
торакс, как у большинства членистоногих. Она делилась на сегменты, за счёт чего животное могло изгибаться, ползать и сворачиваться для защиты. По бокам каждого сегмента были две доли (называвшиеся
плевральными), а одна такая доля (
рахис) шла по центральной оси тела. Благодаря продольному делению тела на три доли трилобиты получили своё название.
[2] Торакс некоторых из них состоял всего из двух-трёх сегментов, они не отличались гибкостью и почти всё время лежали плашмя. У других было много сегментов; такие животные могли сворачиваться в шар, чтобы отвадить хищников, — именно так сегодня делают равноногие ракообразные, к которым относятся всем известные мокрицы. На хорошо сохранившихся окаменелостях видно, что под каждым сегментом торакса имелась пара ножек и к каждой ножке крепилась пара жабр, напоминавших пёрышки.
Хвостовой отдел трилобита называется не брюшком, как у большинства членистоногих, а
пигидием (в переводе с греческого — хвостик). У большинства трилобитов последние сегменты торакса срастались в цельный большой пигидий, по форме напоминавший пластину. Однако оленеллиды были устроены совсем иначе.
Оленеллус и первые трилобиты
Когда вы присмотритесь к трилобитам и научитесь их различать, среди самых узнаваемых будет
Olenellus (рис. 4.3). В 1910 году его назвал и подробно изучил не кто иной, как Чарльз Дулиттл Уолкотт — один из первых экспертов по кембрию (см. главу 1). Наиболее характерная черта оленеллуса — то, что последние сегменты его тела не срослись в пигидий; более того, на хвосте есть длинный шип. Это довольно примитивный и неудивительный приём: оленеллиды являются древнейшими, известными людям, трилобитами.
 Рис. 4.3.
Рис. 4.3. Экземпляр оленеллуса с характерным D-образным головным щитом, булавовидным кончиком глабеллы, большими фасеточными глазами, напоминающими полумесяц, шипами по краям головного щита и на некоторых сегментах торакса, а также без большого цельного пигидия (хвостового сегмента) (иллюстрация из фонда Wikimedia Commons)
Наряду с отсутствием пигидия оленеллус обладает и многими другими реликтовыми чертами. Головной щит у него крупный и по форме напоминает букву D. В верхней части головного щита отсутствуют лицевые швы. Два больших полулунных глаза почти примыкают к бороздчатой глабелле, которая спереди имеет булавовидный вырост. Глаза устроены просто и состоят из множества крошечных, тесно сгруппированных известковых стержней; напоминают типичные фасеточные глаза большинства насекомых и других членистоногих. Такие глаза не могли дать чёткую фотографическую картинку, но с их помощью трилобит мог различать крупные светлые и тёмные области, а также движение поблизости от себя. Изучение экстраординарных образцов кембрийской фауны — например, таких, как сохранились в сланцах Бёрджесс (Канада) или в Маотяньшаньских сланцах (Китай), свидетельствует, что в те времена почти не было крупных плотоядных (см. главу 5). Самым большим хищником, вероятно, был метровый аномалокарис. Окаменелости явно демонстрируют, что он кусал трилобитов, обнаруживаемых в среднекембрийских сланцах Бёрджесс, где открыли этого хищника. Но, по сравнению с поздним палеозоем, пресс хищников был незначительным, и в кембрийском периоде трилобиты оставались сравнительно примитивными, неспециализированными. Вплоть до ордовика не встречается суперхищников, вроде наутилоидеи с раковинами длиной более 6 м. Лишь тогда у трилобитов развились внушительные раковины, приспособленные для зарывания в грунт, плавания и скатывания в шар для защиты от по-настоящему грозных противников.
Ещё одна примечательная черта оленеллуса — множество шипов по краям панциря. Как правило, из заднего угла цефалона у животных выходили шипы (
щёчные шипы). У многих были широкие шипы, отходящие от торакальных сегментов и загибающиеся назад (обычно речь идёт о сегменте 3). У некоторых имелись дополнительные шипы, торчавшие из цефалона вперёд; они зачастую помогают палеонтологам распознавать роды и виды оленеллидов.
После возникновения на атдабанском этапе раннего кембрия (около 520 млн лет назад) оленеллиды процветали, разделившись на множество родов и видов, окаменелости которых находят почти по всему миру, как в атдабанском ярусе, так и в следующем за ним ботомском ярусе раннего кембрия. Затем, в конце тойонского этапа (около 509 млн лет назад) они исчезли. Брюс Либерман из Канзасского университета проанализировал тысячи образцов оленеллидов и заключил, что их предки возникли в раннем кембрии на территории нынешней Сибири или западной России. Но, как и другие трилобиты, они не обызвествлялись и не фоссилизировались вплоть до атдабанского периода.
Что произошло с трилобитами
На протяжении оставшейся части палеозоя трилобиты исчезли постепенно в результате ряда массовых вымираний (рис. 4.4). К ним относятся и многочисленные малые вымирания позднего кембрия, когда несколько последовательных катастроф одна за одной свели на нет биоразнообразие трилобитов. В ордовикском периоде они впервые схлестнулись с крупными хищниками; возможно, это были гигантские наутилоидеи. Вскоре трилобиты стали более специализированными, а значит, не такими похожими друг на друга. Они приспосабливались и становились менее уязвимыми для хищников. Тем временем менялась форма их тел и образ жизни. К числу адаптаций относится способность зарываться в грунт (ею обладали плоские трилобиты асафиды (
Asaphida) или иллениды (
Illaenida), по форме напоминавшие ковш), сворачиваться в шар (калимениды;
Calymenida) или просто уменьшение до крошечных размеров (тринуклеиды;
Trinucleida), в частности криптолитус (
Cryptolithus) — трилобит, похожий на кружевной воротничок, не больше ногтя). Затем наступило позднеордовикское вымирание (около 450 млн лет назад), и лишь несколько эволюционных линий трилобитов сохранились до силура и девона. Последний расцвет трилобитов произошёл в девонском периоде, когда были повсеместно распространены факопиды (
Phacopida) с фасеточными глазами, а также крупные шипастые трилобиты, в частности тератаспис (
Terataspis) длиной около полуметра. Вымирания, произошедшие в позднем девоне (375 и 357 млн лет назад), положили конец всем отрядам трилобитов, кроме относительно мелких и примитивных проэтидов, которые просуществовали ещё 125 млн лет.
 Рис. 4.4.
Рис. 4.4. Диверсификация и вымирание трилобитов (по нескольким источникам)
Трилобиты окончательно исчезли во время Великого пермского вымирания, случившегося около 250 млн лет назад, — крупнейшего массового вымирания в истории Земли, когда исчезло 95% всех морских видов. Эта колоссальная катастрофа погубила не только последних уцелевших трилобитов, но и две господствовавшие группы палеозойских кораллов (табулятов и ругозов), бластоидей (родственных криноидеям, «морским лилиям») и фораминифер-фузулинид (многочисленных простейших, раковины которых по форме напоминали рисовые зёрнышки). Споры о том, что могло повлечь «величайшее из всех массовых вымираний», продолжаются. Однако современные данные указывают на то, что крупнейшие в геологической истории вулканические извержения, при которых образовались так называемые сибирские траппы (лавовые потоки), спровоцировали стремительный парниковый эффект; из-за него океаны разогрелись и стали слишком кислыми для существования разнообразной жизни. При этом атмосфера оказалась перенасыщена углекислым газом, а кислорода в ней было недостаточно. Эти и некоторые другие катастрофические события уничтожили почти все биологические виды, существовавшие на Земле.
Лучше один раз увидеть!
Во многих музеях экспонируются окаменелости трилобитов, хотя в большинстве вы не найдёте хороших и полных экземпляров оленеллидов. Среди них Денверский музей природы и науки, Музей естественной истории под открытым небом (Чикаго), Геологический музей висконсинского университета в Мэдисоне, Смитсоновский институт (Вашингтон, округ Колумбия).
Оленеллиды так широко распространены в некоторых регионах мира, что их легко собирать самостоятельно. Вот некоторые из известных и легкодоступных мест в США. Подробнее о них можно почитать в книгах для собирателей окаменелостей и на специализированных сайтах.
Мраморные горы (Калифорния). Едете по федеральной автомагистрали 40 (на восток или на запад), пока не доберётесь до трассы 78 Келбейкер-Роуд. Сверните на неё и проедьте ещё милю на юг до Т-образного перекрёстка. Поверните налево (на восток) и минуйте заброшенный городок Чемблесс вдоль Нэшнл-Трейлз Хайвей (бывшее американское шоссе 66). Поверните на юго-запад к Кадису. После того как асфальтированная дорога повернёт на восток, и чуть не доезжая до её изгиба к югу (там она пересекает железнодорожные пути), сверните влево на грунтовую дорогу, ведущую на север. По грунтовой дороге — около мили на север, пока не доберётесь до перекрёстка с заезженной грунтовой дорогой, идущей с запада на восток; далее поверните на восток. Проехав с полмили по этой дороге, увидите ещё одну грунтовую дорогу, ведущую на северо-восток к старому карьеру.
Двигайтесь по ней, пока она будет проезжей, а затем дойдите до Лэзем-Шейл (толща бурых сланцев) под серым скалообразующим известняком Чемберлесс. Осмотрите старые открытые выработки серьёзных коллекционеров, переворачивайте большие куски сланца. Увидите много хороших головных щитов любого размера, хотя цельные трилобиты встречаются исключительно редко. Два хороших сайта по теме — «Трилобиты в Мраморных горах, пустыня Мохаве, Калифорния» (
http://inyo.coffeecup.com/site/latham/latham.html) и «Трилобиты Лэзем-Шейл, Калифорния» (
http://www.trilobites.info/CA.htm).
Перевал Эмигрант-Пасс, гряда Нопах-Рейндж (Калифорния). Едете по федеральной автомагистрали 15 к Бейкеру (штат Калифорния). Сверните с магистрали на шоссе 127 штата Калифорния (дорога к Долине Смерти), проделайте по нему 48 миль до Старого Испанского тракта. Поверните направо на Старый Испанский тракт и езжайте через Теокопу к перевалу Эмигрант-Пасс. Породы выходят на поверхность прямо к западу от верхней точки перевала, на южной стороне дороги (координаты GPS: 35.8856N, −116.0603W). Два хороших сайта по теме — «Трилобиты Нопах-Рейндж, округ Иньо, Калифорния» (
http://inyo.coffeecup.com/site/cf/carfieldtrip.html) и «Трилобиты-оленеллиды с Эмигрант-Пасс, Калифорния» (
http://donaldkenney.x10.mx/SITES/CANOPAH/CANOPAH.HTM).
Пик Оук-Спринг, округ Линкольн (Невада). Из Кальенте (штат Невада) едете 10 миль на запад или 33 мили на восток по американскому шоссе 93, вплоть до перекрёстка с двумя шоссе штата Невада: 375 и 318 (между Хико и Эш-Спрингс). По северной стороне магистрали ищите поворот, отмеченный знаменитым указателем Бюро по управлению землями: «Oak Springs Trilobite Site». Поверните на север, езжайте по грунтовой дороге, припаркуйтесь на галечной стоянке и отправляйтесь пешком по Дороге Трилобитов от самого её начала через полынь, пока не дойдёте до ровной местности, усыпанной кусками сланцев Пиоше. Переворачивайте камни и найдёте много красивых головных щитов, иногда и более полные экземпляры разного возраста и размеров.
[3]
Дополнительная литература
Форти Р. Трилобиты. Свидетели эволюции. — М.: Альпина Нон-Фикшн, 2014.
Erwin, Douglas H., and James W. Valentine. The Cambrian Explosion: The Construction of Animal Biodiversity. — Greenwood Village, Colo.: Roberts, 2013.
Foster, John H. Cambrian Ocean World: Ancient Sea Life of North America. — Bloomington: Indiana University Press, 2014.
Lawrance, Pete, and Sinclair Stammers. Trilobites of the World: An Atlas of 1000 Photographs. — New York: Siri Scientific Press, 2014.
Levi-Setti, Ricardo. The Trilobite Book: A Visual Journey. — Chicago: University of Chicago Press, 2014.
05. Это червь или членистоногое?
Происхождение членистоногих
Галлюцигения
Существует гораздо больше видов беспозвоночных, чем позвоночных. По последним оценкам, на Земле насчитывается не менее 10 миллионов видов беспозвоночных, а возможно, и больше... Беспозвоночные доминируют на Земле и по чистой биомассе. Например, во влажном тропическом лесу близ Манауса в бразильской Амазонии на каждом гектаре обитает несколько десятков птиц и млекопитающих, но при этом более миллиарда видов беспозвоночных, абсолютное большинство которых — клещи и ногохвостки. На каждый гектар в этом районе приходится около 200 килограммов чистого веса животной ткани, 93% от этой величины приходится на беспозвоночных. Одни только муравьи и термиты дают около трети биомассы. Итак, если вы погуляете по тропическому лесу, да и почти по любому другому биотопу, если на то пошло, ваше внимание привлекут в основном позвоночные, хотя на самом деле вы окажетесь в мире, населённом прежде всего беспозвоночными.
Эдвард О. Уилсон. «The Little Things that Run the World»
Диковины сланцев Бёрджесс
Один из наиболее захватывающих регионов, известных своими окаменелостями, — легендарные сланцы Бёрджесс, расположенные в Скалистых горах близ Филда (Британская Колумбия). Их случайно открыл палеонтолог-первопроходец Чарльз Дулиттл Уолкотт (см. главу 1), работавший в этих краях со среднекембрийскими породами (возраст около 505 млн лет) летом 1909 года. 30 августа 1909 года его лошадь споткнулась о большой камень, лежавший на тропе. Уолкотт спешился, отбросил булыжник и обнаружил, что снизу тот покрыт окаменелостями, сохранившимися в виде тончайшей плёнки. Он проследил, с какой именно точки на склоне мог упасть этот камень, и вскоре начал большие раскопки (см. рис. 1.4). Каждое лето вплоть до 1924 года он возвращался на сланцы Бёрджесс, в итоге собрав коллекцию из более чем 65 тыс. экземпляров, которая была размещена в Смитсоновском институте. Почти все окаменелости из сланцев Бёрджесс принадлежат мягкотелым организмам, оказавшимся похороненными под подводными оползнями в период среднего кембрия. Дело не только в том, что их засыпало — придонные воды были явно бедны кислородом, из-за чего падальщики и редуценты до них не добрались. Поэтому в сланцах Бёрджесс сохранились нежные тонкие ткани, которые редко попадаются в палеонтологической летописи.
Но Уолкотт был слишком занят, руководя Смитсоновским институтом и выполняя многие другие свои обязанности. Поэтому он успел лишь поверхностно описать эти окаменелости, а в 1927 году умер. Многие неизученные окаменелости остались пылиться в ящиках шкафов. Те ископаемые, которые Уолкотт изучил и сделал о них публикации, были описаны и отнесены к известным типам животных, например к членистоногим или червям. Никто не уделил достаточно времени на тщательное препарирование экземпляров и изучение их мелких деталей.
Так окаменелости и лежали не один десяток лет. В 1949 году легендарный британский эксперт по трилобитам Гарри Уиттингтон принял кафедру в Гарвардском университете. Вскоре он осознал, что колоссальная коллекция образцов со сланцев Бёрджесс, хранящаяся у него в закромах, никогда толком не изучалась. Вернувшись в 1966 году в Англию и став вудвордским профессором палеонтологии в Кембриджском университете, Уиттингтон развернул масштабные работы с этими окаменелостями. Он и его студенты вернулись на место раскопок Уолкотта, добыли сотни новых экземпляров и, кроме того, дотошнее Уолкотта препарировали детали окаменелостей, зачастую копая и под ними, чтобы найти внизу объёмные структуры, которые мог упустить Уолкотт. В течение нескольких следующих лет Уиттингтон и его ученики (особенно Дерек Бриггс, специализировавшийся на животных, напоминавших членистоногих, а также Саймон Конвей Моррис, классифицировавший ископаемые неясного происхождения как древних червей) сделали революционные находки, которые Уолкотт просто не заметил.
Присмотревшись к фауне сланцев Бёрджесс и извлекая объёмные окаменелости, которые можно рассмотреть со всех сторон, приходишь к выводу, что строение многих из них совсем не напоминает организмы современных животных. Так, у опабинии (
Opabinia) было пять глаз на «морде» и длинное членистое тельце, а сама «морда» напоминала длинное сосательное рыльце (рис. 5.1). Крупнейший хищник аномалокарис (
Anomalocaris) достигал метра в длину, имел длинные разветвлённые ротовые придатки, членистое тело с плавниками по бокам и рот, по форме напоминавший дольку ананаса, но работавший по принципу диафрагмы в объективе фотоаппарата (Уолкотт ошибочно принял этот орган за медузу). Виваксия (
Wiwaxia) была небольшим чашевидным существом с рядом острых шипов на спине. Диномисх (
Dinomischus) напоминал более поздние раковинные криноидеи («морские лилии»), но был мягкотелым. Как указывали Уиттингтон, Бриггс и Конвей Моррис, многие из этих существ относились к самостоятельным типам и не могли быть причислены к современным группам, например к членистоногим или червям.
 Рис. 5.1.
Рис. 5.1. Окаменелости из сланцев Бёрджесс, в частности опабиния с длинным рыльцем — слева вверху и на заднем плане вверху (материалы из коллекции Смитсоновского института)
Наряду со всеми этими диковинами, разумеется, попадалось много мягкотелых существ, которые вполне походили на обычных креветок и других членистоногих. Кроме того, как и в других кембрийских залежах, здесь в изобилии встречались среднекембрийские трилобиты — единственные панцирные окаменелости сланцев Бёрджесс. Но их присутствие демонстрирует, насколько избыточно содержание этих организмов в большинстве палеонтологических находок и почему лишь трилобиты в большом количестве сохранились со времён кембрия. Без сланцев Бёрджесс и других мест с исключительно хорошо сохранившимися окаменелостями — таковы, например, Маотяньшаньские сланцы (Китай) и регион Сириус-Пассет (Гренландия), мы бы никогда не узнали, что когда-то дно океана населяли всевозможные странные и причудливые животные, обладавшие неизвестными вариантами строения тела. Ведь они были мягкотелыми и редко фоссилизировались.
В 1989 году Стивен Джей Гулд опубликовал научно-популярный бестселлер «Wonderful Life: The Burgess Shale and the Nature of History» («Живительная жизнь. Сланцы Бёрджесс и природа истории»). В основном книга рассказывает именно об этой формации и является первой работой о сланцах Бёрджесс, рассчитанной на широкую аудиторию. Гулд подробно описывает, как сильно работа Уиттингтона, Бриггса и Конвея Морриса изменила наши представления о природе этой фауны. Также он отметил, насколько заблуждался Уолкотт, стремившийся впихнуть всех животных в существующие типы. Действительно, сланцы Бёрджесс свидетельствуют не о постепенном развитии, диверсификации и распространении жизни со времён кембрия, а о том, что жизнь достигла максимального количества и разнообразия форм в среднем кембрии, а затем девонское вымирание положило конец почти всем эволюционным линиям, за исключением немногих уцелевших (членистоногих, моллюсков и некоторых других).
Гулд выдвинул и более серьёзную идею. С его точки зрения, сланцы Бёрджесс подчёркивали важность
случайности и счастливых совпадений, определивших ход всех следующих событий в истории жизни. Если в широкой перспективе рассмотреть странных животных, плававших в среднекембрийских морях, кто бы мог предположить, что все эти существа были «экспериментальными» животными, которым не суждено дожить даже до конца кембрия? Кто бы мог подумать, что крошечная неброская окаменелость под названием пикайя (
Pikaia) (см. главу 8) — представитель нашей эволюционной линии, позвоночных, которые в итоге станут хозяевами планеты (наряду с членистоногими)?
Если бы по какой-то случайности позвоночные вымерли в кембрии наряду с большинством других экспериментальных форм, как бы развивалась история жизни? Разумеется, не было бы никаких динозавров, а также млекопитающих. И людей. Всякий раз, когда мы перематываем плёнку биологической истории, она проигрывается иначе. Если бы столь случайные и непредсказуемые события, как падение астероида в Мексике и колоссальные извержения лавы в Индии 65 млн лет назад не истребили динозавров, млекопитающие никогда не стали бы крупнее, чем были 120 млн лет назад, в эпоху ящеров, и людей на Земле тоже не было бы. Современный мир — невероятная счастливая случайность, один из миллионов возможных сценариев развития жизни. Все живые организмы — не неизбежный результат долгосрочной эволюции, а потомки тех, кто уцелел после массовых вымираний и других катастроф.
В своей книге Гулд проводит аналогию с известным рождественским фильмом «It’s a Wonderful Life» («Удивительная жизнь») Фрэнка Капры с Джимми Стюартом и Донной Рид в главных ролях. В этом фильме герой Стюарта по имени Джордж Бейли получает возможность увидеть, каков был бы мир, если бы его, Джорджа, никогда не существовало. Герой обнаруживает, что каждая человеческая жизнь и любое незначительное событие влечёт непредсказуемые последствия.
Галлюцинация в камне
Среди наиболее странных и сложных для интерпретации окаменелостей сланцев Бёрджесс был «червь», которого Уолкотт отнёс к роду
Canadia класса полихет. Когда Конвей Моррис стал работать с разнообразными «червями», проигнорированными Уолкоттом, он обнаружил несколько экземпляров, стоявших особняком (рис. 5.2,
А). Они напоминали червей, то есть обладали характерным длинным тельцем, но также имели пары прямых острых выростов с одной стороны тела и нечто вроде ряда «ножек» или «щупалец» — с другой. На кончике тела у них был выцветший «пузырёк», напоминавший голову, — и этим дело не ограничивалось. Определённо, такое существо не напоминало ни одного червя на нашей планете (как ныне живущих, так и вымерших видов). При первой реконструкции у Конвея Морриса получилось тельце на парных прямых острых ножках, а по верхней части тела шёл ряд «щупалец» (рис. 5.2,
В). В 1977 году он переименовал эту окаменелость в галлюцигению (
Hallucigenia), поскольку считал, что такое существо может привидеться лишь в бреду или страшном сне.


 Рис. 5.2.
Рис. 5.2. «Червь» галлюцигения. (
А) окаменелость из сланцев Бёрджесс; (
Б) первая реконструкция, на которой шипы показаны как «ножки»; (
В) современная реконструкция, где шипы показаны на спине. ((
А–
Б) иллюстрации предоставлены S. Conway Morris (Кембриджский университет); (
В) иллюстрации предоставлена Nobumichi Tamura)
Другие учёные не разделяли его уверенности. Некоторые полагали, что эта окаменелость — просто конечность более крупного животного. Уже был известен случай с аномалокарисом, чьи ротовые придатки ошибочно приняли за существ, напоминавших креветок. Но преобладающая точка зрения была такова: галлюцигения могла принадлежать к лобоподам — морским червям с ножками, имевшим неясное происхождение и встречавшимся в раннепалеозойских породах по всему миру.
В 1991 году Ларс Рамскольд и Хоу Сянцань опубликовали работу, в которой описали ещё одну галлюцигениду, микродиктиона (
Microdictyon), найденную в нижнекембрийских Маотяньшаньских сланцах на территории Китая (рис. 5.3,
А). Этот образец сохранился гораздо лучше, чем иные ископаемые галлюцигении. В данном случае учёные убедились, что Моррис реконструировал галлюцигению вверх тормашками (рис. 5.2,
Б)! У микродиктиона по спинке шёл ряд парных шипов, и «ножки» галлюцигении Морриса оказались такими же шипами. Гибкие маленькие «придатки» на спине существа, изображённого Конвеем Моррисом, оказались его лапками, которые, как и полагается, были парными. Ещё удивительнее, что по телу микродиктиона шла полоса маленьких жёстких щитков, которые задолго до того были открыты на материале кембрийской мелкой раковистой фауны, но ранее никто не знал, какому существу они могли принадлежать.

 Рис. 5.3.
Рис. 5.3. Онихофоры и лобоподы. (
А) айшиайя из сланцев Бёрджесс, примитивная онихофора; (
Б) реконструкция некоторых ископаемых лобопод: (1) кардиодиктион, (2) люолишания, (3) галлюцигения, (4) паусиподия, (5) микродиктион, (6) ониходиктион (иллюстрации (
А–
Б) предоставлены S. Conway Morris (Кембриджский университет))
Микродиктион помог не только поставить галлюцигению с головы на ноги, но и разгадать другую тайну — их общее происхождение. Когда обоих животных расположили правильно, стало ясно, что это лобоподы, существа, обычные на кембрийском морском дне. В сланцах Бёрджесс были найдены окаменелости, которые сохранились ещё лучше и бесспорно относились к лобоподам. Речь об айшиайе (
Aysheaia) (рис. 5.3,
Б). Работая с относительно полными экземплярами, учёные наконец смогли выяснить, что представляли собой лобоподы. Оказалось, что это далёкие предки современного зоологического типа, представители которого ползают в джунглях, — онихофор, также называемых бархатными червями.
Кто такие членистоногие
Насекомые, пауки, скорпионы, раки, усоногие, мечехвосты и трилобиты — всё это представители членистоногих, крупнейшего зоологического типа на Земле. По любым меркам членистоногие являются доминирующими животными на нашей планете и всегда были таковыми — как бы нам ни хотелось считать хозяевами себя. Насчитывается более миллиона видов членистоногих (вероятно, больше их видов пока не открыто), что составляет более 85% как от всех видов животных — их 1,4 млн, — так и от общего количества особей (рис. 5.4). Существует почти 900 тыс. видов насекомых, из них более 340 тыс. одних жуков. Когда великого биолога Дж. Б. С. Холдейна спросили, что он может сказать о Творце на основании своего научного опыта, тот ответил: «Наверняка Бог был без ума от жуков». Для сравнения: тип хордовые, к которому относимся мы, включает менее 45 тыс. видов, причём их значительная часть — рыбы. Млекопитающих чуть более 4000 видов.
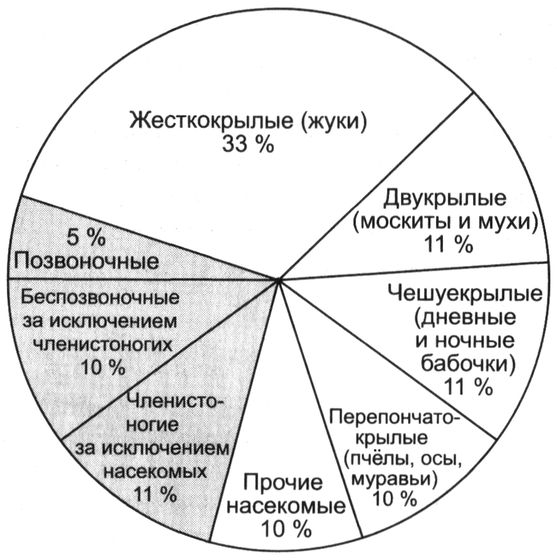 Рис. 5.4.
Рис. 5.4. Биоразнообразие животных и преобладание членистоногих, особенно насекомых (рисунок Pat Linse, на основании нескольких источников)
Если общее биоразнообразие вас не впечатляет, как насчёт численности? Членистоногие известны молниеносным размножением в благоприятных условиях. Вспомните нашествия саранчи; подумайте, с какой скоростью тли могут облепить растение; вообразите, сколько особей обитает в муравейнике или термитнике. Если всего пара тараканов станет бесконтрольно размножаться, уже через семь месяцев у неё может быть 164 млрд потомков! В тропиках на паре гектаров могут жить несколько десятков птиц и млекопитающих, но более миллиарда членистоногих, в том числе клещей, жуков, ос, мотыльков и мух. В единственном муравейнике может обитать миллион насекомых. В самых густонаселённых частях океана в кубометре воды живут миллионы особей крошечного зоопланктона (креветки, веслоногие, криль и ракушковые рачки).
Кроме того, членистоногие отличаются редкой приспособляемостью и могут занять практически любую экологическую нишу — кроме тех, где выгодно иметь большие размеры тела. Есть летающие членистоногие и членистоногие, обитающие в пресной и солёной воде; членистоногие, переносящие экстремальные температуры, которые могут быть как ниже точки замерзания воды, так и почти на точке её кипения. Многие членистоногие являются внешними и внутренними паразитами других организмов. Такая приспособляемость объясняется, прежде всего, анатомией животных. Эти существа состоят будто из отсеков, мелкие сегменты без труда «прибавляются» к членистоногому или «вычитаются» из него, а также легко меняют форму. Каждый сегмент оснащён парой суставчатых конечностей, которые могут быть «переоборудованы» в ротовые придатки, ножки, усики, клешни или хелицеры, плавательные органы, крылья и многие другие структуры. Наиболее характерная черта всех членистоногих — твёрдая внешняя оболочка, так называемый экзоскелет, под которым находятся мускулы и мягкие ткани. Скелет позвоночных, напротив, находится под покровом мышц и мягких тканей. Экзоскелет даёт разнообразные преимущества: защищает от хищников, служит водонепроницаемой оболочкой, которая в своё время позволила членистоногим переселиться из океана на сушу. Но жёсткая оболочка не растёт, поэтому членистоногому приходится время от времени линять — ломать старый экзоскелет и отращивать новый, более просторный. В течение краткого периода после линьки тело членистоногого мягкое, незащищённое.
Линька — основной ограничивающий фактор у членистоногих. Она может давать большие преимущества и имеет значительные недостатки. Например, форма тела у некоторых насекомых может полностью измениться после линьки: гусеница превращается в куколку, куколка — в бабочку или мотылька, чьё тело устроено принципиально иначе, чем у гусеницы.
Именно линька — причина миниатюрности. Членистоногое достигает определённых размеров и больше расти не может. В противном случае, после очередной линьки оно превратилось бы в кисель. Причиной тому является растущая сила тяжести, действующая на крупных животных. Вот почему на суше никогда не существовало насекомых крупнее гигантской стрекозы меганевры (
Meganeura) с размахом крыльев один метр или многоножки артроплевры (
Arthropleura), которая могла вырасти до трёх метров. Хотя морские членистоногие и бывают чуть крупнее наземных (их поддерживает вода), они тоже не вырастают крупнее крабоида и некоторых гигантских ракоскорпионов, обитавших в силурийском периоде и достигавших трёх метров в длину. В следующий раз, когда будете смотреть дешёвый ужастик про гигантских муравьёв или богомолов, можете посмеяться, поскольку по законам биологии такие твари существовать не могут. К сожалению, очень немногие сценаристы разбираются в зоологии настолько, чтобы знать об этом.
Бархатные черви и членистоногие
Большинство из нас никогда не видели живого бархатного червя — если, конечно, вы не из тех людей, кто живёт в тропических джунглях Южного полушария и любит по ночам ворошить кучи гнилых листьев (рис. 5.5). Тем не менее есть целый тип таких существ, называемых онихофорами. Около 180 их видов обитают в лесах Африки, Южной Америки и Юго-Восточной Азии. Большинство — крошечные по размеру (полсантиметра в длину), но есть и те, кто достигает 20 см. Они отдалённо напоминают гусениц: по нижней стороне длинного червеобразного тела находится множество коротких ножек, и на конце каждой — твёрдый крючковатый коготь, примерно как у насекомых и других членистоногих. На голове у онихофор есть ротовые придатки и усики (имеются у многих насекомых, но отсутствуют у червей). Их примитивные глаза напоминают оцеллии (простые глазки) членистоногих. Такой глаз худо-бедно даёт картинку, но онихофорам и не требуется острое зрение в тёмном влажном мире, где они обитают.
 Рис. 5.5.
Рис. 5.5. Онихофора, или бархатный червь (из коллекции IMSI Master Photo Collection)
Бархатные черви — это хищники-засадники, питающиеся мелкими насекомыми, многоножками, улитками и червями, живущими в палой листве. Как правило, они засекают жертву по изменению потоков воздуха, подползают к вероятной добыче, мягко и грациозно извиваясь, а затем несколько раз осторожно к ней прикасаются, чтобы определить, достаточно ли она мелкая и годится ли в пищу, либо это крупное существо, которое может оказаться хищником. Если это потенциальная пища, онихофоры выделяют из желёз, идущих у них вдоль тела, зловонную слизь, с помощью которой захватывают и подавляют жертву. Кроме того, благодаря слизи онихофоры неаппетитны для хищников. Напав на жертву, животное любой ценой найдёт её, если той удастся ускользнуть. Как только добыча обездвижена, бархатный червь убивает её ударом мощных челюстей, после чего дожидается, когда содержащиеся в слизи ферменты частично не переварят её внутренности, подготовив к употреблению.
У онихофор, в отличие от членистоногих, нет твёрдой хитиновой оболочки. Вместо неё у них обычная кожа, состоящая из дермы и эпидермиса, а форма тела (как у большинства червей) поддерживается благодаря давлению жидкости во внутренних полостях. Эластичная кожа позволяет забиваться в крошечные трещины, прячась от хищников.
Такая стратегия защищает бархатных червей и от высыхания, либо они могут закопаться в почву. Кожа онихофор покрыта сотнями крошечных мягких волокнистых щёточек, благодаря чему червь и кажется бархатистым на ощупь. Они настолько мелкие, что газообмен происходит в основном через кожу. Простые трахеи на коже служат дыхательными органами, но в отличие от обычных членистоногих, способных закупоривать свои трахеи, у онихофор они всегда открыты. Поэтому животные могут обитать только во влажных тропических регионах, где им не грозит высыхание.
В принципе бархатные черви непримечательны, если не считать их размножения. Многие виды онихофор вынашивают яйца внутри тела и являются живородящими. У самцов некоторых видов семя выделяется из специального органа на голове, поэтому, чтобы оплодотворить самку, они засовывают голову ей во влагалище.
Макроэволюция от типа к типу
Почему бархатные черви так интересны и важны? Это стопроцентная переходная форма от червей к членистоногим. У онихофоры — длинное мягкое тело, как у большинства червей, но при этом имеются многие более совершенные признаки членистоногих. Частичная сегментация тела, глаза и усики, напоминающие аналогичные органы членистоногих, крючковатые «коготки» на неуклюжих лапках, как у гусеницы, — вот лишь некоторые примеры анатомического сходства.
Ещё важнее, что онихофорам приходится линять, чтобы расти. Эта черта роднит их с членистоногими и несколькими группами беспозвоночных: тихоходками, круглыми червями (нематодами) и некоторыми другими. Эта характеристика настолько фундаментальна с точки зрения эмбриологии и строения тела многих животных, что убедительно доказывает их близкое родство. На самом деле, большая группа типов, включающая всех подобных животных (членистоногих, онихофор, круглых червей, тихоходок и др.), была названа линяющими (
Ecdysozoa). Мало того, в последние годы внимательно изучают ДНК и иные молекулярные системы этих животных. Удалось выяснить, что для всех линяющих характерны общие последовательности в ДНК, а также другое молекулярное сходство, указывающее на их близкое родство.
Окаменелости примитивных членистоногих, а также крошечные пластинки лобоподов встречаются в двух самых ранних ярусах кембрия — немакит-далдынском и томмотском, — сформировавшихся задолго до атдабанского, к которому относится Кембрийский взрыв. Но тогда как членистоногие уцелели? Сначала в кембрии, из которого вышли трилобиты, а затем в силуре, когда первые многоножки, скорпионы и насекомые выбрались на сушу; лобоподы вымерли в девоне. Однако незадолго до того их потомки, «бархатные черви», тоже обосновались на суше. Эти мягкотелые существа почти не фоссилизировались, но «бархатный червь» илиод (
Ilyodes), найденный в отложениях карбона, свидетельствует, что они обитали на суше уже 360 млн лет назад. С тех пор онихофоры и ведут в джунглях планеты свою неприметную жизнь.
Лучше один раз увидеть!
Сланцы Бёрджесс расположены на территории национального парка Йохо в Британской Колумбии. Добраться туда нелегко от любой дороги, поэтому они открыты лишь для профессиональных исследователей. Однако окаменелости из сланцев Бёрджесс экспонируются в некоторых музеях. В Музее естественной истории под открытым небом Чикаго можно посмотреть компьютерную анимацию, выводимую сразу на три экрана, где демонстрируется, как могла бы выглядеть фауна сланцев Бёрджесс в кембрийском океане. Там вы увидите плывущую пикайю, ползущих галлюцигению и виваксию; опабинию, пытающуюся поймать червя-приапулиду оттойю; рой марелл, а также аномалокариса, охотящегося на трилобита. Под этой анимацией — пояснительные панели и 24 окаменелости из сланцев Бёрджесс.
Другие подобные музеи в США — это, в частности, Денверский музей природы и науки, Геологический музей висконсинского университета в Мэдисоне, Музей естественной истории им. Сэма Ноубла в Оклахомском университете (Норман, штат Оклахома), Смитсоновский институт (Вашингтон, округ Колумбия). В Канаде окаменелости из сланцев Бёрджесс есть в коллекциях Канадского музея природы (Оттава, провинция Онтарио), Королевского музея Онтарио (город Торонто), Королевского Тирреловского палеонтологического музея в Драмхеллере (провинция Альберта). В Европе такие окаменелости можно увидеть в Седвижском музее естественных наук Кембриджского университета (Кембридж, Англия) и в Музее естествознания (Вена, Австрия).
Дополнительная литература
Conway Morris, Simon. The Crucible of Creation: The Burgess Shale and the Rise of Animals. — Oxford: Oxford University Press, 1998.
Erwin, Douglas H., and James W. Valentine. The Cambrian Explosion: The Construction of Animal Biodiversity. — Greenwood Village, Colo.: Roberts, 2013.
Foster, John H. Cambrian Ocean World: Ancient Sea Life of North America. — Bloomington: Indiana University Press, 2014.
Gould, Stephen Jay. Wonderful Life: The Burgess Shale and the Nature of History. — New York: Norton, 1989.
06. Это червь или моллюск?
Происхождение моллюсков
Пилина
Если устроить состязание между беспозвоночными по размерам, скорости и интеллекту, то большинство золотых и серебряных медалей достались бы кальмарам и осьминогам. Но тип моллюсков стал вторым по величине в царстве животных (более 100 000 описанных видов) благодаря не этим эффектным призёрам. Достижением моллюски обязаны в первую очередь медлительным и неповоротливым улиткам, с долей участия ещё более медленных мидий и устриц. Слово mollusca означает «мягкотелые». Действительно, своим нежным и сочным мясом моллюски пришлись людям по вкусу, как никакие другие беспозвоночные. Но многие моллюски более известны своими прочными раковинами, материал для которых эти медлительные и уязвимые животные постепенно выделяют в качестве защиты от плотоядных врагов. По иронии судьбы именно за красоту и ценность этих раковин моллюски подверглись со стороны человека хищническому истреблению, а некоторые даже оказались на грани исчезновения.
Ральф и Милдред Буксбаум. «Animals without Backbones»
Недостающие звенья найдены
Ископаемая летопись полна удивительных промежуточных форм, демонстрирующих, например, эволюцию лошади, начиная от мелких четырёхпалых предков, либо эволюцию млекопитающих от немлекопитающих (см. главы 19 и 22). Но многим недостаточно этой горы доказательств, люди спрашивают: как все современные типы животных (моллюски, черви, членистоногие, иглокожие и т. д.) могли произойти от общего предка? Где доказательства столь кардинальных изменений строения тела или макроэволюции?
Очень долго отсутствовали ископаемые свидетельства того, как всё происходило, за исключением того, что ярко выраженные анатомические черты разных существ свидетельствовали: они действительно произошли от общего предка. Например, связь между членистоногими и бархатными червями выявили по сходству живых экземпляров задолго до того, как ископаемая летопись подтвердила такие изменения, а впоследствии их близкое родство подкрепили новейшие факты (см. главу 5).
Или возьмём другой пример: моллюски. Сегодня данный тип включает свыше 100 тыс. описанных видов — больше чем любой другой тип, кроме членистоногих. Среди моллюсков попадаются такие медленные примитивные существа, как хитоны и блюдечки, сидящие на камнях в приливных заводях или медленно ползающие, поедая водоросли; и безголовые мидии с устрицами, неподвижные и питающиеся, фильтруя воду через жабры; и кальмары с осьминогами — стремительные и умные, способные общаться при помощи эффектных узоров на шкуре, а также решать довольно сложные задачи. Подобно членистоногим, моллюски завоевали большинство экологических ниш на Земле: они живут в толще воды вместе с планктоном, на морском дне и на суше (таковы, например, сухопутные улитки и слизни). Большинство моллюсков мелкие, но среди них есть настоящие великаны — скажем, гигантский кальмар, достигающий 18 м в длину; двустворчатый моллюск тридакна, чья раковина может иметь более метра в поперечнике, а также колоссальная морская улитка
Campanile giganteum с огромной спиралевидной раковиной более метра.
Как выглядел общий предок всех этих улиток, брюхоногих и кальмаров? У какого животного есть базовые «кирпичики» для эволюции организмов подобных форм? Откуда произошли моллюски и каково их место среди остальных типов?
Большинство специалистов по моллюскам полагают, что форма тела у общего предка всех моллюсков должна базироваться на элементах, которые присутствуют в строении всех представителей данного типа (рис. 6.1). Зачастую такое существо называют «гипотетическим прамоллюском», исходя из его простого строения и набора черт, характерного для многих моллюсков. Вероятно, оно могло быть покрыто мясистой прослойкой, мантией, которая обрастала секретируемым веществом, формировавшим простую раковину в форме колпачка — сегодня ими обладают блюдечки, одни из самых примитивных моллюсков. На брюхе такое существо должно было иметь широкую толстую ногу, которая позволяла бы плотно присасываться к камням для защиты и медленно отползать в безопасное место, где животное могло бы кормиться.
 Рис. 6.1.
Рис. 6.1. Развитие моллюсков от гипотетического первопредка (иллюстрация создана по материалам
Euan N. K. Clarkson. Invertebrate Palaeontology and Evolution, 4th ed. — Oxford: Blackwell, 1993 и
Donald R. Prothero. Bringing Fossils to Life: An Introduction to Paleobiology, 3rd ed. — New York: Columbia University Press, 2013. — Fig. 16.3)
У всех ныне живущих моллюсков есть пищеварительный тракт, идущий от рта до ануса, а также дыхательная система с жабрами, напоминающими пёрышки. Жабры извлекают кислород из морской воды и выделяют углекислый газ. Они находятся в специальном «кармане» мантии, так называемой мантийной полости. Прамоллюск должен был обладать всеми этими признаками, а также выделительной и репродуктивной системой. Итак, первые моллюски были очень похожи на блюдечки: простая колпачковидная раковина, формируемая из секрета мантии; широкая нога для ползания и прикрепления к камням; однонаправленный пищеварительный тракт от рта до ануса; дыхательная система, а также большинство других систем, имеющихся сегодня у основных групп моллюсков: выделительная, репродуктивная и т. д.
Первые моллюски
Морские биологи располагают всеми возможностями для изучения живых моллюсков. За их поведением можно наблюдать в океанариумах и природе. Все мягкие ткани моллюсков можно препарировать и рассматривать. Специалисты по молекулярной генетике могут секвенировать ДНК моллюсков по самому крошечному образцу тканей и узнать, какие животные состоят с ними в наиболее близком родстве. Все эти исследования однозначно свидетельствуют: ближайшими ныне живущими родственниками моллюсков являются кольчатые черви — например, всем известные дождевые и многощетинковые (полихеты), в изобилии обитающие практически во всех морских экосистемах. Тем не менее остаётся огромный пробел: как существо, напоминающее дождевого червя, могло эволюционировать в блюдечко — организм с несегментированным телом и твёрдой раковиной?
Проблему осложняет тот факт, что большинство червей не оставляют окаменелостей — не считая ходов в грунте, которые мало что могут поведать о том, кто их оставил. С другой стороны, моллюски, как правило, имеют всего один жёсткий орган — раковину, которая даёт на порядок меньше информации, чем мягкие ткани. Но палеонтологи изрядно напрактиковались в работе с простыми раковинами древних моллюсков и в поиске всевозможных подсказок, остающихся на месте мягких тканей.
Ещё в 1880-е годы палеонтологи приступили к описанию простых моллюсков с раковинами-колпачками, живших в раннем палеозое (рис. 6.2). Эти окаменелости сохранялись плохо, поэтому о них было трудно сказать что-то более определённое, чем просто отметить сходство с раковинами блюдечек. Соответственно, их обладатели должны были вести примерно такой же образ жизни, как блюдечки. В 1880 году шведский палеонтолог Густав Линдстрём описал ископаемую силурийскую раковину, найденную на острове Готланд, которую назвал
Tribidium unguis (видовое имя происходит от латинского слова, означающего «копыто» или «ноготь», поскольку внешне раковина напоминала ноготь). К 1925 году эту окаменелость переименовали в
Pilina unguis. Никто из палеонтологов того времени не мог сказать о находке что-то более определённое, чем то, что «она очень похожа на блюдечко». Соответственно, её считали очень примитивным блюдечком. Однако на внутренней стороне хорошо сохранившихся раковин шли два ряда рубцов, и это значило, что у моллюска были парные мускулы. Однако без мягких тканей дальнейшие исследования окаменелости были затруднительны.
 Рис. 6.2.
Рис. 6.2. Окаменелость простого колпачковидного моллюска Pilina, напоминающего современное морское блюдечко (из фонда Wikimedia Commons)
Со временем множество окаменелостей этих простых существ с округлыми раковинами накопилось в пластах от кембрия до девона. Некоторые палеонтологи полагали, что эти ископаемые могут относиться к древнейшим, самым примитивным моллюскам, но образцы по-прежнему оставались слишком фрагментарными, чтобы судить о них с уверенностью. Позже были найдены колпачковидные, створчатые и спиральные окаменелости, относящиеся к мелкой раковистой фауне (см. главу 3), позволяющие предположить, что в раннем кембрии жили предки моллюсков (см. рис. 3.2). Но в распоряжении палеонтологов были лишь контуры раковин и чёткие отпечатки некоторых органов существ — на основании этого и оставалось теоретизировать.
Галатея и переворот в биологии
В конце 1940-х годов начался период бурного развития океанографии и морской биологии. Сражения между подводными лодками в годы Второй мировой войны поведали нам о мире, ранее остававшемся почти неизученным, и о более чем 70% земной поверхности, покрытой океанами. Вскоре после окончания войны правительства многих государств (особенно США, Великобритании и Дании) начали финансировать крупные научные экспедиции, занимавшиеся картографированием морского дна и разведкой того, что там находится, а также сбором образцов подводных минералов и морской фауны со всего мира. Для решения задачи переоборудовали грозные субмарины минувшей войны. На борту они несли протонные магнетометры, первоначально разработанные для поиска подводных лодок, а теперь позволившие получить ключевые доказательства в пользу спрединга морского дна и тектоники плит. Экспедиции методично брали осадочные керны почти во всех районах морского дна; с помощью эхолота замеряли глубину взятия проб; сбрасывали с кормы лодки динамитные шашки, чтобы определить структуру осадочного слоя на морском дне по расходящимся от места взрыва звуковым волнам.
Среди других первопроходческих послевоенных операций была и вторая экспедиция «Галатея», проведённая датчанами с 1950 по 1952 год. Корабль назвали в честь героини древнегреческого мифа о Пигмалионе и Галатее. По сюжету мифа скульптор Пигмалион изваял из мрамора прекрасную женщину, которую назвал Галатеей, и влюбился в неё. Он был настолько очарован своим творением, что боги оживили Галатею, вняв его молитвам. Некоторым здесь вспомнится сюжет бродвейского мюзикла «Моя прекрасная леди», в котором профессор Генри Хиггинс (Пигмалион) преображает бедную девушку из трущоб Элизу Дулиттл (Галатею) в изысканную аристократку. Мюзикл, в свою очередь, поставлен на основе знаменитой пьесы Джорджа Бернарда Шоу «Пигмалион», где обыгран сюжет древнегреческого мифа.
Первая экспедиция «Галатея» состоялась в 1845–1847 годах. Её выполнил трёхмачтовый парусник, перед экипажем которого поставили задачу исследовать воды, омывающие крупнейшие датские колонии по всему миру. В 1941 году журналист Хакон Мильхе и океанограф-ихтиолог Антон Фредерик Брунн добивались финансирования для другой экспедиции, преследовавшей научные и коммерческие цели. Однако Вторая мировая война и вторжение в Данию нацистов вынудили отложить экспедицию.
В июне 1945 года, сразу после окончания войны, датское научное сообщество возобновило привлечение значительных средств и начало строить большие планы. Учёные приобрели снятый со службы британский сторожевой корабль «Лит». Это было судно с долгой и славной историей сопровождения других кораблей через Атлантику в годы войны, топившее подлодки. Датчане переоснастили его для океанографических целей и переименовали в «Галатея-2». В отличие от первой «Галатеи», этот корабль был рассчитан на глубоководные исследования, вычерпывание осадка с океанического дна и промер максимальных глубин. Корабль побывал в некоторых местах, которые посещала и экспедиция середины XIX века, но главным достижением второй экспедиции стало драгирование вод на глубине 10 190 метров в Филиппинском жёлобе (на тот момент это были самые глубоководные из всех добытых образцов), а также во многих других глубинных районах океана. Так удалось добыть существ, ранее не попадавшихся учёным.
Вместе со многими эффектными и причудливыми глубоководными рыбами, другими морскими созданиями тогда был открыт занятный моллюск, добытый в 1952 году на глубине более 6000 метров в Центральноамериканском жёлобе (рис. 6.3). Когда зоолог Хенниг Лемхе в 1957 году опубликовал работу об этом организме, он понимал, что это поистине революционная находка, и назвал моллюска
Neopilina galatheae — в честь окаменелости пилина и корабля, позволившего сделать открытие. Действительно, моллюск был родственником таинственных колпачковидных окаменелостей из раннего палеозоя, и по его мягким тканям палеонтологи смогли интерпретировать таинственные рубцы и отметины на ископаемых образцах. Знаменитый зоолог Энрико Швабе назвал это животное «величайшей сенсацией XX века».


 Рис. 6.3.
Рис. 6.3. «Живое ископаемое» неопилина, реликт раннего кембрия, переходная форма между кольчатыми червями и моллюсками. (
А) сегментированные парные жабры по обе стороны от ноги в центральной части тела. Также имеются парные сегментированные сократительные мускулы и другие системы органов; (
Б–
В) ныне живущие неопилины (иллюстрации предоставлены J. B. Burch, Мичиганский университет)
Лемхе указывал, что неопилина — настоящее «живое ископаемое», реликтовый род из класса моллюсков моноплакофор (
Monoplacophora; в переводе с греческого — несущий одну раковину), которые исчезли из палеонтологической летописи в девоне. А какую удивительную информацию принесло изучение этого образца! Именно там, где на окаменелостях находились два ряда рубцов, неопилина имела парные мускулы, от которых и оставались рубцы. Это свидетельствовало, что мускулы у такого моллюска сегментированы, как у кольчатых червей. Оказалось, что у неопилины сегментированы не только мускулы, но и жабры, почки, несколько сердец, парные нервные цепи и гонады. В общем, неопилина демонстрирует, что таинственные окаменелости-моноплакофоры принадлежали полумоллюскам-получервям: эти существа имели сегментированное тело и все системы органов, в точности как у их червеподобных предков, но также обладали мантией, раковиной, широкой ногой и другими чертами, присущими примитивным раковинным моллюскам — например, хитонам и блюдечкам.
С тех пор как неопилину описали в 1957 году, было найдено много живых и ископаемых моноплакофор. В настоящее время известно 23 ныне живущих вида этого класса. Такие «живые ископаемые» встречаются в основном на глубинах от 6000 до 6500 м, но некоторые обитают и в гораздо более мелких водах — всего на 175-метровой глубине.
Об их среде обитания известно мало, поскольку они живут глубоко и погибают вскоре после того, как их отлавливают и извлекают на поверхность, где давление и температуры сильно отличаются от условий глубокого океана. Предполагается, что моноплакофоры питаются на илистом грунте, выцеживая питательные вещества из донных отложений и захватывая опускающийся на эти глубины планктон. Это характерно для большинства организмов, живущих там, где очень темно и не может происходить фотосинтез.
Как такая важная группа животных могла долго ускользать от внимания учёных? Основная причина в том, что у нас почти не было средств для сбора и изучения организмов из глубочайших частей океана. Вторая экспедиция «Галатея» стала одной из первых, взявшихся за решение данной задачи. На самом деле ныне живущая моноплакофора
Veleropilina zografi была открыта ещё в 1896 году, но этого моллюска ошибочно причислили к обычным блюдечкам и забыли. Учёные повторно исследовали её только в 1983 году и осознали, что их коллеги-предшественники видели живую моноплакофору задолго до того, как была открыта неопилина.
Открытие неопилины является одним из классических примеров того, как загадочная группа ископаемых животных, долго считавшихся исчезнувшими, в целости и сохранности повторно обнаруживалась в глубинах океана. Ещё важнее, что описание многих ныне живущих и вымерших моноплакофор показывает, как моллюски эволюционировали от общего предка, роднящего их с кольчатыми червями, а затем утратили сегментацию и разделились на одностворчатых, двустворчатых, головоногих и другие классы этого важного типа. Соответственно, палеонтологическая летопись подкрепляет результаты исследований в области анатомии и молекулярной биологии: моллюски произошли от кольчатых червей, а представители класса моноплакофор — переходные формы, демонстрирующие макроэволюцию от типа к типу.
Дополнительная литература
Ghiselin, Michael T. The Origin of Molluscs in the Light of Molecular Evidence //
Oxford Surveys in Evolutionary Biology. — 1988. — Vol. 5. — 66–95.
Giribet, Gonzalo, Akiko Okusu, Annie R. Lindgren, Stephanie W. Huff, Michael Schrödl, and Michele K. Nishiguchi. Evidence for a Clade Composed of Molluscs with Serially Repeated Structures: Monoplacophorans Are Related to Chitons //
Proceedings of the National Academy of Sciences. — 2006. — Vol. 103. — 7723–7728.
Morton, John Edward. Molluscs. — London: Hutchinson, 1965.
Passamaneck, Yale J., Christoffer Schander, and Kenneth M. Halanych. Investigation of Molluscan Phylogeny Using Large-subunit and Small-subunit Nuclear rRNA Sequences //
Molecular Phylogenetics and Evolution. — 2004. — Vol. 32. — 25–38.
Pojeta, John, Jr. Molluscan Phylogeny //
Tulane Studies in Geology and Paleontology. — 1980. — Vol. 16. — 55–80.
Runnegar, Bruce. Early Evolution of the Mollusca: The Fossil Record //
John D. Taylor, ed. Origin and Evolutionary Radiation of the Mollusca. — Oxford: Oxford University Press, 1996. — P. 77–87.
Runnegar, Bruce, and Peter A. Jell. Australian Middle Cambrian Molluscs and Their Bearing on Early Molluscan Evolution //
Alcheringa. — 1976. — Vol. 1. — 109–138.
Runnegar, Bruce, and John Pojeta Jr. Molluscan Phylogeny: The Paleontological Viewpoint //
Science. — 1974. — October 25. — P. 311–317.
Salvini-Plawen, Luitfried V. Origin, Phylogeny, and Classification of the Phylum Mollusca //
Iberus. — 1991. — Vol. 9. — 1–33.
Sigwart, Julia D., and Mark D. Sutton. Deep Molluscan Phylogeny: Synthesis of Palaeontological and Neontological Data //
Proceedings of the Royal Society. — 2007. — Vol. В 247. — 2413–2419.
Yonge, C. M., and T. E. Thompson. Living Marine Molluscs. — London: Collins, 1976.
07. Растущие из моря
Происхождение наземных растений
Куксония
Наиболее убедительное доказательство эволюции флоры — растительная палеонтологическая летопись. Глубоко в земной коре остались свидетельства последовательного развития и изменений, которые за миллионы лет претерпели разные группы царства растений. Каждый год палеоботаники извлекают новые ископаемые образцы, помогающие добавить ещё несколько элементов в картину, которая, как мы надеемся, однажды сложится в непрерывную историю развития царства растений — от эпохи, отстоящей от нас более чем на миллиард лет, до сегодняшнего дня. За этот долгий период в растительном мире произошли коренные изменения. Целые классы возникали, переживали расцвет и исчезали; без палеонтологической летописи современные ботаники даже не подозревали бы, что такие группы растений когда-то существовали.
Теодор Делеворьяз. «The Little Things that Run the World»
Мы любуемся изумительными лесами и саваннами Земли и прославляем «зелёную планету», которая изобилует растительностью, обеспечивающей существование самых разнообразных животных. Но так было не всегда. На протяжении большей части 4,5 млрд лет в истории Земли наша планета была пустынным и негостеприимным местом. Не существовало наземных растений, которые могли бы выжить на её бесплодной поверхности. Поэтому голые породы подвергались интенсивной химической эрозии, все их питательные вещества утекали в океан, причём морские организмы не могли их потреблять. Первые 1,5 млрд лет в истории жизни на Земле единственными фотосинтезирующими организмами были сине-зелёные бактерии (цианобактерии), обитавшие на океаническом мелководье и формировавшие строматолиты (см. главу 1). Затем, около 1,8 млрд лет назад, появились первые признаки существования водорослей — настоящих растений с эукариотическими клетками (имеют чётко выраженное ядро для хранения ДНК, а также органеллы — например, хлоропласты, в которых протекает фотосинтез). Цианобактерии и водоросли продолжили образовывать на морском мелководье огромные илистые подушки.
Экстремальные перепады температур, сильнейшие ливни и стоки, не сдерживаемые растительным покровом, а также отсутствие озонового слоя (в атмосфере не было свободного кислорода) — по этим причинам лишь немногие растения могли переселиться из воды на сушу. Животные и растительные клетки подвергались жёсткому ультрафиолетовому излучению, вызывавшему генетические мутации и в конечном итоге губившему клетки. Пока не было озонового слоя, только защитный водяной покров оберегал большинство организмов от ультрафиолета.
По данным химии, около 1,2 млрд лет назад первые организмы начали осваивать сушу. Вероятно, это были очень простые сообщества водорослей и грибов, так называемые криптогамные корочки, сильно напоминающие органический налёт, образующийся на пустынном грунте, который долго остаётся непотревоженным. Здесь можно провести параллели с лишайниками, разрушающимися в сухих руслах, поскольку лишайник — это не отдельный организм, а симбиоз водорослей и грибов. Вероятно, криптогамные корочки были единственной жизнью на суше и могли содействовать укреплению почвы, её стабилизации против выветривания и вымывания. В то же время они помогали морским водорослям и цианобактериям закачивать в атмосферу больше кислорода.
Естественно, при отсутствии достаточного количества растительной пищи на суше не было и животных. Последние нуждались не только в пище, но и в значительном количестве свободного кислорода в воздухе, чтобы дышать. Очевидно, такое количество кислорода накопилось в атмосфере не ранее 530 миллионов лет назад. Сочетание экстремальных перепадов температур, отсутствие укрытий и пищи, а также бесконтрольная эрозия — всё это превращало сушу в довольно враждебную среду обитания, непригодную для заселения большинством живых существ.
Первые наземные растения
Итак, зелёная планета, которую мы воспринимаем как данность, существует относительно недавно. Чтобы приступить к завоеванию суши, растения должны были стать более совершенными, чем карликовые водоросли, образующие подводные маты. Водоросли растут, пока остаются в воде, но на суше они могут оставаться, лишь будучи влажными — иначе гибнут.
Кроме того, размножаться водоросли могут только в воде: мужские гаметы просто плывут к женским. Например, у зелёных водорослей и многих других примитивных растений чередуются поколения, появившиеся в ходе полового размножения (с участием гаплоидных сперматозоидов и яйцеклеток) и бесполого (размножающиеся вегетативно) (рис. 7.1). Диплоидное растение, обладающее как женским, так и мужским набором хромосом, называется
спорофит. В спорофитах протекает мейоз: в
спорангии образуются споры и происходит половое размножение. Гаплоидное растение (всего с одним набором хромосом, претерпевшее мейоз) называется
гаметофит. В нём в специальных органах образуются отдельные сперматозоиды, яйцеклетки либо оба вида гамет. Чередование поколений — обычный репродуктивный механизм, свойственный многим группам примитивных растений и животных, в том числе большинству кораллов, морских анемонов и медуз, а также группе крошечных морских раковинных амёб, называемых фораминиферами. Спорофиты у примитивной наземной флоры (например, у папоротников) — это видимая часть растения. Спорофит извергает летучие гаплоидные споры, возникающие в процессе мейоза. Споры могут приземлиться во влажном месте и прорасти, образовав крошечное (менее сантиметра в высоту) растение-гаметофит. Сперматозоиды и яйцеклетки хранятся в гаметофите отдельно друг от друга, поэтому сперматозоиды могут подплыть к яйцеклеткам только во влажной среде. Это ограничивает возможности большинства примитивных наземных растений; в процессе размножения данное «слабое звено» не позволило им заселить более засушливые экосистемы.
 Рис. 7.1.
Рис. 7.1. Упрощённый жизненный цикл бессемянного сосудистого растения. Зрелый спорофит производит споры, из которых вырастает гаметофит, а гаметофит, в свою очередь, производит сперматозоиды и яйцеклетки, из которых путём полового размножения появляется новый спорофит (по
Donald R. Prothero and Robert H. Dott Jr. Evolution of the Earth, 6th ed. — New York: McGraw-Hill, 2001)
Возможность высыхания — ещё одна проблема, с которой сталкиваются наземные растения. Если поверхность растения не погружена в воду, она иссыхает как выброшенные на берег водоросли, если растение не защищено особым воскоподобным покровом, или
кутикулой, для удержания воды. Но кутикула тоже мешает водообмену на поверхности растения: так ему сложнее поглощать углекислый газ, выделять кислород и регулировать транспирацию водяного пара. Крошечные поры, называемые устьицами, — это отверстия в кутикуле. Они могут открываться и закрываться, регулируя водообмен и газообмен. Однако в процессе открытия устьиц вода теряется.
Итак, что гласит палеонтологическая летопись о проникновении растений на сушу? Первые ископаемые следы — это споры мхов и печёночников, двух низкорослых растений, сегодня встречающихся в большинстве экосистем (рис. 7.2). По возрасту эти споры относятся к ордовику (им около 450 млн лет), но некоторые из них, возможно, относятся ещё к среднему кембрию (около 520 млн лет назад). Сегодня существует около 900 родов и 25 тыс. видов этих примитивнейших наземных растений. Они заняли на суше практически все экологические ниши, даже холодное и влажное побережья Антарктики, но не живут в солёной воде. Эти растения претерпели множество адаптаций, которые помогли им выжить на суше — в частности, способность останавливать метаболизм в неблагоприятных условиях, например при засухе или экстремальных температурах; тенденция расти купами; способность прорастать фрагментами, превращающимися в новые растения, и заселять пустынные участки голого камня, где почти нет почвы, либо расти на поверхности других организмов — например, деревьев.
 Рис. 7.2.
Рис. 7.2. Четырёхчастные споры времён позднего ордовика, найденные в Ливии, — древнейшие следы наземных растений. Увеличение в 1500 раз (фотография предоставлена Jane Gray)
Высокие первопроходцы: сосудистые растения
Чтобы растения могли жить на суше и достигать большой высоты, им требуются сложные системы органов для доставки жидкости вверх, против действия силы тяжести, для обеспечения дыхания, удаления отходов и поддержки всего растения. Морские водоросли — например ламинария — могут иметь многометровые нити, но поскольку они постоянно находятся в воде, им не требуется передавать жидкость из конца в конец.
Растения, обладающие органами для такой передачи, называют
сосудистыми, поскольку они располагают сетью трубок, чтобы передавать жидкости и питательные вещества из одной части растения в другую — подобно тому, как наша сердечно-сосудистая система переносит жидкость (кровь) ко всем частям тела, обеспечивая их питательными веществами и утилизируя продукты обмена веществ. Однако сосудистые растения будто «распяты»: вода и питательные вещества поступают снизу, из почвы, а солнечный свет, необходимый для фотосинтеза, льётся сверху. Корень растения высасывает из почвы воду и питательные вещества, и растение переносит их к листьям, где протекает фотосинтез (растение поглощает углекислый газ и выделяет кислород). При этом теряется некоторое количество воды.
Когда растения начали расти из воды, они столкнулись с двумя проблемами. Во-первых, воду и питательные вещества нужно транспортировать в верхнюю часть растения. Во-вторых, растение должно тянуться вверх и противостоять силе гравитации, направляющей его вниз. Решение заключалось в эволюции продолговатых проводящих клеток,
трахеид, выстланных
лигнином — побочным продуктом метаболизма с участием воды. Такая проводящая ткань пролегает внутри стебля как непрерывная центральная нить. У более совершенных растений трахеиды могут накапливаться слоями, образуя крупные древесные стволы. Такие сосудистые растения называются трахеофитами, поскольку внутри у них находятся трахеиды.
Открытие Изабель Куксон
Древнейшие ископаемые трахеофиты — крошечные, при этом они плохо сохранились, поскольку растения состояли из мягких органических веществ и не имели древесных тканей, повышающих вероятность сохранения. Такие растения ордовикского периода пока не известны, но в силурийском периоде (433–393 млн лет назад) существовали простые растения, получившие имя «куксония» (
Cooksonia) (рис. 7.3,
А–
В). Палеоботаник Уильям Генри Лэнг в 1937 году назвал их так в честь неутомимой собирательницы Изабель Куксон, обнаружившей первые образцы этих ископаемых в карьере Пертон на территории Уэльса.


 Рис. 7.3.
Рис. 7.3. Куксония. (
А–
Б) окаменелости; (
В) реконструкция внешнего вида живого экземпляра, на которой показаны устьица и спорангии ((
А–
Б), иллюстрации предоставлены Hans Steuer; (
В) илл. предоставлена Nobumichi Tamura)
Куксония устроена максимально просто для сосудистого растения: большинство образцов просто расплющены и имеют только примитивный стебель (обычно менее 3 мм в диаметре), который разделяется на две веточки. Большая часть растений имела не более 10 см в длину. Многие из этих ветвистых стеблей оканчивались своеобразными шариками (так они выглядели на первых расплющенных окаменелостях), где, как предполагалось, могли образовываться споры, — спорангиями. Однако после изучения лучших из сохранившихся образцов и по итогам более детальной работы недавно удалось выяснить, что спорангии были не округлыми, а по форме напоминали воронки или раструбы с конической полостью сверху, прикрытой «веком», которое распадалось, выпуская наружу споры (рис. 7.3,
В).
У куксонии не было листьев. Вероятно, она фотосинтезировала всей поверхностью. У неё определённо не имелось более сложных органов — цветов или семян. По-видимому, куксония не обладала и отдельными корнями, а разрасталась при помощи коротких горизонтальных соединительных стебельков, или
ризом, как многие современные растения с подземными побегами. Итак, куксония могла размножаться вегетативно. Вокруг уплощённых, плохо сохранившихся образцов видны тёмные области, которые могут быть следами сосудистой ткани, хотя они слишком нечёткие, чтобы судить с уверенностью. Кроме того, как минимум, у некоторых образцов, видимо, были устьица, дополнительно подтверждающие, что куксония фотосинтезировала всей поверхностью, тогда как более совершенные растения фотосинтезируют с помощью специальных органов — например, листьев или иголок.
С растениями, называемыми куксонией, связано минимум четыре типа спор, поэтому большинство палеоботаников считают этот род таксономической «мусорной корзиной», включающей несколько эволюционных линий очень примитивных растений. Но эти образцы и их детали сохранились слишком плохо, чтобы куксонию можно было уверенно разделить на несколько родов, как того требует таксономия. Однажды её всё-таки классифицируют по родам — так рано или поздно происходило с большинством таксономических «мусорных корзин».
Планета начинает зеленеть
Почти никого, кроме палеоботаников, такое неказистое и маленькое растение не воодушевляет. Но куксония и начало всех сосудистых растений — великий эволюционный и экологический прорыв. Существование сосудистых растений на суше и «зелёных зон» открыло множество других возможностей, особенно для животных. В позднем ордовике начинают попадаться образцы грунта с прорытыми ходами, которые, вероятно, проделали многоножки. Скорее всего, это и были первые сухопутные животные. Затем, начиная с силура, обнаруживаются окаменелости других членистоногих, в том числе скорпионов, пауков, губоногих и первых бескрылых насекомых. Суша уже не была бесплодной, на ней развивалась сложная пищевая сеть растительноядных членистоногих и разнообразных хищников, пожиравших как травоядных, так и друг друга. Наконец, спустя около 100 млн лет после того как членистоногие заселили сушу, из воды выбрались первые амфибии (см. главу 10). Отныне суша никогда не оставалась совершенно голой, всегда была покрыта зелёным ковром растений.
Прослеживая поздний силур, мы наблюдаем ещё большее разнообразие простых сосудистых растений. Затем, в девоне биоразнообразие растений увеличивается взрывным образом, а к концу девона появляются первые леса. Наряду с мхами и печёночниками развивались и более сложные растения, например папоротники. Кроме того, в позднем силуре или в девоне возникли ещё две важные группы ныне живущих растений. Во-первых, плауновидные, или ликоподии, стелющиеся по земле. Эти «живые ископаемые» сегодня невысокие и неброские, но их предки в позднем палеозое росли огромными лесами, состоявшими в основном из плаунов высотой более 36 м. Это были крупнейшие наземные растения, которые видела наша планета к тому времени (рис. 7.4,
А–
Б).

 Рис. 7.4.
Рис. 7.4. Плауновидные: (
А) современные плауны, или ликоподии, представляющие собой в основном низкорослые небольшие растения; (
Б) реконструкция 50-метрового лепидодендрона — древовидного плауна, произраставшего в болотах карбона (детали ствола, коры, листьев, шишек, спор и семян реконструированы по разным находкам) (иллюстрации предоставлены Bruce Tiffney)
Другая важная группа включала хвощи, или членистостебельные (рис. 7.5,
А–
Б). Сегодня эти примитивные растения (один род, «живое ископаемое»
Equisetum) в изобилии растут на песчаных и каменистых почвах поблизости от воды. В их волокнистых стеблях содержатся крошечные кремнистые частицы, напоминающие наждак, поэтому животным трудно их поедать. Хвощи оставляют очень характерные отпечатки, поскольку каждый сегмент полого стебля по всей длине покрыт серией желобков или бороздок, а от соседних сегментов его отделяют ярко выраженные коленца, из которых растут листья. Каждый хвощёвой стебель ветвится из ризомы, которая размножается вегетативно. Хвощ — одно и самых неприхотливых растений, стремительно занимающее любую подходящую среду. Он быстро оккупирует влажные места целого сада, если его не держать под контролем, а подземный стебель хвоща почти невозможно полностью выкорчевать. Возвращается, что ни делай. Среди вымерших хвощей карбона были и такие, что достигали в высоту более 20 м (рис. 7.5,
Б).

 Рис. 7.5.
Рис. 7.5. Членистостебельные: (
А) гигантский хвощ
Calamites из карбона, достигавший 20 м в высоту; (
Б) современный хвощ
Equisetum. Листья лучеобразно расходятся из коленцев на стебле (иллюстрации предоставлены Bruce Tiffney)
Наряду с примитивными спорофитами в позднем девоне появились первые растения, размножавшиеся семенами. Семена имеют твёрдую оболочку, что позволяет им прорастать и без погружения в воду. Некоторые из исчезнувших «семенных папоротников» (это не истинные папоротники, а похожие на них растения, имевшие настоящие семена) стали первыми большими деревьями высотой до 12 м.
На смену девонским лесам из семенных папоротников в карбоне (360–303 млн лет назад) пришёл настоящий взрыв биоразнообразия папоротников, мхов, плаунов, хвощей и семенных папоротников. Угольные болота карбона, в которых они росли, породили буйную растительность на больших территориях в тропиках Северной Америки, Европы и Азии. Когда эти растения отмирали и погружались в болотную жижу, они не разлагались без остатка так быстро, как в современных болотах. Тогда почти не было животных вроде термитов, в ходе эволюции приспособившихся перерабатывать жёсткую клетчатку древесного лигнина, из которой состояли деревья. Поэтому стволы накапливались слой за слоем, подвергались давлению и высоким температурам, в результате чего превращались в уголь.
Когда такие колоссальные объёмы органики оказались связанными в виде угля в земной коре, изменились атмосфера и климат Земли. По мере накопления угля атмосфера освобождалась от углекислого газа, который запечатывался в земной коре. Вскоре «парниковый» климат раннего карбона (на полюсах не было льда, атмосфера насыщена углекислым газом, а уровень океана высок, поэтому большая часть континентальной территории находилась под водой) превратился в «ледниковый» климат позднего карбона (на Южном полюсе появилась ледяная шапка, понизилось содержание углекислого газа в атмосфере и значительно отступили океаны, когда часть воды в виде льда оказалась заперта в полярных регионах). «Ледяной» климат царил на Земле почти 200 млн лет, вплоть до середины юрского периода (эпоха расцвета динозавров), когда вновь наступило глобальное потепление, обусловленное коренными изменениями в мантии и океанических бассейнах.
Циклы от «парникового» до «ледникового» климата и обратно за миллиарды лет истории нашей планеты повторялись несколько раз. На самом деле именно благодаря существованию животных и растений Земля обитаема, а не превратилась в пекло вроде Венеры или в ледяную пустыню, как Марс. Земные организмы формируют резервуары углерода в виде известняков (это делают в основном животные) и угля (растения). Так, углекислый газ запечатывается в земной коре. Получается термостат, оберегающий планету от неконтролируемого повышения или понижения температуры.
К сожалению, мы случайно стали переделывать планету, нарушая этот естественный цикл. С тех пор как началась промышленная революция, мы сожгли миллионы тонн угля и выпустили в атмосферу диоксид углерода, который давно её покинул. Теперь диоксид углерода вышел из-под контроля, разгоняя наш антропогенный «суперпарниковый» эффект со скоростью, беспрецедентной даже по геологическим меркам. Сами того не зная, мы нарушили тонкий баланс углерода в атмосфере, океанах и земной коре. На нашей планете уже происходят климатические катаклизмы, обусловленные этими изменениями, а нашим детям и внукам придётся расплачиваться за эти опасные эксперименты, в ходе которых мы испортили глобальный термостат.
Лучше один раз увидеть!
В очень немногих музеях экспонируются образцы древнейших растений. В Музее естественной истории под открытым небом в Чикаго можно увидеть ринию (
Rhynia) — близкую родственницу куксонии, а также великолепные окаменелости и диорамы с изображением лесов, покрывавших угольные болота. В Денверском музее природы и науки есть выставка примитивных растений, а также диорама угольного болота (такую же диораму вы найдёте в Смитсоновском институте (Вашингтон, округ Колумбия)).
Древнейший лес на Земле рос в девонском периоде (380 млн лет назад) близ современного города Гилбоа в штате Нью-Йорк. Окаменелости разных частей деревьев представлены в музее Гилбоа (http://www.gilboafossils.org/), в ратуше Гилбоа и в Музее штата Нью-Йорк (город Олбани).
Дополнительная литература
Gensel, Patricia G., and Henry N. Andrews. The Evolution of Early Land Plants //
American Scientist. — 1987. — Vol. 75. — 468–477.
Gray, Jane, and Arthur J. Boucot. Early Vascular Land Plants: Proof and Conjecture //
Lethaia. — 1977. — Vol. 10. — 145–174.
Niklas, Karl J. The Evolutionary Biology of Plants. — Chicago: University of Chicago Press, 1997.
Stewart, Wilson N., and Gar W. Rothwell. Paleobotany and the Evolution of Plants. 2nd ed. — Cambridge: Cambridge University Press, 1993.
Taylor, Thomas N., and Edith L. Taylor. The Biology and Evolution of Fossil Plants. — Englewood Cliffs, N. J.: Prentice-Hall, 1993.
08. Ихтиолог на палеорыбалке
Происхождение позвоночных
Хайкоуихтис
Жаберные щели, язычные хрящи, синаптикулы, эндостиль и хорда. Не поспоришь, что все эти органы отличают протохордовых от морских рыб и указывают им, равно как и нам, на их низкое происхождение. Щитовидная и вилочковая железа, стержень под хордой — вот что объединяет нас с миногами, катранами и сельдью, немногими из тех, что издавна разнообразили наш стол.
Уолтер Гарстанг. «Larval Forms with Other Zoological Verses»
Хью Миллер и древний красный песчаник
Наряду с остальными млекопитающими мы, люди, а также птицы, рептилии, амфибии и рыбы являемся
позвоночными, то есть животными, имеющими хребет. Откуда взялись позвоночные? Что могут поведать о происхождении нашего типа древнейшие ископаемые рыбы? Чтобы получить ответы на эти вопросы, нужно перенестись в Шотландию конца XVIII века.
В конце XVIII века преимущественно в Великобритании начала развиваться новая молодая научная дисциплина — геология. Один из первых шотландских натуралистов Джеймс Хаттон заложил основы современной геологии на материале собственных путешествий по Шотландии. Их итогом в 1788 году стала публикация работы «Theory of Earth» («Теория Земли»), где Хаттон изложил научный подход к вопросу возникновения Земли.
Среди британских стратиграфических подразделений, которые Хаттон тщательно изучал, была мощная серия песчанистых пород, известная как древний красный песчаник. Он широко распространён в Шотландии, также встречается во многих районах восточной и центральной Англии. Чем больше Хаттон его изучал, тем яснее перед ним вырисовывались следы огромного горного хребта, который полностью разрушился, а его породы оказались разнесены ручьями и реками, отложившись в виде галечника и песчаника этой формации. Во многих местах он пролегает почти горизонтально через эрозионную поверхность, врезаясь в более древние породы, которые сначала были выворочены набок, а затем подвергались эрозии, попав из горизонтального положения в вертикальное. Такой пример углового несогласия убедил Хаттона, что мир невероятно стар и зародился «в незапамятные времена». Мир оказался явно древнее 6000 лет — своего возраста согласно Библии; данная оценка во времена Хаттона была общепризнанной.
Догадка учёного оказалась недалека от истины. Сегодня древний красный песчаник датируется девонским периодом (около 345 млн лет назад). Опрокинутые породы ниже углового несогласия по возрасту относятся к силуру (около 425 млн лет назад) и возникли во времена Каледонской складчатости (Каледония — древнеримское название Шотландии). Эта складчатость развернулась, когда ядро Европы (так называемый Балтийский щит) столкнулось с другой тектонической плитой, к которой сейчас относится север Канады и Гренландия. В ходе этого титанического акта горообразования были стёрты в порошок все силурийские породы, сформировавшиеся незадолго до того. Возникшие в результате Каледонские горы быстро разрушились, превратившись в речной песок, который в итоге кристаллизировался в виде древнего красного песчаника. Кэтскиллский песчаник в штате Нью-Йорк аналогичным образом сформировался в результате эрозии Акадских гор, составлявших единый пояс вместе с Каледонскими горами.
Спустя одно поколение после Хаттона древний красный песчаник получил известность благодаря тому, что на него обратил внимание скромный шотландский каменотёс по имени Хью Миллер. Он был сыном капитана, но посещал школу только до 17 лет, то есть не получил систематического образования, достаточного для изучения окаменелостей. На портретах его изображали крепким, широкоплечим, сильным мужчиной (вероятно, такую фигуру он приобрёл, много лет работая с камнем), носившим густые локоны и такие же пышные бакенбарды (рис. 8.1). Молодые годы Миллер провёл в каменных карьерах, прежде всего на древнем красном песчанике. В месяцы, свободные от работы, он прочёсывал прибрежные обнажения песчаника, где одну за другой находил красивых окаменевших рыб. Другие рабочие, трудившиеся там же, вскоре тоже собрали немало образцов, и Миллер взялся изучать находки. К 1834 году кварцевая пыль из карьеров начала гробить его лёгкие, поэтому он бросил работу каменотёса и отправился в Эдинбург, где планировал стать банкиром и писателем.
 Рис. 8.1.
Рис. 8.1. Портрет Хью Миллера (иллюстрация из фонда Wikimedia Commons)
Даже имея ограниченное образование, он стал одним из первых научно-популярных авторов в истории палеонтологии. В 1834 году Миллер опубликовал «Scenes and Legends of North Scotland» («Зарисовки и легенды Северной Шотландии») — настоящий бестселлер о геологии и естественной истории Шотландии, написанный для постоянно ширившейся в те годы читательской аудитории, интересовавшейся естествознанием. Он продолжил работу в 1841 году, написав книгу «The Old Red Sandstone: New Walks in an Old Field» («Древний красный песчаник, или Новые прогулки по старым местам»), где описывает это стратиграфическое подразделение, его удивительных ископаемых рыб и «морских скорпионов». Эту книгу Миллер самостоятельно проиллюстрировал (рис. 8.2). Следующий отрывок позволяет в полной мере оценить его стиль:
Стены в моём чулане наполовину увешаны причудливыми ископаемыми из Нижнего Древнего красного песчаника; поистине вряд ли где-либо была собрана вместе более странная коллекция разных существ; тварей, даже тип которых утерян, диковинных и нескладных — настолько, что натуралист не без труда отнесёт их к тому или иному классу. Животные, похожие на лодки с вёслами и уключинами; панцирные рыбы, напоминающие черепах сверху и снизу, в прочных костяных доспехах, увенчанные в лучшем случае одним веслоподобным плавником; рыбы не столь двусмысленных форм, но с плавниками, густо покрытыми чешуёй. Твари, усаженные шипами, иные, блистающие финифтяными покровами, словно кто-то изысканно их отполировал. Хвост у всех экземпляров — одна из наиболее узнаваемых частей тела, но и он какой-то неровный — у современных рыб хвосты расходятся симметричными половинами по обе стороны от хребта, а у этих тварей хвост растёт книзу от уменьшающегося позвоночника, который продолжается вплоть до самой оконечности плавника. От всех этих форм веет глубокой древностью, периодом, «который давно не в моде».
 Рис. 8.2.
Рис. 8.2. Лопастепёрая рыба глиптолепис (отдалённо родственная амфибиям) и ископаемая двоякодышащая рыба диптерус (гравюра из книги
Hugh Miller. The Old Red Sandstone, or, New Walks in an Old Field. — Edinburgh: Johnstone, 1841)
Благодаря своим книгам Миллер вскоре стал настоящей знаменитостью среди естествоиспытателей, хотя профессиональным палеонтологом не был. К счастью, на заседании Британской ассоциации по содействию научному прогрессу он познакомился с легендарным швейцарским палеоихтиологом Луи Агассисом. Миллер смог передать свои образцы человеку, способному их проанализировать. Агассис действительно вскоре описал и назвал все замечательные окаменелости Миллера.
В своих книгах Миллер продвигал собственные религиозные убеждения и боролся против пришедшего из Франции интереса к эволюционным теориям, которые были готовы расцвести в Британии. Его книга «The Foot-prints of the Creator: or, The Asterolepis of Stromness» («Следы Создателя, или Астеролепис стромнесский») была опубликована в 1849 году и содержала нападки на сенсационные эволюционные идеи, которые в своей книге «Vestiges of the Natural History of Creation» («Пережитки естественной истории творения»), вышедшей в 1844 году, развивал шотландский издатель Роберт Чемберс.
Однако Миллер не был сторонником буквального толкования Библии. Подобно большинству британских геологов своего времени, он считал Ноев потоп локальным событием, охватившим только Месопотамию, а палеонтологическую летопись трактовал как последовательные акты творения и уничтожения, не упомянутые в Библии. Признавая, что в палеонтологической летописи прослеживаются хронологические изменения, он тем не менее не соглашался, что более поздние виды произошли от более ранних.
К сожалению, в возрасте 54 лет Миллера стали мучить странные сильные головные боли и помрачение рассудка, и он застрелился, как только отослал издателю корректуру своей последней книги — «The Testimony of Rocks» («Признание скал»). Научный мир оплакивал его, в честь Миллера устроили одно из самых грандиозных погребальных шествий в истории Эдинбурга. Дэвид Брюстер написал: «Мистер Миллер — один из немногих представителей шотландской науки, который смог восстать над рутиной своей скромной профессии и силой своего гения, а также выдающегося характера занять относительно высокое место в социальной иерархии». В честь него были названы разнообразные окаменелости, в том числе «морской скорпион»
Hughmilleria и примитивная рыба, сегодня именуемая
Millerosteus, а также многие виды рыб с
milleri в названии.
Эпоха рыб
Древний красный песчаник сформировался в девонском периоде — в эпоху рыб, поэтому отражает эволюцию рыб, живших в то время. Там обнаруживаются не только акулы и лучепёрые рыбы, сохранившиеся до наших дней, но многие лопастепёрые и, в частности, двоякодышащие (см. рис. 8.2). Были распространены примитивные челюстные (так называемых панцирные) рыбы, голова и грудь которых были покрыты плотными щитками. Панцирные рыбы полностью вымерли к концу девона.
Эти же окаменелости дали первые доказательства в пользу масштабной распространённости панцирных бесчелюстных рыб. В 1830–1840-е годы Агассис описал некоторых из них, в том числе птерасписа (
Pteraspis) и цефаласписа (
Cephalaspis) (рис. 8.3). Миллер заявлял, что обнаруженные им окаменелости рыб не являются доказательством эволюции, но был недостаточно хорошим анатомом, чтобы судить об этом. Тем не менее присутствие этих бесчелюстных позвоночных в девоне свидетельствовало, что современных челюстных рыб от бесчелюстных беспозвоночных отделяет несколько витков эволюции.
 Рис. 8.3.
Рис. 8.3. Панцирная бесчелюстная рыба цефаласпис (гравюра из книги
Hugh Miller. The Old Red Sandstone, or, New Walks in an Old Field. — Edinburgh: Johnstone, 1841)
Вскоре окаменелости этих панцирных бесчелюстных рыб обнаружили во многих других регионах. Они ещё раз показали, как челюстные позвоночные эволюционировали от бесчелюстных предков (см. рис. 8.4). Птераспис и его родичи (гетеростраки) обычно имели обтекаемое, покрытое панцирем тело, по форме напоминавшее торпеду. Из боков и спины такой рыбы часто торчали длинные шипы, а хвост с главным плавником смотрел вниз (см. рис. 8.5). У гетеростраков был крошечный рот, напоминавший щель, и не было челюстей, равно как отсутствовали и сильные мускулистые плавники, служившие бы рулём. Вероятно, они плавали как головастики, всасывая воду и отфильтровывая питательные частицы, проходившие через рот и жабры. Напротив, цефаласписы (см. рис. 8.3 и 8.4) и родственные им остеостраканы (также называемые остракодермами) имели покатую голову с плоским низом, а главный плавник у них на хвосте смотрел вверх (как у современных акул). Считается, что они плавали у дна и рылись в иле в поисках пищи, фильтруя осадок через бесчелюстную пасть.
 Рис. 8.4.
Рис. 8.4. Генеалогическое древо бесчелюстных рыб, на котором показаны разные группы (рисунок Карла Бьюэлла из работы:
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 9.8)

 Рис. 8.5.
Рис. 8.5. Бесчелюстная панцирная рыба птераспис, гетерострак: (
А) головной щит; (
Б) реконструкция живой рыбы (иллюстрация (
А) ив фонда Wikimedia Commons; иллюстрация (
Б) представлена Nobumichi Tamura)
Удочка в прошлое
С годами учёные находили всё больше окаменелостей панцирных бесчелюстных рыб в девонских, а затем и в силурийских отложениях по всему миру. Но единственной частью их тела, которая легко фоссилизировалась, был наружный панцирь. Подобно акулам и большинству примитивных рыб, они не имели костного скелета, только хрящевой, который плохо сохраняется в окаменелостях. Если бы не панцирь, ни одна из этих рыб не могла бы сохраниться в палеонтологической летописи.
Довольно долго отсутствовали какие-либо доказательства существования бесчелюстных (и любых других) рыб ранее силура. В ордовикских морях господствовали крупные хищники, например наутилоидеи длиной 5,5 м. Однако, несмотря на изобилие ископаемой ордовикской морской фауны, там не удавалось найти и следа костей. Единственными намёками на их существование были редкие находки, как в песчанике Хардинг близ Каньон-Сити (штат Колорадо). Эта порода датируется средним ордовиком и содержит массу осколков костного панциря, принадлежавшего бесчелюстной рыбе астрапису (
Astrapis). К 1970–1980-м годам удалось найти полные экземпляры этих древнейших позвоночных, например арандасписа (
Arandaspis) из Австралии и сакамбасписа из Южной Америки, позднее также обнаруженного в Австралии.
Все эти ордовикские бесчелюстные рыбы были устроены не сложнее фильтраторов, покрытых тонкими пластинами костного панциря. Их широкие плоские тела почти не имели выдающихся частей — каких-либо плавников или шипов. Только щелевидный рот для всасывания воды, которая изобиловала пищей, и простой асимметричный хвост. Вместо пластинчатого панциря, как у птерасписа, эти рыбы были покрыты сотнями крошечных костяных кусочков, которые напоминали кольчугу. Крошечные глазки и ряды канальцев на поверхности тела (боковые линии) позволяли им улавливать движение воды вокруг. Все эти ордовикские рыбы встречаются исключительно редко по сравнению с большинством других животных того времени. Удручало и то, что не была известна ни одна подобная рыба из кембрия.
Наконец, в 1970-е Джек Репецки, палеонтолог из Геологической службы США, стал работать с микроокаменелостями, так называемыми конодонтами, из песчаника Дэдвуд, что в штате Вайоминг, датируемого поздним кембрием. Растворяя обызвествлённые окаменелости в поисках конодонтов (состоят из фосфата кальция, как и кости позвоночных), он обнаружил фрагменты интересной формы и понял, что перед ним — кусочки кожистого панциря бесчелюстной рыбы анатолеписа (см. рис. 8.6). Позднее развернулись долгие споры о том, на самом ли деле эти образцы принадлежат позвоночному. В итоге они разрешились, и сегодня анатолепис считается древнейшим известным позвоночным, от которого остались костные окаменелости.
 Рис. 8.6.
Рис. 8.6. Отдельно взятый небольшой фрагмент пластины (около миллиметра в диаметре) с кожистого панциря кембрийской бесчелюстной рыбы анатолеписа (Anatolepis) — одного из древнейших позвоночных, обладавшего костями (иллюстрация предоставлена Геологической службой США)
Соединяя звенья
Итак, постепенно находя окаменелости позвоночных во всё более древних породах, мы, наконец, оказываемся в отложениях, которые сформировались до появления костей. В настоящее время фрагменты кожистого панциря анатолеписа по-прежнему остаются древнейшими ископаемыми останками существа, обладавшего костями. Все более древние животные были мягкотелыми, состояли из хрящей и менее прочных тканей, которые могли фоссилизироваться лишь изредка, в наиболее благоприятных условиях.
Поскольку дальнейших находок костных окаменелостей не последовало, биологи и палеонтологи попытались соединить эволюционные звенья между позвоночными и их предками, причём решили делать это снизу вверх.
Здесь мы не испытываем недостатка в материале, так как многие промежуточные организмы, связывающие позвоночных с остальным царством животных, сохранились до наших дней, а от других остались многочисленные окаменелости. Млекопитающие, птицы, рептилии, амфибии и рыбы принадлежат к типу хордовых, которые именуются так, поскольку их эмбрионы (а иногда и взрослые особи) обладают длинной гибкой хрящевой струной (хордой), идущей вдоль спины и поддерживающей всё тело. Хорда — предшественница позвоночника.
Ближайшие родственники хордовых относятся к другому типу, полухордовым (рис. 8.7). Сегодня они представлены кишечнодышащими и перистожаберными. Кишечнодышащие (они же кишечножаберные) напоминают неподготовленному наблюдателю обычных червей, но их эмбрионы обладают зачатками хорды и настоящей глоткой (фарингосом), которая есть у всех хордовых. Кроме того, их нервный ствол идёт вдоль спины, а пищеварительный тракт — вдоль брюха; такая конфигурация характерна для большинства хордовых, тогда как у беспозвоночных она противоположная: нервный ствол идёт вдоль брюха, пищеварительный тракт — вдоль спины. Это анатомическое сходство подкрепляется эмбриологическими чертами, свойственными хордовым. Наконец, молекулярный анализ ДНК показывает, что полухордовые очень близки к общему предку всех позвоночных, а также к ближайшим беспозвоночным родичам — иглокожим (к ним относятся морские звёзды, голотурии, морские ежи и пр.).
 Рис. 8.7.
Рис. 8.7. Происхождение хордовых от беспозвоночных; исходная версия, описанная Уолтером Гарстангом и Альфредом Ш. Ромером более века назад. Варианты строения многих взрослых особей (например, взрослых оболочников) оказались тупиковыми эволюционными ветвями, но личинки оболочников сохраняют длинный хвост и другие черты, позже сыгравшие важную роль в происхождении более совершенных хордовых (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 9.4)
Следующее звено на пути к позвоночным — группа, представленная более 2000 видов, обитающих в океанах всего мира. Речь идёт об оболочниках или асцидиях (рис. 8.7). Как и кишечнодышащие, асцидии не напоминают неискушённому наблюдателю рыб, но внешнее впечатление обманчиво. Взрослые особи у них неказистые, похожи на мешочек
киселя и фильтруют морскую воду через своё тело-корзинку. Но личинки оболочников очень напоминают рыб или головастиков; у них есть хорошо развитая хорда, длинный мускулистый хвост с парными мышцами, а также голова с большой глоткой. Это лишь некоторые их важнейшие черты. Опять же, направление эволюции прослеживается на уровне эмбрионов, а эмбриологические доказательства подкрепляются молекулярными, чётко демонстрирующими, что оболочники ближе к позвоночным, чем какие-либо другие морские беспозвоночные.
Последнее звено, связывающее беспозвоночных с позвоночными, — ланцетники (
Branchiostoma) (см. рис. 8.7). Этот неприметный кусочек плоти обычно имеет всего несколько сантиметров в длину, но если к нему присмотреться, оказывается, он очень похож на рыбу, хоть рыбой и не является. У ланцетников длинная гибкая хорда, поддерживающая тело и снабжённая множеством V-образных мышечных связок, расположенных по всей длине животного; благодаря этим связкам оно отлично плавает. Нервные волокна ланцетника идут вдоль спины, а пищеварительный тракт — вдоль брюха, как у всех хордовых. Нет ни челюстей, ни зубов, но рот ведёт к глотке с жаберным мешком, куда животное захватывает пищу. У ланцетников нет настоящих глаз, только светочувствительное пятно на передней оконечности тела, позволяющее им различать свет и тень. Эти существа проводят всю жизнь, закопавшись хвостом в донный ил, высунув лишь голову и хватая ртом частицы пищи.
Наконец, несколько хорошо сохранившихся окаменелостей ланцетников демонстрируют, что эти животные существовали уже в раннем кембрии — именно в тот период, когда начиналась эволюция рыб. Среди этих окаменелостей — пикайя (
Pikaia), найденная в сланцах Бёрджесс в Канаде (см. главу 6), и схожая окаменелость юннанозоон (
Yunnanozoon), принадлежащая к Шэньянской фауне (Китай), датируемой ранним кембрием (518 млн лет назад).
Нерыбоподобное звено
Итак, мы восстановили родословную позвоночных до бесчелюстных рыб, живших в ордовике, по пути миновали девон и добрались до самых давних костных окаменелостей, относящихся к позднему кембрию. Древнейшие рыбы были мягкотелыми, поэтому мы не располагаем более ранним ископаемым костным материалом. Мы поднимались от корней эволюционного древа мягкотелых хордовых — там нам встретились кишечнодышащие (полухордовые), далее — оболочники и, наконец, ланцетники, которые сильно напоминают рыб, но лишены некоторых важнейших анатомических признаков, таких как чётко выраженная голова, двухкамерное сердце и эмбриологическая черта — клетка нервного гребня, позволяющая отнести животное к позвоночным. Нам требуется лишь найти животное, которое было бы мягкотелым, обладало большинством признаков позвоночных, но при этом не имело бы костного панциря, — и получилась бы непрерывная цепочка.
Действительно, в 1999 году несколько китайских учёных вместе с Саймоном Конвеем Моррисом сообщили об открытии окаменелостей, названных Хайкоихтис (рыба из Хайкоу) и относящихся к раннему кембрию (518 млн лет назад). Эта рыба принадлежит к Шэньянской фауне на территории Китая. К ней же принадлежит юннанозоон, ископаемый ланцетник. Хайкоуихтис имел длину всего около 2,5 см, но его окаменелости демонстрируют ряд примечательных черт (рис. 8.8). У этого животного есть явно выраженная голова (не то что у ланцетника), за которой расположены жабры и жаберные щели — до девяти с каждой стороны. У рыбы есть короткая хорда, а на длинном цилиндрическом теле имеется широкий спинной плавник, идущий от середины хребта до кончика хвоста. У основания хвоста находится брюшной плавник. Плавники поддерживают радиальные хрящи, как у других бесчелюстных рыб, например у миног и миксин.

 Рис. 8.8.
Рис. 8.8. Хайкоихтис (
А) окаменелость; (
Б) реконструкция — как могла бы выглядеть живая рыба (иллюстрация (
А) предоставлена D. Briggs; иллюстрация (
Б) предоставлена Nobumichi Tamura)
В том же докладе описывается более примитивная рыбоподобная окаменелость, относящаяся к Шэньянской фауне Китая. Эта рыба, получившая название миллокуньминия, тоже обладает чётко очерченной головой, хрящевым черепом, пятью или шестью жаберными щелями за головой, хордой, пролегающей вдоль спины, а также длинным парусообразным спинным плавником, идущим от головы до кончика хвоста; под хвостом у неё — набор парных брюшных плавников. Известен всего один экземпляр миллокуньминии, причём не очень хорошо сохранившийся, поэтому трудно судить о ней с уверенностью. Однако имеющиеся данные позволяют предположить, что это ещё более примитивное хордовое, чем хайкоуихтис.
Наконец, третье существо из нижнекембрийских отложений называется хайкоуэлла (
Haikouella). Её длина составляла от 20 с лишним до 40 мм; известно более 300 экземпляров. У этого животного определённо была голова, мозг, жабры и хорда, поддерживавшая хорошо развитые мышцы туловища, идущие до самого хвоста. Кроме того, сердце и кровеносная система, а также длинный спинной плавник вдоль всего тела и хвоста, небольшие брюшные плавники. Некоторые экземпляры позволяют предположить, что по бокам головы у этой рыбы имелись глаза — первые глаза у хордовых, если гипотеза верна.
Итак, раннекембрийские отложения в Китае поведали нам о множестве мягкотелых хордовых, которые явно принадлежат к эволюционному древу позвоночных и были более развитыми существами, чем ланцетники (рис. 8.9). Им оставалось нарастить лишь небольшой костный панцирь. Обретя его, эти животные превратились в бесчелюстных панцирных рыб, которых Хью Миллер открыл почти 200 лет назад. Переход от беспозвоночных, таких как кишечнодышащие и оболочники, к первым настоящим рыбам теперь полностью восстановлен, недостающих звеньев или пробелов на данном отрезке палеонтологической летописи не осталось.
 Рис. 8.9.
Рис. 8.9. Этапы эволюции от кишечнодышащих к ланцетникам, хайкоуэлле, хайкоуихтису и, наконец, к костистым бесчелюстным анапсидам (рисунок Карла Бьюэлла на основе работы
D.-G. Shu et al. Lower Cambrian Vertebrates from South China //
Nature. — 1999. — November 4)
Лучше один раз увидеть!
Раннекембрийские окаменелости из Китая не выставляются. Однако во многих музеях вы найдёте отличные экспозиции древних ископаемых рыб. Среди них: Американский музей естественной истории (Нью-Йорк), Кливлендский музей естественной истории, Музей естественной истории под открытым небом (Чикаго), Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия). В Элджинском музее (Элджин, Шотландия) находится крупнейшая коллекция ископаемых рыб и других окаменелостей, добытых из расположенного поблизости Древнего красного песчаника, а также большой архив книг, статей и заметок Хью Миллера (http://elginmuseum.org.uk/museum/collections-fossils/).
Дополнительная литература
Forey, Peter, and Philippe Janvier. Evolution of the Early Vertebrates //
American Scientist. — 1984. — Vol. 82. — 554–565.
Gee, Henry. Before the Backbone: Views on the Origin of Vertebrates. — New York: Chapman & Hall, 1997.
Long, John A. The Rise of Fishes: 500 Million Years of Evolution. — Baltimore: Johns Hopkins University Press, 2010.
Maisey, John G. Discovering Fossil Fishes. — New York: Holt, 1996.
Moy-Thomas, J. A., and R. S. Miles. Palaeozoic Fishes. — Philadelphia: Saunders, 1971.
Shu, D.-G., H.-L. Luo, S. Conway Morris, X.-L. Zhang, S.-X. Hu, L. Chen, J. Han, M. Zhu, Y. Li, and L.-Z. Chen. Lower Cambrian Vertebrates from South China //
Nature. — 1999. — November 4. — 42–46.
09. Мегачелюсти
Самая крупная рыба
Мегалодон
У акулы есть всё, что может привести в восторг учёного. Они красивы, боже, как они красивы! Акула — это невероятно чёткий, удивительно отлаженный механизм. Они грациозны, как птицы, и непостижимо загадочны, как любое другое существо на земле. Никто не знает, сколько они живут и каким инстинктам — за исключением голода — подчиняются. Существует более двухсот пятидесяти разновидностей акул, и как сильно они отличаются друг от друга!
Питер Бенчли. «Челюсти»
Прогулка на холм Шарктус
В конце 1950-х и 1960-е, годы моего детства, проведённые в Южной Калифорнии, я был без ума от динозавров и прочих доисторических существ. Тогда я был младшим бойскаутом, исходил все важнейшие россыпи окаменелостей, расположенные поблизости; в том числе ракушняки в каньоне Топанга и залежи в каньоне Ред-Рок, содержащие останки млекопитающих (и те и другие относятся к миоцену). Но мне снова и снова доводилось слышать истории о легендарном холме Шарктус близ Бейкерсфилда, где акульи зубы и морские окаменелости скапливались в глубоких водах, плескавшихся на месте древней Калифорнийской долины 16–15 млн лет назад. Однако никто не знал, как туда добраться и пособирать окаменелости: большая часть этих залежей находилась на территории частных владений, где повсюду стояли заборы с табличками «Вход воспрещён».
В течение следующих 30 лет научной карьеры я занимался другими вещами, пока около 1997 года коллеги мне не сказали, что местный фермер Боб Эрнст разрешает студенческим группам и исследователям из некоммерческих организаций собирать образцы на его участке. Мне удалось побеседовать с Эрнстом лично, и вскоре мой палеонтологический курс из колледжа Оксидентал, а также два курса из Калифорнийского технологического института начали полным составом останавливаться в его усадьбе, чтобы побывать на холме Шарктус в рамках учебных экспедиций. В 2002 году я понял, что на этом холме нужно провести гораздо больше исследований, чем уже было сделано. Мы со студентами применяли технологию магнитной стратиграфии: измеряли разницу показателей магнитного поля Земли в разных породах, чтобы датировать отложения точнее, чем когда-либо. Мы работали вместе с Ларри Барнсом из Музея естественной истории округа Лос-Анджелес (настоящий ветеран Шарктуса, который начал там трудиться ещё в начале 1960-х) и многими другими, стараясь идентифицировать разнообразных сухопутных зверей, чьи останки оказались в глубокой воде, а затем были погребены и фоссилизировались вместе с окаменелостями акул и морских млекопитающих. Результаты этих исследований опубликованы (в основном в 2008 году) в соавторстве со студентами, и теперь мы ориентируемся в данных отложениях лучше, чем ранее.
Визит на легендарные россыпи костей — настоящее откровение. Сначала проезжаете одно за другим огромные нефтяные месторождения (пока держите путь на северо-восток, из Бейкерсфилда к подножию Сьерра-Невады). Месторождения нефти вокруг Бейкерсфилда по-прежнему активно разрабатываются, и, кроме того, они в числе крупнейших в Калифорнии. Наконец, добираетесь до поворота с грунтовой дороги к воротам на ранчо. Те заперты на замок, открываемый и закрываемый специальным кодом. Едете ещё милю-две по грунтовой дороге среди низких покатых холмов, покрытых травой и колючим кустарником, — и вдруг видите места, основательно разрытые бульдозером. Выпрыгиваете из машины, хватаете своё снаряжение и начинаете ползать по костным отложениям.
Практически всё, что вам понадобится из инструментов, — шило или подобная вещь, чтобы прощупывать мягкий песок, а также метёлочка или рисовальная кисточка, чтобы стряхивать пыль с находок. Время от времени Боб Эрнст вызывал бульдозериста. Тот приезжал, вычищал вскрышу (пустую породу), обнажая слой, полный окаменелостей, и оставлял его для обработки палеонтологами. Многие приходят работать в респираторе, так как почва в этих местах может быть заражена лихорадкой долины Сан-Хоакин. Это тяжёлая лёгочная болезнь, вызываемая грибными спорами. Большую часть года тут приходится носить шляпу и свободную одежду, чтобы защититься от палящего солнца, и использовать крем от загара. Также не помешает хорошая подушка для сидения или валик — вам придётся провести не один час на жёсткой поверхности.
Но дело того стоит! Здешний грунт сложен из твёрдых фрагментов костей и зубов (более 200 образцов на кубический метр породы). Иногда можно найти целый череп или скелет кита, а вокруг — сравнительно рыхлый песчаный матрикс, который относительно легко разгребать. Никаких прочных долот и кирок! С каждым новым заходом вы будете находить всё больше мелких акульих зубов. Кстати, нужны перчатки, потому что акульи зубы по-прежнему острые и ими можно порезаться, если неосторожно копаться в песке. Пусть акулы на Шарктусе давно вымерли, они по-прежнему кусаются!
Подавляющее большинство зубов принадлежит разным видам акул-мако (
Isurus), а вообще здесь были найдены зубы около 30 других видов акул (рис. 9.1). Вы найдёте множество отдельных костных фрагментов, которые нельзя классифицировать, а также сильно повреждённые китовые позвонки, которые никто не берёт, поскольку по ним невозможно ничего определить или узнать. Иногда попадаются сильно обызвествлённые ушные кости китов (сильно отличаются у разных видов), а ещё реже — части других морских млекопитающих, которые определённо стоит сохранить. В этих костных отложениях в изобилии встречаются разнообразные останки морского зверя, принадлежащие в том числе десяткам видов китов и дельфинов, разным видам древних тюленей и морских львов и вдобавок диковинным животным, таким как вымершие десмотилии, напоминавшие гиппопотамов, и исчезнувшие родичи ламантинов. Но самый большой приз — огромный треугольный зуб гигантской акулы мегалодона (
Carcharocles megalodon). Боб Эрнст разрешал посетителям забирать со своего ранчо любые другие окаменелости, которые они найдут (палеонтологам-музейщикам отдавал даже хорошо сохранившиеся китовые черепа), но не зубы мегалодона. Их он оставлял себе, так как они пользуются спросом у коллекционеров, и от продажи этих зубов Эрнст оплачивает счета. Взамен фермер разрешал людям копаться на ранчо и пользоваться его гостеприимством. В 2007 году мой добрый друг Боб скоропостижно скончался, и ситуация изменилась.
 Рис. 9.1.
Рис. 9.1. Типичные зубы с холма Шарктус, в том числе зуб мегалодона, вокруг которого лежат зубы самого распространённого здесь вида акул мако (Isurus) (фотография предоставлена R. Irmis из Палеонтологического музея Калифорнийского университета)
Костные отложения холма Шарктус долго оставались загадкой. Насколько они древние? Как сформировались? Какая здесь была глубина? Каким образом в одном слое могло накопиться такое количество зубов и костей? Барнс в основном разрешил эти проблемы, причём давно, а благодаря недавним исследованиям Николаса Пайенсона из Смитсоновского института и моим удалось ответить и на большинство других вопросов.
Начнём с самого простого. Наша палеомагнитная датировка показала, что участок алевролитов Раунд-Маунтин, к которому относятся эти отложения, сформировался от 15,9 до 15,2 млн лет назад, поэтому возраст костных залежей — около 15,5 млн лет. Микроокаменелости в алевролите свидетельствуют, что здесь было очень глубоко (минимум 1000 м).
Но откуда такая концентрация костей? Видимо, в глубоководной акватории, располагавшейся здесь в миоцене, исключительно медленно накапливался осадок, поскольку костный слой считается задержанным наносом, то есть долговременным скоплением костей и зубов, которые накапливались на морском дне, почти не перемешиваясь с осадком. Очевидно, в силу геологического строения этой местности большая часть грязи и песка, вымываемая с суши, не попадала на дно или просто огибала его.
Практически все окаменелости изломаны или расчленены — соответственно, животные погибали или были разорваны на части до того, как попадали на дно. Там они накапливались вместе с акульими зубами, которые у акулы регулярно выпадают и заменяются новыми. Однако иногда встречаются полные, неповреждённые скелеты китов и других морских млекопитающих — то есть трупы могли опускаться на дно нетронутыми, не доставаясь падальщикам. Возникают и так называемые китовые оазисы. Накопление костей происходило в период среднемиоценового климатического оптимума, когда тёплый глобальный климат поспособствовал огромной эволюционной радиации планктона, морской флоры и фауны, особенно китов во всём мире. Благодаря таким условиям здесь не только паслись огромные стада китов (а где киты, там и акулы), но и осадок накапливался медленнее, чем на более ранних и поздних этапах миоцена.
Разнообразие здешних окаменелостей поражает: найдено не менее 150 видов позвоночных и более 30 видов акульих зубов, хотя абсолютное большинство составляют зубы акул мако (см. рис. 9.1). Здесь обитали огромные морские черепахи, втрое крупнее кожистых черепах — самых больших современных рептилий. На других участках алевролитов Раунд-Маунтин, прежде всего расположенных ниже этих, на мелководных песках Ольсез, находят разнообразных двустворчатых и одностворчатых моллюсков. В отложениях попадаются окаменелости минимум 30 видов морских млекопитающих.
Но больше всего мы с коллегами удивились тому, сколько всевозможных скелетов сухопутных млекопитающих попало в воду и утонуло. В результате более чем векового сбора окаменелостей в музейных коллекциях скопилось много разнообразных ископаемых, в основном неопределённого происхождения. В 2008 году мы с Ларри Барнсом, Ричардом Тедфордом, Клейтоном Реем, Сэмюэлем Маклеодом, Мэтью Лайтером, Дэвидом Уистлером и Сяомином Вонгом, наконец, все их опубликовали. На подготовку ушли десятилетия. Там есть мастодонты, два вида носорогов, тапиры, множество лошадей и верблюдов; дромомерициды, напоминающие оленей; настоящие кошачьи, псовые, куньи и вымершие амфиционы. Все эти млекопитающие уже найдены и в других, находящихся неподалёку отложениях среднего миоцена — например, в Барстоу и каньоне Ред-Рок в Калифорнии, а также в других местах, разбросанных по всему западу США (особенно в регионе Великих Равнин — штатах Небраска, Вайоминг и Южная Дакота). Работая над этим проектом, я возил с собой самые интересные образцы, летая самолётами из города в город, и, чтобы идентифицировать мои окаменелости, сравнивал их с экспонатами из местных музеев.
Кишащие акулами миоценовые моря
Гигантские акулы, подобные тем, что встречались в районе Шарктус-Хилл, обитали в морях по всему миру. Их окаменелости в большом количестве обнаруживаются на знаменитом руднике Ли-Крик в Северной Каролине, в отложениях Костяной долины во Флориде, ракушняках Колберт-Клиффз по берегам Чесапикского залива, а также во многих других классических местонахождениях миоценовой фауны на территории США. Также их находят в Европе, Африке и на многих островах Карибского моря, в том числе на Кубе, в Пуэрто-Рико и на Ямайке. Зубы
мегалодона разбросаны по всему миру, от Канарских островов до Австралии, Новой Зеландии, Японии и Индии. Их извлекали даже из глубин Марианского желоба, расположенного в Тихом океане близ Филиппин. Считается, что древнейшие образцы добыты из олигоценовых отложений, чей возраст составляет около 28 млн лет. Эти зубы чаще всего встречаются в породах, формировавшихся в условиях сравнительно тёплого климата, характерного для раннего-среднего миоцена, но попадаются и в плиоценовых отложениях (5–2 млн лет назад). Самые молодые известные образцы таких зубов имеют возраст около 2,6 млн лет.
Проблема с изучением акул заключается в том, что зубы — единственные костные фрагменты их организма. Поэтому большинство ископаемых акул известны по зубам, и только по ним. Остальной «скелет» состоит из хрящей, которые фоссилизируются редко (рис. 9.2). Иногда акулий позвоночник частично обызвествляется, благодаря чему известно несколько акульих хребтов, в том числе несколько хребтов мегалодона. А мелкие детали зубов — основа классификации большинства ископаемых акул, у которых не осталось ныне живущих потомков. К счастью, мы располагаем подробными данными о зубах современных акул, поэтому их родственные связи можно восстановить по имеющимся в избытке мягким тканям. Затем большинство ископаемых акульих зубов можно соотнести с видами современных акул. В данном контексте взаимосвязи видов проясняются.
 Рис. 9.2.
Рис. 9.2. Реконструированный хрящевой «скелет» мегалодона длиной более 10 метров (фотография предоставлена Stephen Godfrey, Калвертский морской музей, Соломонс, Мэриленд, США)
Однако с мегалодонами существует проблема. Когда Луи Агассис в 1835 году изучил первые связанные с ними ископаемые образцы, он причислил животное к роду кархародон, к которому относится современная большая белая акула (
Carcharodon carcharias). Своими простыми очертаниями эти широкие треугольные зубы напоминали зубы большой белой акулы, только увеличенные. Мнение об их родстве считалось общепринятым на протяжении десятилетий; до недавнего времени его придерживалось большинство учёных. Однако в последнее десятилетие ряд специалистов по акулам стали отмечать, что мегалодон родственен не большой белой акуле (
Carcharodon), а вымершей акуле
Carcharocles — несколько иному представителю ламнообразных акул (к этому семейству также относятся мако и другие виды). Некоторые даже считают, что мегалодон произошёл от ископаемой акулы
Otodus и должен быть включён в этот род. В настоящее время представляется, что сообщество палеонтологов, изучающих акул, в целом отдаёт предпочтение
Carcharocles относительно других вариантов. Такой точки зрения буду придерживаться и я в рамках данной главы. Хотя главу можно было бы с тем же успехом назвать кархародоном, и большинство палеонтологов ничего не имели бы против.
Вот такая рыба!
Как бы мы его ни называли, мегалодон был исполинским хищником, вероятно, крупнейшим из всех, когда-либо обитавших в океане. Он был намного крупнее самой большой современной рыбы — китовой акулы (
Rhincodon typus), которая совершенно безобидна и питается планктоном, заглатывая его с большими объёмами воды. Не сравнится с мегалодоном и гигантская акула — вторая по величине из ныне живущих (
Cetorhinus maximus). Есть предположение, что рыба
лидсихтис, обитавшая в юрском периоде, могла быть крупнее, но известные экземпляры лидсихтиса слишком фрагментарны, чтобы с уверенностью судить о его размерах. В настоящее время считается, что максимальная длина лидсихтиса могла достигать 16 м.
Здесь мы опять же сталкиваемся с проблемой, так как в нашем распоряжении есть лишь зубы и несколько обызвествлённых, частично сохранившихся хребтов мегалодона. Все предположения о его размерах делаются исходя из того, каков масштаб акульего зуба относительно тела животного. Такие оценки осложняются ещё и тем, что ранее палеонтологи были склонны реконструировать челюсти мегалодона (хрящевые и потому не сохранившиеся) на основании самых крупных зубов, имевшихся в коллекции, без использования более мелких боковых зубов, уступающих по размеру передним. Соответственно, знаменитая модель челюстей мегалодона, однажды воздвигнутая в Американском музее естественной истории, была, видимо, преувеличена, поскольку строилась лишь на основании передних зубов (рис. 9.3).
 Рис. 9.3.
Рис. 9.3. Знаменитая реконструкция челюстей мегалодона, выполненная Бэшфордом Дином в Американском музее естественной истории около 100 лет назад, с использованием лишь самых крупных зубов. Сегодня такая модель кажется преувеличенной, поскольку не включает более мелкие боковые зубы (изображение № 336000 из библиотеки Американского музея естественной истории)
С учётом всех названных проблем палеонтологи разработали весьма хитроумные способы, позволяющие оценить размеры мегалодона (рис. 9.4). Первая модель, сделанная Бэшфордом Дином из Американского музея естественной истории, предполагала завышенный размер челюстей (см. рис. 9.3). В соответствии с данной реконструкцией акула могла достигать 30 м в длину. При использовании другого метода сравнивается высота эмали на зубах крупнейших известных акул, и получается, что мегалодон был значительно мельче (13 м). В 1996 году Майкл Готтфрид и несколько других специалистов рассмотрели 73 экземпляра точно измеренных белых акул и вывели формулу определения длины тела на основе размеров крупнейшего зуба. Самый большой зуб мегалодона, оказавшийся у них в распоряжении, имел длину всего 168 мм, и, таким образом, общая длина акулы составила 16 м. Однако в настоящее время найдены зубы мегалодона длиной до 194 мм, что даёт оценку ближе к 20 м. В 2002 году Клиффорд Джеремия попытался оценить длину акулы исходя из величины основания зуба у самого корня и в результате получил длину 16,5 м, хотя зубы, которые он использовал для расчёта, были не самыми крупными из известных. Кеншу Шимада тоже в 2002 году применил ещё один метод, при котором определял длину тела, масштабируя высоту коронковой части зуба. По измерениям самых крупных зубов он вычислил, что акула могла достигать 17,9 м. Однако Патрик Шембри и Стэфон Папсон предположили, что крупнейшие экземпляры могли вырастать до 24–25 м в длину, что практически совпадает с первой завышенной оценкой, сделанной Бэшфордом Дином около ста лет назад.
 Рис. 9.4.
Рис. 9.4. Сравнение размеров акул, в том числе большой белой акулы (
Carcharodon carcharias) и китовой акулы (
Rhincodon typus) — крупнейшей из ныне живущих рыб, а также две разные оценки длины мегалодона (рисунок Mary P. Williams)
 Рис. 9.5.
Рис. 9.5. Реконструкция
Carcharoles megalodon в натуральную величину, выставленная в Музее естественной истории города Сан-Диего (фото автора)
Короче говоря, оценка длины мегалодона — сложная задача, которую можно решить несколькими способами. Но учёные сходятся на том, что эта акула определённо достигала 16 м и, вероятно, могла вырастать до 25 м. Даже консервативная оценка превышает 12,7 м — длину крупнейших известных экземпляров современной китовой акулы, а также шестнадцатиметровую длину лидсихтиса. Поэтому, независимо от метода оценки, мегалодон был крупнейшей рыбой, когда-либо обитавшей в океанах.
Оценив длину тела, можно попытаться вычислить массу такой рыбы. Готтфрид с коллегами изучили распределение длины по сравнению с массой тела для 175 экземпляров большой белой акулы на разных стадиях роста и вывели формулу, позволяющую вычислять массу по длине тела. Мегалодон длиной 16 м должен был весить около 48 т, 17-метровый весил бы 59 т, а 20,3-метровый — 103 т.
Хотя до сих пор найдены лишь зубы и несколько частично минерализованных скелетов мегалодона, хрящевой скелет этого чудовища можно реконструировать, масштабируя размеры скелета современной большой белой акулы. Такая реконструкция была сделана и выставлена (см. рис. 9.2) в Калвертовском морском музее на острове Соломонс (штат Мэриленд). Там есть множество интересных миоценовых окаменелостей из скал Калверт-Клиффз и с берегов Чесапикского залива. В некоторых музеях (например, в Музее естественной истории Сан-Диего) воссоздали «чучела» мегалодона в натуральную величину.
Гроза морей
Огромные размеры мегалодона заставляют задуматься, почему он вырастал таким гигантом? Наиболее распространённое объяснение таково: вероятно, в миоцене акулы не испытывали недостатка в крупной добыче; особенно этому поспособствовала массовая радиация разнообразных китов и дельфинов, которая пришлась на ранний и средний миоцен. Мегалодон превосходил по размеру почти всех китов, находимых в тех же отложениях (кроме самых крупных), был истинным суперхищником, способным убивать и поедать практически любых обитателей миоценового океана.
Тому есть многочисленные ископаемые свидетельства. Глубокие борозды и царапины, которые могли оставить только зубища мегалодона, встречаются на многих фоссилизированных китовых костях. Акула глодала кости, отрывая плоть от скелета. Список китообразных со следами нападений мегалодона очень велик: это и дельфины, и другие мелкие киты, и цетотерии, сквалодонты, кашалоты, гладкие киты, полосатики (в их числе финвал и синий кит), а также тюлени, морские львы, ламантины и морские черепахи (втрое превосходившие размерами крупнейших современных черепах). Зуб мегалодона был найден в окаменелости вместе с покусанной ушной костью морского льва. Кроме того, обнаружили несколько зубов мегалодона в хребтах китов, а также многочисленные и частично обглоданные скелеты китов (особенно в районе холма Шарктус) в окружении мегалодоньих зубов.
Разумеется, список этим не исчерпывается. Большинство акул (особенно большие белые) всеядны и питаются чем придётся. Они нападают на любые движущиеся предметы, которые в состоянии схватить. Вот почему в наше время при вскрытии многих акул в их брюхе находят разнообразный мусор (в том числе дорожные знаки, обувь и крючья). Мегалодон явно мог есть более мелкую рыбу и небольших акул, если они ему попадались. Но гигантизм — это, прежде всего, адаптация к охоте на большую дичь, китов в первую очередь, которым до появления мегалодонов был не страшен ни один морской хищник.
Следы от укусов на теле одного ископаемого кита, имевшего около 9 м в длину, показывают, как предпочитал нападать мегалодон. Укусы сосредоточены в местах с крупными костями (плечи, ласты, грудная клетка, верхний отдел позвоночника), а не на мягком подбрюшье, куда метят современные большие белые акулы. Таким образом, мегалодон пытался раздавить или прокусить сердце либо лёгкие кита и быстро его умертвить. Это, в свою очередь, объясняет, почему зубы мегалодона были такими толстыми и мощными: они развивались для раскусывания костей. Кроме того, мегалодон часто хватал жертву за ласты, поскольку следы укусов чаще всего встречаются на окаменелостях костей верхних конечностей. Если акуле удавалось одним махом сильно перемолоть, растрепать или оторвать один из плавников, добыча в результате такого увечья практически не могла сопротивляться, и акула легко приканчивала её ещё несколькими укусами.
Хищнический образ жизни этих гигантских акул подсказывает, почему они постепенно вымерли в позднем миоцене и плиоцене. Хотя в среднем миоцене они и находились на вершине пищевой цепочки, к началу плиоцена появились более крупные киты, покуситься на которых мегалодоны не могли, а также крупные хищные киты — в частности, сквалодонты и кашалоты. Позднемиоценовый кашалот
Livyatan melvillei был настоящим гигантом (18 м в длину) и крупнейшим хищным млекопитающим, обитавшим в океанах. Кстати, его название происходит от слова «левиафан» и указывает на Германа Мелвилла, автора книги «Моби Дик». Вероятно, этот монстр мог бы съесть мегалодона, если бы захотел.
Затем, когда в течение плиоцена температура мирового океана стала понижаться (особенно после того, как 4–3 млн лет назад сформировались полярные ледники), зубы мегалодона попадаются всё реже. В породах позднего плиоцена они встречаются исключительно редко. Это свидетельствует о том, что мегалодоны не выдержали конкуренции со сверхкрупными хищными китами в условиях похолодания океана. Как бы то ни было, они вымерли.
Документальная фантастика
Когда в 1980-е годы активно распространялось кабельное телевидение, появилось много нишевых каналов, каждый из которых был рассчитан на специфическую аудиторию: например, каналы по гольфу, полицейским детективам или истории. К сожалению, из-за отсутствия регулирования на рынке телепередач в конце 1980-х годов образовались многочисленные коммерческие каналы, конкурировавшие друг с другом за позиции в рейтингах, их исходное назначение вскоре было забыто. Так, канал Discovery (изначально созданный для трансляции научно-популярной документалистики) в настоящее время заполонили сомнительные сериалы на паранормальные и псевдонаучные темы. Естественно, отказ от исходной научно-образовательной миссии не замедлил сказаться и на научной документалистике, которая там осталась.
В своё время хитом на Discovery была передача «Shark Week» («Неделя с акулами»). Тогда она рассказывала исключительно о настоящих акулах и их биологии. Но в 2013 году на канале вышел нелепый псевдодокументальный фильм под названием «Megalodon: The Monster Shark Lives» («Мегалодон. Чудовищная акула жива»), за которым в 2014 году последовал «Megalodon: The New Evidence» («Мегалодон. Новые доказательства»). В обеих лентах присутствовали неясные и жутковатые кадры, плохо освещённые снимки, а также реконструкции, выполненные с помощью компьютерной графики. Выступали актёры, выдававшие себя за учёных. Наконец, излагались многочисленные «воспоминания» некой семьи, столкнувшейся с мегалодоном во время морского путешествия.
Лишь в заключительных титрах, занимавших в обоих фильмах несколько секунд, появлялось уведомление о том, что фильм полностью выдуман. Выступая на публике, его продюсеры без устали намекали, что «истина где-то рядом». Естественно, большинство зрителей, успевавшие посмотреть только часть ленты либо не видевшие финального уведомления, воспринимали всё серьёзно. Поэтому многие по-прежнему верят, что мегалодоны могут таиться в глубинах.
Учёные и научные журналисты возмутились, жёстко отреагировав на трансляцию этой «документальной чуши». Но, вероятно, это не помогло: фильм «Megalodon: The Monster Shark Lives» собрал 4,8 млн просмотров и стал одним из самых популярных фильмов в истории Сети. Можете не сомневаться, что канал Discovery будет ежегодно выдавать подобные ленты для «Shark Week». В конце концов, он не является официальным каналом уровня BBC и не обязан обеспечивать правдивость сюжетов. При отсутствии регулирования мотивация канала заключается в привлечении зрителей и поддержке рейтингов для рекламодателей, а в таком случае все средства хороши.
Лучше один раз увидеть!
С тех пор как умер Боб Эрнст, организация «Эрнст Куорриз» (http://www.sharktoothhillproperty.com/) допускает к костным отложениям большинство групп, не имеющих коммерческих целей (за символический взнос, который действительно того стоит). Музей естественной истории и науки Буэна-Виста (http://www.sharktoothhill.org/index.cfm?fuseaction=page&page_id=11) обеспечивает своим сотрудникам привилегии при раскопках.
Окаменелости мегалодона и реконструкции этой акулы экспонируются во многих музеях. Челюсти мегалодона висят под потолком в Зале происхождения позвоночных в Американском музее естественной истории (штат Нью-Йорк). Там же представлены многие другие ископаемые акулы. В Музее естественной истории и науки Буэна-Виста (Бейкерсфилд, штат Калифорния) представлена крупнейшая коллекция окаменелостей с холма Шарктус, в том числе челюсти мегалодона. Реконструированный скелет длиной 10,5 м и много зубов мегалодона вы найдёте в Калвертовском морском музее на острове Соломонс (штат Мэриленд). Во Флоридском музее естественной истории (город Гейнсвилл) есть поразительная экспозиция: несколько пар реконструированных челюстей мегалодона разного размера. Модель мегалодона в натуральную величину висит под потолком галереи в Музее естественной истории Сан-Диего, где также собраны интересные образцы зубов.
Дополнительная литература
Compagno, Leonard, Mark Dando, and Sarah Fowler. Sharks of the World. — Princeton, N. J.: Princeton University Press, 2005.
Ellis, Richard. Big Fish. — New York: Abrams, 2009.
Ellis, Richard. The Book of Sharks. — New York: Knopf, 1989.
Ellis, Richard. Monsters of the Sea: The History, Natural History, and Mythology of the Oceans’ Most Fantastic Creatures. — New York: Knopf, 1994.
Ellis, Richard, and John E. McCosker. Great White Shark. — Stanford, Calif.: Stanford University Press, 1995.
Klimley, A. Peter, and David G. Ainley, eds. Great White Sharks: The Biology of Carcharodon carcharias. — San Diego: Academic Press, 1998.
Long, John A. The Rise of Fishes: 500 Million Years of Evolution. — Baltimore: Johns Hopkins University Press, 2010.
Maisey, John G. Discovering Fossil Fishes. — New York: Holt, 1996.
Renz, Mark. Megalodon: Hunting the Hunter. — New York: Paleo Press, 2002.
10. Рыба, вышедшая из воды
Происхождение земноводных
Тиктаалик
Но зачем вообще жить в подобной среде? Что могло заставить рыбу покинуть толщу воды и поселиться на мелководье? Подумайте вот о чём: едва ли не все рыбы, жившие в реках в те далёкие времена, 375 миллионов лет назад, были хищниками того или иного рода. Некоторые из них достигали в длину пяти метров — вдвое больше, чем самый крупный тиктаалик. Самая обычная рыба, остатки которой встречаются рядом с остатками тиктаалика, превышала два метра в длину и имела голову размером с баскетбольный мяч. Её зубы были размером с костыли, которыми закрепляют железнодорожные рельсы. Хотелось бы вам поплавать в этой древней реке?
Нил Шубин. «Внутренняя рыба»
Из воды на сушу
С тех самых пор, как в 1859 году Чарльз Дарвин опубликовал свою книгу «Происхождение видов», учёные искали окаменелости, которые помогли бы понять, как произошёл один из ключевых витков эволюции: рыба выползла на сушу и превратилась в наземное существо. Разумеется, целый класс животных, земноводные (амфибии), так и живёт на переходном этапе. Некоторые из них почти всю жизнь проводят в воде и редко выбираются на сушу. Другие никогда не залезают в воду, им приходится жить во влажной среде. Многие ведут «двойную жизнь».
Ещё до публикации книги Дарвина некоторые учёные отмечали общие черты у амфибий и двоякодышащих рыб. Такие животные обладают некоторыми чертами земноводных (в частности, имеют лёгкие), но остаются рыбами с плавниками. Правда, плавники двоякодышащих и прочих лопастепёрых рыб состоят из тех же костей, что и конечности амфибий. Но и эта грань не была достаточно чёткой. Американский чешуйчатник (
Lepidosiren paradoxa) настолько специализирован, что имеет лишь крошечные плавники-ленточки и плавает как угорь. Когда его открыли в 1837 году, появилось предположение, что это выродившееся земноводное. Почти аналогичная ситуация возникла, когда Ричард Оуэн описал в 1839 году африканского протоптера (
Protopterus). Убеждённый противник эволюционизма, Оуэн проигнорировал явное анатомическое сходство двоякодышащих и земноводных, подчеркнув странные специализированные черты животного, например крошечные плавники-ленточки. Лишь после того, как в 1870 году был открыт австралийский рогозуб (
Neoceratodus forsteri), появилась возможность убедиться, что некоторые современные двоякодышащие рыбы имеют крепкие лопастевидные плавники ровно с такими же костями, как и в конечностях амфибий. Это наблюдение подкреплялось всё новыми ископаемыми находками примитивных двоякодышащих, подтверждавшими, что у большинства двоякодышащих имелись многочисленные «земноводные» черты, а не странные «специализации», как у протоптера и чешуйчатника.
Всё-таки в палеонтологической летописи между двоякодышащими рыбами и древнейшими амфибиями зияла жуткая пропасть. В 1881 году Джозеф Ф. Уайтивз описал
Eusthenopteron foordi — вероятно, одно из лучших ископаемых переходных звеньев. К сожалению, это описание заняло всего два абзаца; там не было иллюстраций и не прояснялось, какими «земноводными» признаками обладала эта рыба. Эустеноптерон был крупной (до 1,8 м) лопастепёрой рыбой и напоминал амфибий сильнее, чем любой сохранившийся до наших дней представитель двоякодышащих или целакант (рис. 10.1). Известны сотни красивых экземпляров этой рыбы, добытые в знаменитом районе близ Мигуаша, на берегу залива Скауменак в канадском Квебеке. Хотя тело эустеноптерона всё ещё напоминало рыбье, в его плавниках имелись все кости, входящие в состав верхних и нижних конечностей амфибий. Строение костей черепа тоже было как у потенциального предка амфибий.
 Рис. 10.1.
Рис. 10.1. Сравнение элементов скелета ихтиостеги и эустеноптерона (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 10.5)
Впоследствии открыли окаменелости, свидетельствующие, что двоякодышащие и прочие лопастепёрые рыбы во множестве обитали в позднем девоне (385–355 млн лет назад). К началу карбона (355–331 млн лет назад) появился ряд настоящих амфибий (в XIX веке их называли «стегоцефалы» или «лабиринтодонты»; в настоящее время эти термины устарели). Однако их окаменелости более распространены в породах позднего карбона. Итак, где ископаемые переходные звенья? Было найдено множество позднедевонских окаменелостей морских рыб, но лишь немногие из них могли оказаться пресноводными (среди которых предполагалось найти ископаемое существо, расположенное на условной «грани» между рыбами и земноводными).
Прорыв случился благодаря стечению обстоятельств и политическим интересам. В 1920-е годы между Норвегией и Данией возник территориальный спор за Восточную Гренландию. Впоследствии датское правительство и фонд, учреждённый знаменитой датской пивоваренной компанией Carlsberg, профинансировали трёхлетнюю экспедицию к Восточной Гренландии, включавшую три высадки на огромный остров — летом 1931, 1932 и 1933 года. Участники экспедиции рассчитывали провести достаточное количество исследований, чтобы обосновать датские территориальные претензии на Восточную Гренландию, поскольку Норвегия никаких изысканий там не вела. Во главе экспедиции стоял знаменитый датский геолог и исследователь Лауге Кох; также в её состав входила по-настоящему звёздная плеяда датских и шведских геологов, географов, археологов, зоологов и ботаников.
Среди учёных, привлечённых к работам в Восточной Гренландии, был и Гуннар Сёве-Сёдерберг, шведский палеонтолог и геолог. Он получил образование в Уппсальском университете и впоследствии стал там профессором геологии. На момент первой экспедиции в Гренландию ему исполнился всего 21 год. Вскоре Сёве-Сёдерберг обнаружил окаменелости удивительных существ, которых назвал ихтиостегами и акантостегами, а также более примитивных лопастепёрых рыб (в частности,
остеолеписа, очень похожего на эустеноптерона плюс многих двоякодышащих рыб). Очевидно, все эти животные плавали в пресных или солоноватых водах, когда Восточная Гренландия располагалась в тропиках, а девонская эпоха рыб (см. главу 8) подходила к концу. В течение 1920-х и в начале 1930-х годов Сёве-Сёдерберг опубликовал краткие описания ископаемых, планируя позже заняться более детальным анализом. Однако ему было не суждено этого сделать: в 1948 году учёный умер от туберкулёза в возрасте 38 лет.
Сёве-Сёдерберг был представителем старинной шведской научной школы, занятой изучением древних ископаемых рыб. Поскольку шведы снаряжали полярные экспедиции в Гренландию, на Шпицберген и в другие места, повсюду открывали множество ископаемых рыб, эта дисциплина вскоре стала их специализацией. Основателем Стокгольмской палеонтологической школы, которая базировалась преимущественно в стенах Шведского музея естественной истории, был прославленный Эрик Стенсё, известный детальными описаниями девонских панцирных бесчелюстных рыб. В его распоряжении находилось такое множество качественных экземпляров, что некоторые из них он мог резать на тонкие слои (технология серийных срезов) и рассматривать детали нервов, кровеносных сосудов и прочую внутреннюю анатомию, которую при описании
ископаемых рыб обычно не затрагивают. Сегодня рентгеновская компьютерная томография высокого разрешения позволяет палеонтологам сканировать цельную окаменелость, не разрезая её послойно и не разрушая в каких-либо иных целях.
После смерти Сёве-Сёдерберга его гренландские окаменелости исследовал Эрик Ярвик, ученик Стенсё. Он сопровождал Сёве-Сёдерберга в его последних экспедициях в Гренландию, а затем вернулся туда, чтобы собрать новые окаменелости. Ярвик работал аккуратно и методично, никогда не спешил публиковать результаты исследований; годами препарировал срезы эустеноптерона, изучая детали внутренней анатомии его черепа. Он
50 лет работал над окаменелостями ихтиостеги, найденными Сёве-Сёдербергом, пока наконец не подготовил основательную публикацию в 1996 году, когда ему было уже 89 лет. В палеонтологии позвоночных есть легендарные истории об учёных, которые годами корпели над окаменелостями, не обнародовав ни строчки из своих исследований, Ярвик же получает пальму первенства как самый неторопливый палеонтолог в мире. Хотя его исследования были важны, а проделанная описательная работа впечатляет, он придерживался довольно странных взглядов на разные группы окаменелостей, считавшихся в палеонтологических кругах неубедительными. Ярвик умер в 1998 году, в почтенном 91-летнем возрасте.
Поскольку подробное описание ихтиостеги, подготовленное Ярвиком, было опубликовано лишь в 1996 году, первые реконструкции этих ископаемых, выполненные ещё Сёве-Сёдербергом, оставались единственными хорошо документированными материалами по этим ихтиофибиям с 1920-х вплоть до 1980-х годов. Таким образом, ихтиостега стала архетипическим ископаемым переходным звеном между эустеноптероном и древними амфибиями (см. рис. 10.1). Как и амфибии, ихтиостега имела четыре лапы с пальцами, а не лопастевидные плавники, как её предки. Однако её передние конечности были недостаточно сильны, чтобы много ходить, и новейшие исследования показывают: она могла передвигаться, вероятно, лишь короткими «перебежками», волоча за собой задние конечности, больше напоминавшие плавники. Передние и в особенности задние конечности ихтиостеги были гораздо лучше приспособлены для движения в воде, где животное отталкивалось ими подобно современным тритонам и саламандрам. Ихтиостега обладала крепкими рёбрами с гребнями, которые, вероятно, помогали ей удерживать над водой грудную клетку с лёгкими. Однако они не обеспечивали рёберного дыхания, характерного для большинства земноводных. Другой «земноводной» чертой ихтиостеги была её длинная плоская морда с глазами, направленными вверх, а также короткая черепная коробка. Эустеноптерон обладал более «рыбьим» цилиндрическим черепом, короткой мордой и длинной черепной коробкой. Его глаза смотрели в стороны по бокам головы; ещё у него были большие жаберные крышки. Однако, если не считать конечностей, плечевых и бедренных костей, ихтиостега во всём напоминала рыбу. У неё имелся большой хвостовой плавник и многие «рыбьи» признаки черепа, большие жаберные крышки, слуховой аппарат, приспособленный к воде, и боковая линия (каналы на морде, с помощью которых животное отслеживало движение и направление течений в воде).
В 1980-е годы основным центром исследований ихтиофибий была уже не Швеция, а Кембриджский университет, где сбором новых окаменелостей и повторением работы представителей стокгольмской школы занимались Дженни Клэк, Пер Альберг, Майкл Коутс и др. Вот как рассказывает об этом Клэк:
В 1985-м я стала задумываться об экспедиции в Восточную Гренландию, на это меня подвигнул мой муж Роб. Тем временем я познакомилась с Питером Френдом с геологического факультета, расположенного в Кембридже через дорогу от нас. Питер был руководителем нескольких экспедиций в ту часть Гренландии, которая меня интересовала. Оказалось, у него был студент, Джон Николсон, собравший несколько окаменелостей в рамках работы над своей диссертацией о восточногренландских осадочных породах верхнего девона (он сделал это в 1968–1970 годах). Питер принёс эти образцы, пылившиеся в подвале, а также показал мне блокнот Джона, в котором тот делал заметки о своей экспедиции 1970 года. Ремарка Джона о том, что на горе Стенсё (800 м) в изобилии встречаются кости ихтиостеги, была поразительной и необычайной. Собранные им окаменелости складывались в единый блок, включавший три частично сохранившихся черепа и фрагменты плечевого пояса, — но принадлежали они не ихтиостеге, а акантостеге — родственному ей виду, тогда ещё малоизвестному. Питер предложил мне связаться со Свендом Бендикс-Альмгреном, заведующим отделом палеонтологии позвоночных в геологическом музее Копенгагена. Датчане по-прежнему снаряжали геологические экспедиции в национальный парк «Восточная Гренландия», где расположены девонские разработки. Поэтому подготовку к экспедиции мне следовало начинать именно со знакомства с Бендикс-Альмгреном. Также Питер посоветовал мне наладить контакт с Нильсом Хенриксеном из Гренландской геологической службы (ГГС). По чистой случайности и невероятному везению ГГС как раз вела проект в том самом месте, куда мне требовалось отправиться, и последний сезон работ там приходился на лето 1987 года. Получив финансирование от Университетского зоологического музея, Фонда Ганса Гадова в Кембридже и Фонда Карлсберг в Копенгагене, мы: я, мой муж Роб, Пер Альберг, который тогда был моим студентом, а также Свенд Бендикс-Альмгрен вместе со своим студентом Биргером Йоргенсоном отправились в полуторамесячную экспедицию под эгидой ГГС в июле–августе 1987 года. Руководствуясь полевыми заметками Джона Николсона, мы наконец-то отыскали то место, откуда он взял образцы акантостеги, а затем и конкретный горизонт на месте залегания, к которому они относились. В сущности, это был крошечный, но изобильный «раскоп» с акантостегами.
Открытие гораздо более полных экземпляров акантостеги стало настоящим прорывом. В 1952 году Ярвик назвал акантостегу, имея в распоряжении скудный и малоизученный материал. Новые окаменелости, собранные Клэк и её группой в конце 1980-х и в 1990-е годы, дали по акантостеге гораздо более полную и информативную картину, особенно в сравнении с исходным материалом по ихтиостеге (рис. 10.2). В целом маленькая акантостега напоминала рыбу значительно сильнее, нежели ихтиостега. В отличие от ихтиостеги, она не могла ползать по суше на своих лапах: у неё не было запястных, локтевых и коленных суставов. Конечности позволяли ей лишь плескаться и преодолевать преграды под водой. Ещё удивительнее, что на передних конечностях акантостеги было целых семь или восемь пальцев, а не пять, как у большинства позвоночных! Акантостега обладала значительно более крупным хвостовым плавником, чем ихтиостега, её рёбра были слишком коротки и не могли поддерживать животное на суше, не позволяли они и дышать вне воды. Однако у акантостеги были некоторые развитые «земноводные» черты: она могла слышать как в воде, так и в воздухе, имела прочные плечевые и бедренные кости, четыре конечности с пальцами, а также шейный сустав, позволявший ей вращать головой. Напротив, рыба не имеет шеи и не может вертеть головой. Чтобы изменить курс или схватить добычу, она делает рывок всей передней частью тела.
 Рис. 10.2.
Рис. 10.2. Сравнение скелетов ихтиостеги (внизу) и акантостеги (вверху) (рисунок предоставлен M. Coates)
Ваша внутренняя рыба
Работа Дженни Клэк оживила интерес к исследованию переходных форм от рыб к амфибиям, и вскоре к ним подключились многие другие палеонтологи. Одним из них был молодой энтузиаст-учёный Нил Шубин. Он получил высшее палеонтологическое образование в Колумбийском университете и Американском музее естественной истории в Нью-Йорке, где я в то время учился в аспирантуре. Там мы и познакомились в 1980 году, после чего стали вместе изучать эволюцию лошади мезогиппуса — это был первый опубликованный исследовательский проект Шубина. Затем Шубин отправился готовить докторскую диссертацию в Гарвард, где изучал механизмы эволюционного развития, определяющие формирование конечностей и пальцев у амфибий. Первым местом работы Нила стал пост преподавателя анатомии на медицинском факультете Пенсильванского университета в Филадельфии, где он сблизился с Тедом Дэшлером из Академии естественных наук. Вместе они исследовали срезы грунта в местах прокладки дорог через девонские красноцветные отложения по всей Пенсильвании, пока не нашли кое-какие фрагментарные окаменелости рыб и ихтиофибий.
Но Шубина интересовала более крупная добыча. Как он рассказывает в своей книге «Внутренняя рыба», они с Дэшлером знали, что придётся искать породы старше 363 млн лет (такие, как в Восточной Гренландии, где были найдены ихтиостега и акантостега), но моложе 390–380 млн лет (из которых добыто большинство лопастепёрых рыб, предков амфибий). Шубин и Дэшлер прогнозировали, что в верхнедевонских пресноводных отложениях должны найтись ископаемые переходные формы, более примитивные, чем акантостега, но более совершенные, чем эустеноптерон. Они могли бы заполнить пробел, захватывавший период с 380 по 363 млн лет назад. Шубин и Дэшлер изучали геологические карты по первому изданию легендарного учебника «Evolution of the Earth» («Эволюция Земли»; 1971), написанного Робертом Х. Доттом-младшим и Роджером Баттеном. Штудируя карту верхнедевонских обнажений, они нашли три перспективных региона: Восточная Пенсильвания (как раз там они и работали), Восточная Гренландия (образцы оттуда уже были добыты датчанами, шведами и группой Клэк) и остров Элсмир в канадской части Арктики (который никто не изучал). Внимательнее изучив опубликованные отчёты геологической службы, они выяснили, что обнажения на Элсмире были именно девонскими, их возраст составлял 380–363 млн лет и породы там были именно такими, в которых хорошо сохраняются окаменелости пресноводных рыб и амфибий. Оказалось, что возраст этих пород — примерно 375 млн лет.
К концу 1990-х Шубин, Дэшлер и их команда собрали все необходимые разрешения и оборудование, а также получили финансирование на припасы и аренду вертолёта, который бы доставлял их на остров, а затем забирал оттуда. Серьёзная экспедиция в такой регион, как Арктика, — не увеселительная прогулка! Исследователям требовался полный комплект арктического снаряжения, в особенности тёплая одежда для защиты от летних морозов и укреплённые палатки, способные выдерживать ураганные ветра, служить тёплым укрытием во время частых бурь. Кроме кирок, лопат и прочих стандартных палеонтологических инструментов, учёные вооружились винтовками, так как приходилось всерьёз опасаться белых медведей.
Начиная с 2000 года, в самой середине каждого лета они делали короткие вылазки на остров Элсмир. В первые несколько лет результаты не радовали: породы оказались морскими, а не пресноводными по происхождению. Наконец, они нашли породу, богатую пресноводными окаменелостями, какую и искали.
В 2000 году было открыто место, позже названное Птичьим карьером, где к 2003 году удалось найти множество фрагментарных рыбьих окаменелостей. В 2004 году учёные углубились на три метра в подповерхностные породы и открыли ископаемое, оправдавшее все тяготы. Шубин с коллегами выбрали для него название тиктаалик, что в переводе с инуктитута, одного из эскимосских языков, значит налим. Лишь спустя ещё два года эти окаменелости как следует препарировали для исследований, были готовы все анализы и описания, после чего о тиктаалике впервые объявили в статье, опубликованной в 2006 году. Описание его задних конечностей увидело свет в 2012-м.
Удалось добыть более 10 особей тиктаалика, от 1 до 3 м в длину (рис. 10.3). Более того, самый полный экземпляр тиктаалика сохранился практически целиком, в нём недостаёт лишь фрагментов хвоста и задних конечностей, но задние конечности хорошо изучены по другим экземплярам. Как и следовало ожидать от существа, жившего на 12 млн лет раньше ихтиостеги и акантостеги, тиктаалик больше напоминает рыбу по многим признакам. В его лопастевидных плавниках есть все элементы, из которых впоследствии развились конечности земноводных, однако вместо пальцев — всё ещё плавниковые лучи. У него рыбья чешуя, одновременно присутствуют (как у большинства ихтиофибий) жабры (узнаваемые по костям жаберной дуги) и лёгкие (распознаваемые по брызгальцам в голове). Нижняя челюсть и нёбо у него тоже рыбьи. Но, в отличие от любых рыб, у него есть земноводные черты: укороченный уплощённый череп с подвижной шеей; бороздки в задней части черепа для расположенных там барабанных перепонок; крепкие рёбра, а также мощные конечности, плечевые и тазовые кости. Плавники тиктаалика, как и у акантостеги, были недостаточно гибкими и сильными, чтобы он мог далеко ползать по земле или ходить, отрывая брюхо от грунта. Плавники же, вероятно, лишь позволяли животному плескаться в воде и поддерживали тиктаалика, так что он мог осмотреться над поверхностью. Подобно многим другим ихтиофибиям (а также многим современным амфибиям, особенно тритонам и саламандрам), тиктаалик, наверное, большую часть времени проводил в воде, охотясь по берегам ручьёв, в которых обитал.

 Рис. 10.3.
Рис. 10.3. Тиктаалик: (
А) скелет; (
Б) реконструкция внешнего вида (иллюстрации предоставлены N. Shubin)
Вот как писал о нём Роберт Холмс в журнале New Scientist:
Спустя пять лет раскопок на острове Элсмир, на крайнем севере территории Нунавут, они наконец нашли сокровище: несколько рыб, сохранившихся так превосходно, что их скелеты были совершенно целы. Изучая эти экземпляры, команда Шубина, к всеобщему восторгу, обнаружила, что перед ними — как раз недостающее переходное звено, которое они искали. «Мы нашли существо, которое как нельзя чётко отделяло рыб от амфибий», — сказал Дэшлер.
Клэк отметила: « Это одна из тех находок, которую можно показать и заявить: “Говорил же вам, она должна быть!” — и вот она перед вами».
Поиск «ещё более переходных» ископаемых продолжался. Но одно стало очевидным: выход из воды на сушу не был резким скачком, каким его представляли биологи на протяжении более ста лет. Достаточно обратить внимание на широчайшую эволюционную радиацию лучепёрых рыб (
Actinopterygii), к которым относятся 99% особей в рыбных садках, на рыбных рынках и в больших аквариумах. Все современные рыбы, за исключением миног, миксин, акул, скатов, двоякодышащих и целакантов, относятся к лучепёрым. У них нет таких крепких костей, как у лопастепёрых рыб; их плавники поддерживаются благодаря длинным и тонким костным или хрящевым стержням.
Лучепёрые рыбы научились по-разному передвигаться на суше, используя свои слабенькие плавники. Например, илистые прыгуны живут отчасти в воде, отчасти на суше, обитая в мелководных заводях и среди корней мангров. На передних плавниках они могут медленно ползать на границе суши и воды (рис. 10.4). Лягушковый клариевый сом — просто напасть на юго-востоке США, так как может ползать по суше от пруда к пруду, чтобы искать пищу или спасаться из высыхающих водоёмов. Рыба-ползун тоже может рыскать по суше в поисках более уютных водоёмов и даже забираться на деревья. Многие рыбы, в частности бычковые и скорпены, приспособились жить в приливных заводях и во время отлива проводят некоторое время на воздухе. Их передние плавники видоизменились, на них можно ползать и перебираться через камни. Другие рыбы, ведущие в основном водный образ жизни, превратили лучи передних плавников в «пальцы»: с их помощью рыба может закапываться в подводный грунт и толкать себя вперёд.
 Рис. 10.4.
Рис. 10.4. Илистый прыгун кормится червями на литорали в Японии (фотография Alpsdake, фонд Wikimedia Commons)
Между разными группами лучепёрых рыб отсутствует близкое родство, поэтому описанные адаптации к жизни на суше развились независимо друг от друга. Разумеется, только из-за серьёзного прессинга и в погоне за большими преимуществами рыбы начали проникать в наземные экосистемы (пусть всего на минуты или часы). Постепенные изменения лопастепёрых рыб, сначала превратившихся в полуводных, а затем — в настоящих наземных животных, не так уж невозможны, как раньше казалось учёным.
Недавно группа учёных под руководством Эмили Стэндэн опубликовала исследование, продемонстрировавшее, насколько легко рыба может выйти из воды. Эксперимент провели на очень слаборазвитой костной рыбе — многопёре (
Polypterus) из Африки, которая находится в отдалённом родстве с такими примитивными лучепёрыми рыбами, как осетровые и веслоносовые. Плавники многопёра сильно напоминают плавники древнейших лопастепёрых рыб, он практически является переходным звеном между лопастепёрыми и лучепёрыми. Исследователи выращивали многопёров на суше, а не в их обычной водной среде обитания (эти рыбы хорошо дышат воздухом).
Действительно, спустя несколько поколений потомки этих рыб стали более крепкими и лучше приспособились ползать по земле благодаря так называемой адаптационной гибкости, позволяющей животным видоизменяться на эмбриональном этапе развития и приспосабливаться к новым условиям. Как указывает Стэндэн, адаптационная гибкость позволяет не только ответить на вопрос, почему столь многие лучепёрые рыбы научились ползать по земле, но и объясняет механизмы, благодаря которым это же удалось лопастепёрым рыбам.
Итак, у нас есть непрерывная последовательность ихтиофибий — от существ, которые были настоящими рыбами (например, лопастепёрые), до тиктаалика и акантостеги и далее до других животных, которые ещё сильнее напоминали амфибий (см. рис. 10.5). Всем, кому кажется невероятным, что рыба могла выползти из воды и стать сухопутным животным, достаточно взглянуть на эти невероятные окаменелости — и всё понять.
 Рис. 10.5.
Рис. 10.5. Происхождение земноводных от рыб (рис. Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 10.6)
Лучше один раз увидеть!
Насколько мне известно, немногочисленные окаменелости ихтиостеги и акантостеги экспонируются только в Зоологическом университетском музее Кембриджского университета и Шведском музее естественной истории, расположенном в Стокгольме.
В некоторых музеях США можно увидеть отпечатки скелета и реконструкции тиктаалика. В частности, это Академия естественных наук Дрексельского университета (Филадельфия), Музей естественной истории под открытым небом (Чикаго), Музей сравнительной зоологии Гарвардского университета (Кембридж, штат Массачусетс) и Музей естественной истории и науки Цинциннати. Одни из лучших окаменелостей лопастепёрых рыб и древних амфибий находятся в Американском музее естественной истории в Нью-Йорке.
Дополнительная литература
Шубин Н. Внутренняя рыба: история человеческого тела с древнейших времён до наших дней. — М.: Астрель; Corpus, 2010.
Clack, Jennifer A. Gaining Ground: The Origin and Early Evolution of Tetrapods. — Bloomington: Indiana University Press, 2002.
Daeschler, Edward B., Neil H. Shubin, and Farish A. Jenkins Jr. A Devonian Tetrapod-like Fish and the Evolution of the Tetrapod Body Plan //
Nature. — 2006. — April 6. — P. 757–763.
Long, John A. The Rise of Fishes: 500 Million Years of Evolution. — Baltimore: Johns Hopkins University Press, 2010.
Maisey, John G. Discovering Fossil Fishes. — New York: Holt, 1996.
Moy-Thomas, J. A., and R. S. Miles. Palaeozoic Fishes. — Philadelphia: Saunders, 1971.
Shubin, Neil H., Edward B. Daeschler, and Farish A. Jenkins Jr. The Pectoral Fin of Tiktaalik roseae and the Origin of the Tetrapod Limb //
Nature. — 2006. — April 6. — P. 764–771.
Zimmer, Carl. At the Water’s Edge: Macroevolution and the Transformation of Life. — New York: Free Press, 1998.
11. «Лягомандра»
Происхождение лягушек
Геробатрахус
Теории приходят и уходят. Лягушка остаётся.
Жан Ростан. «Тревоги биолога»
«Человек — свидетель потопа»
В начале XVIII века учёные ещё придерживались полярных мнений о происхождении окаменелостей и предлагали многочисленные объяснения присутствия этих странных объектов в горных породах. Латинское слово
fossilis буквально переводится как ископаемое. То есть что угодно, извлечённое из-под земли, в том числе кристаллы, самородки и другие небиологические объекты изначально приравнивались к окаменелостям. Некоторые учёные полагали, что окаменелости — творения дьявола, заложенные в скалах, чтобы смутить верующих и посеять сомнения. Другие утверждали, что они могли вырасти в камне под действием таинственных
формообразующих сил (
vis plastica) или что какие-то существа заползли в скальные трещины, были там раздавлены и погибли, а их скелеты замурованы. И очень немногие связывали фоссилизированные останки двустворчатых и одностворчатых моллюсков с их современными потомками.
Многие окаменелости на тот момент были просто неузнаваемы, поскольку не походили ни на одно современное существо. Считалось, что странные треугольные объекты, так называемые каменные языки (
glossopetrae), упали с неба и обладают мистическими свойствами, в частности могут излечивать от укусов змей и обезвреживать яды. Но в 1669 году датский врач Нильс Стенсен (более известный под латинизированным именем Николай Стенон) заметил «каменные языки» во рту акулы и понял, что это зубы. Большинство людей считали аммониты останками окаменевших змей. Ведь наутилус помпилиус, или обыкновенный кораблик, был открыт только в начале XIX века. Считалось, что членики криноидей — это звёзды, упавшие с неба.
В частности, на представления об окаменелостях по-прежнему влияла Библия. Так, в 1726 году швейцарский натуралист Иоганн Шейхцер описал одну окаменелость как «костный скелет одного из тех нечестивцев-грешников, что были свидетелями потопа». Это был большой скелет, от головы до тазовых костей длиной около метра, с черепом, верхними конечностями и хребтом. Причём его нашли в горах и, следовательно, он должен был принадлежать человеку, погибшему во время Всемирного потопа. Шейхцер назвал его
Homo diluvii testis («Человек — свидетель потопа») (см. рис. 11.1). Но в 1758 году Иоганн Геснер, один из первопроходцев-натуралистов, оспорил эту гипотезу, предположив, что окаменелость принадлежит каракатице. Затем в 1777 году Петрус Кампер доказывал, что это ящерица. В 1802 году Мартин ван Маур купил данный экземпляр для Музея Тейлора в Харлеме, где он находится до сих пор. В 1836 году окаменелость получила официальное название
Andrias scheuchzeri, которое можно перевести как шейхцеровский человек.
 Рис. 11.1.
Рис. 11.1. «Человек — свидетель потопа» Иоганна Шейхцера — экспонат Музея Тейлора в Харлеме (Нидерланды) (из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 1.4)
Эта ошибка сохранялась в течение почти ста лет после того, как Шейхцер описал «человека». Когда Наполеон аннексировал Нидерланды, образец отправился в Париж, где его стал изучать знаменитый барон Жорж Кювье — основоположник палеонтологии позвоночных и сравнительной анатомии. Он исследовал окаменелый скелет и заметил многие детали, которые не удалось увидеть поначалу, особенно в верхних конечностях. Кроме того, всю свою профессиональную карьеру Кювье изучал сравнительную анатомию, поэтому сразу понял, что перед ним не человеческий скелет. Сравнив его с некоторыми аналогами, он пришёл к выводу, что это даже не примат и не млекопитающее, а гигантская саламандра!
Такие гигантские саламандры существуют до сих пор. Два вида, обитающие в Японии и Китае, даже крупнее окаменелости Шейхцера (рис. 11.2). Китайская исполинская саламандра достигает в длину почти 2 м и может весить до 36 кг. Она относится к тому же роду, что и находка Шейхцера, но называется
Andrias davidanus и обитает в порожистых горных реках и озёрах с чистой водой. Обычно встречается в лесистых регионах, а также на высоте от 100 до 1500 м. Японский вид называется
Andrias japonica. Это животное немного мельче китайской исполинской саламандры и живёт примерно в таких же местах. Оба вида относятся к исчезаемым, так как человек разрушает их среду обитания, а столь крупным водным животным для выживания требуется большая территория. Кроме того, за ними охотятся, поскольку эти саламандры используются в традиционной китайской медицине, из-за которой на грани вымирания уже оказались носороги, тигры, панголины и многие другие животные.
 Рис. 11.2.
Рис. 11.2. Исполинская китайская саламандра (фотография предоставлена Luke Linhoff)
Жизнь по обе стороны
В главе 10 мы узнали, как земноводные произошли от лопастепёрых рыб в позднем девоне. Но как они разделились на знакомые нам современные группы амфибий — в частности, лягушек, жаб и саламандр? Опять же, в палеонтологической летописи сохранились отдельные замечательные экземпляры, демонстрирующие этапы данного эволюционного процесса.
Слово «амфибия» происходит от греческого
amphibion, означающего «живущий по обе стороны» — то есть и в воде, и на суше. Такая двойная жизнь — одна из характерных черт амфибий. Большинство из них могут припеваючи жить и там, и там, поскольку в обоих случаях получают достаточно влаги. Пустынные жабы приспособились существовать в мире почти без воды и кое-как выживают под землёй, где прохладно и сыро. Однако большинству амфибий по-прежнему требуются влажные места, чтобы откладывать яйца и замыкать жизненный цикл. Правда, некоторые амфибии живородящие, яиц не откладывают.
Современные амфибии исключительно разнообразны, известно свыше 5700 их видов. Более 4700 — это лягушки и жабы, всего 655 — саламандры и тритоны. Кроме того, около 200 видов относятся к третьей группе амфибий — безногие. Последние обитают преимущественно под землёй, во влажных тропических почвах Южной Америки, Африки и Азии. Некоторые имеют крошечные глаза, позволяющие отличать свет от тьмы, у других глаза расположены на кончиках чувствительных щупалец, но большинство безногих слепы. Неспециалисту они больше всего напоминают гигантских земляных червей.
Размеры амфибий очень разнообразны: от крошечной новогвинейской лягушки
Paedophryne amanuensis длиной всего 7,7 мм до исполинской китайской саламандры. Тритоны и саламандры обладают таким же продолговатым телом, длинным хвостом и простыми передними конечностями, как большинство примитивных амфибий (в частности, тиктаалик, ихтиостега и акантостега, о которых мы говорили в главе 10).
 Рис. 11.3.
Рис. 11.3. Сравнение скелетов триадобатрахуса (слева) и современной лягушки (справа). Хотя на первый взгляд они схожи, триадобатрахус был гораздо примитивнее любых современных лягушек, имел много предкрестцовых позвонков, небольшие неразвитые бёдра (а не продолговатые, как у лягушек), короткие передние и задние конечности, не позволявшие прыгать, немного удлинённый хвост, а также гораздо более примитивный череп (рисунок Mary P. Williams)
Из всех ныне живущих амфибий лягушки достигли самой впечатляющей изменчивости формы тела по сравнению с той, что была у их общего предка. Все, кому доводилось препарировать лягушку на уроках биологии, знают, что строение лягушачьего тела поистине уникально. Пусть лягушки и жабы бесхвостые, их личинки (головастики) появляются на свет с хвостом, который укорачивается по мере того как особь взрослеет. Голова у лягушек короткая, с тупой широкой мордой, позволяющей широко раскрывать рот и захватывать пищу (часто с помощью длинного липкого языка). Благодаря очень длинным и сильным задним лапкам лягушка может совершать огромные прыжки (чтобы ловить добычу или спасаться от хищников), а также отлично плавать. Её туловищный отдел невелик, включает маленькие толстые рёбра и очень продолговатые тазовые кости, поддерживающие мышцы задних конечностей. Поскольку рёбра лягушки не участвуют в дыхании, животное пользуется надувным глоточным мешком, который помогает ему делать вдох и выдох (а также издавать разные звуки). Лягушки бывают очень разных размеров: от крошечной новогвинейской лягушки до лягушки-голиафа длиной более 300 мм и весом до 3 кг. Эта лягушка такая огромная, что питается не только насекомыми, но и птицами, и мелкими млекопитающими.
Как ни грандиозна лягушка-голиаф, в 1993 году группа учёных, исследовавших позднемеловые породы Мадагаскара, нашла окаменелость ещё более крупного существа. Потратив 15 лет на сборку скелета из мелких фрагментов (труднее всего было сложить череп из 75 кусочков), они опубликовали описание этой лягушки в 2008 году. Её назвали
Beelzbufo ampinga (дьявольская жаба). Родовое название состоит из имени Вельзевул (прозвище дьявола означает «Повелитель мух») и слова
bufo — общего названия жаб. Видовое имя переводится с малагасийского как щит. Это животное относилось к роду рогатых лягушек, ныне обитающих в Южной Америке. Таким образом, некогда рогатые лягушки были распространены по всей Гондване, к которой относилась большая часть суши современного Южного полушария. Примечателен размер дьявольской жабы. На примере почти полного скелета можно заключить, что она имела длину 40 см и весила 4 кг — на треть крупнее лягушки-голиафа! У неё была очень большая голова и широченный рот — такой огромный, что она, пожалуй, могла глотать детёнышей динозавров (в те времена Мадагаскар был населён ящерами).
Сокровища красноцветных отложений
Мы коротко рассмотрели биоразнообразие современных амфибий и их размеры. А что насчёт их ископаемых предков? После ихтиофибий (см. главу 10) произошёл гигантский эволюционный взрыв разнообразия амфибий, который пришёлся на карбон (350–300 млн лет назад) и пермь (300–250 млн лет назад). Большинство из них относилось к трём ныне исчезнувшим группам, которые в своё время были самыми крупными животными, господствовавшими на суше, пока в начале перми не наступила эпоха рептилий.
Пожалуй, лучшим местом для сбора раннепермских амфибий и сосуществовавших с ними сухопутных животных являются красноцветные отложения северного Техаса, особенно в районе Уичита-Фолс и Сеймур, а также территория Оклахомы на границе с Техасом. Эти невероятные залежи окаменелостей в 1877 году открыл Эдвард Дринкер Коп, один из пионеров палеонтологии. На лошади и в фургоне, заручившись поддержкой всего пары местных добровольцев, он нашёл районы, буквально усыпанные фрагментами костей, черепами и скелетами. Полный фургон образцов Коп собрал всего за несколько дней, став первым из множества американских палеонтологов, работавших на этих богатых залежах, и отправил находки в Филадельфию, где собирался их изучать.
Практически все учёные, писавшие об эволюции ранних рептилий и амфибий, собирали образцы в красноцветных отложениях Техаса; в том числе маститые исследователи, имена которых знает каждый палеонтолог. Это и Сэмюэль Уэнделл Уиллистон, работавший в Канзасском университете в 1890-е годы и в Университете Чикаго вплоть до своей смерти в 1918 году; и Альфред Ш. Ромер, работавший в университете Чикаго в 1920-е годы и в Гарварде (до 1970-х); и Эверетт «Оул» Олсен, работавший сначала в университете Чикаго, а затем в Калифорнийском университете Лос-Анджелеса.
Сбор образцов в этих краях — не пикник. Летом пекло и бушуют песчаные бури, всё заметающие красной пылью: еду, питьевую воду, снаряжение, глаза и другие чувствительные части тела. Грунтовые воды здесь горячи, как чай, и дурны на вкус, полны розоватой грязи и щелочей. Если пить слишком много такой жидкости, образуются камни в почках. Найдя хорошее место, собиратели должны глубоко копать и хорошенько прятаться, чтобы не перегреться на солнце и не надышаться пыли.
Но дело того стоит! Самое распространённое ископаемое в красноцветных отложениях — хищник диметродон, размером примерно с тигра, имевший гребень на спине. Он хорошо известен по пластмассовым наборам динозавров для детей и из детских книг на ту же тему (см. главу 19). Однако диметродон был не динозавром, а одним из первых представителей эволюционной линии, от которой произошли млекопитающие. Эта линия называется синапсиды, или протомлекопитающие. В своё время их называли зверообразными рептилиями, хотя синапсиды рептилиями не являлись. Большинство экземпляров достигали 2–4 м в длину, весили до 270 кг и имели спинные гребни высотой до 1,2 м. Диметродон, вероятно, был доминирующим хищником своего времени, охотился на более мелких синапсид с гребнями на спине, в частности на растительноядных эдафозавров, а также на всевозможных мелких настоящих рептилий, в том числе на капторинуса (размером с ящерицу), близкого родственника черепах.
Но синапсиды и останки рептилий в техасских красноцветных отложениях — лишь крошечная часть большой истории. Хотя диметродон и был царём природы в ранней перми, именно тогда амфибии достигли максимального разнообразия и наибольших размеров. Многие из них были господствующими хищниками, конкурировавшими за пищу в суровом мире.
Когда амфибии правили миром
Самой многочисленной и эффектной из трёх групп позднепалеозойских амфибий были темноспондильные (ранее именовавшиеся лабиринтодонтами) . Большинство из них походили на толстых крокодилов с длинным торсом и хвостом, а также мощными и широко расставленными конечностями. Однако, в отличие от крокодилов, они имели гигантские уплощённые черепа с глазницами, направленными вверх. Огромная пасть лабиринтодонта была усажена острыми коническими зубами. Голова некоторых специализированных темноспондильных, так называемых архегозавров, на первый взгляд напоминала крокодилью (у архегозавров была длинная узкая морда). Одним из архегозавров являлся прионозух, обнаруженный в среднепермской формации Педро до Фого в Бразилии (её возраст — 270 млн лет). Прионозух жил в лагунах и реках, походил на крокодила не только формой тела, но и длинной узкой мордой, приспособленной для охоты на рыбу и другую водную живность. Именно такой образ жизни ведёт гавиал. Если, как полагают некоторые учёные, прионозух действительно достигал 9 м в длину, он был крупнейшим земноводным, когда-либо жившим на Земле и, в частности, превосходил по размерам любых ныне живущих крокодилов. Другие учёные считают, что оценки его тела и хвоста преувеличены, и длина прионозуха вполне могла не превышать 5 м.
Древнейшие темноспондильные вырастали всего до метра, но к пермскому периоду они превратились в одних из самых крупных сухопутных животных, каких знала наша планета. Одна из наиболее распространённых окаменелостей в раннепермских красноцветных отложениях Техаса —
эриопс, крупный лабиринтодонт, известный по множеству полных скелетов (рис. 11.4,
А). У него было большое тело длиной до 2 м, с крепким хвостом и сильными лапами. Черепа некоторых крупных особей достигала 60 см в длину! Эриопс был одним из крупнейших сухопутных животных раннепермской эпохи, мог охотиться как в воде, так и на суше. Чуть более примитивный
эдопс, также найденный в раннепермских красноцветных отложениях Техаса, обладал ещё более длинным черепом, а следовательно, был крупнее эриопса.


 Рис. 11.4.
Рис. 11.4. Древние амфибии: (
А) эриопс, представитель темноспондильных; (
Б) реконструкция диплокаулуса, представителя тонкопозвонковых; (
В) сеймурия, представитель антракозавров (иллюстрации (
А) и (
В) — из фонда Wikimedia Commons; иллюстрация (
Б) предоставлена Nobumichi Tamura)
К концу пермского периода крупные сухопутные лабиринтодонты перешли к полностью водному образу жизни, возможно, отступив под напором более крупных хищных синапсид, завоёвывавших сушу в то время. Темноспондильные смогли пережить крупнейшее массовое вымирание в истории Земли в конце пермского периода (250 млн лет назад). Они жили и в триасе (250–200 млн лет назад), их останки часто встречаются в отложениях на месте болот и озёр, например на территории национального парка Petrified Forest («Окаменелый лес») в штате Аризона. У этих последних лабиринтодонтов были слабые лапы, вероятно, непригодные для ходьбы по суше; уплощённые головы с глазами, смотревшими только вверх, и массивные жирные тела, приспособленные к жизни на мелководье и охоте на водоплавающую дичь.
Вторая вымершая группа амфибий — тонкопозвонковые (лепоспондилы), существовавшие с начала карбона до начала перми, но только в Европе и Северной Америке. Большинство из них были мельче темноспондильных, живших в то же время. Тонкопозвонковые имели длинное тело, как у саламандры, и маленькие лапки. Вероятно, эти животные вели преимущественно водный образ жизни. Некоторые из них, например аистоподы, полностью утратили конечности и напоминали водоплавающих змей. Другие, в частности микрозавры, больше напоминали ящериц, имели глубокие черепа и сильные лапы. Самым известным представителем тонкопозвонковых является диплокаулус (рис. 11.4,
Б), который выглядел весьма странно. Этот представитель тонкопозвонковых, хорошо известный по тем же красноцветным техасским отложениям, достигал метра в длину, а своим приземистым телом напоминал саламандру. Он был почти полностью покрыт крепким панцирем, имел широкие мощные челюсти.
Самой удивительной частью тела диплокаулуса была его голова, которая по форме напоминала бумеранг. Череп был уплощённым, и по обе стороны от него отходили своеобразные «рога». Глазницы направлены вверх. Назначение этих странных «рогов» до сих пор остаётся предметом для дискуссий. Некоторые считают, что они могли выполнять роль подводных крыльев, позволяя диплокаулусу изящно плавать, двигаясь вверх-вниз; при этом голова-бумеранг обеспечивала подъёмную силу. Однако комплекция животного была слишком слабой, и вряд ли у него имелись крепкие кости, необходимые для поддержки сильных плавательных мышц. Другие считают, что при такой форме головы хищникам было сложно съесть диплокаулуса, так как из-за рогов эту голову не удавалось заглотить даже самым крупным раннепермским плотоядным. Глаза, направленные вверх, позволяют предположить, что диплокаулус был хищником-засадником, лежал на дне реки или пруда, а затем бросался вперёд и вверх, хватая добычу мощными челюстями, возможно, предварительно оглушая её ударом «рогов». Наиболее сильная гипотеза такова: возможно, «рога» аналогичны рогам антилоп и оленей, самцы которых пользуются ими для демонстрации силы и привлекательности, когда ищут самок для спаривания. Тот факт, что «рога» диплокаулуса росли (можно проследить по окаменелостям, у некоторых «рога» маленькие), а также наличие среди диплокаулусов самцов с увесистыми рогами и самок с более мелкими рожками делает эту гипотезу наиболее вероятной.
Третья группа вымерших амфибий — так называемые антракозавры, своеобразная мусорная корзина для всех сравнительно высокоразвитых земноводных, чья эволюционная линия впоследствии приведёт к появлению рептилий (рис. 11.4,
В). Техасские красноцветные отложения изобилуют замечательными экземплярами, например окаменелостями трёхметрового диадектеса (размером с бегемота), а также сеймурий, которые очень напоминали рептилий. Эти животные названы в честь города Сеймур, расположенного в центре техасских красноцветных отложений.
В поисках «Лягомандры»
Крупнейшие участники «техасской палеонтологической лихорадки», которая развернулась в середине XX века (в частности, Ромер и Ольсон) уже умерли, но их ученики продолжают наведываться туда и собирать важные окаменелости. Среди важнейших представителей этого второго поколения были Роберт Кэрролл из музея Редпасс в Монреале (учившийся у Ромера в Гарварде), Роберт Рейш (первый ученик Кэрролла, в настоящее время работающий в Университете Торонто), покойный Николас Хоттон из Смитсоновского института (учившийся у Ромера и Ольсона в Чикаго), покойный Питер Вон (учился у Ромера, а затем вместе с Ольсоном воспитал многих палеонтологов, работая в Калифорнийском университете Лос-Анджелеса). Современные палеонтологи, интеллектуальные «внуки» Ромера и Ольсона, продолжают делать важные открытия.
В 1994 году Смитсоновский институт организовал экспедицию в район Сеймур; её возглавил Хоттон. Группа работала в округе Бейлор. Там обнаружили много ископаемых рыб и амфибий, но в полевых условиях не было возможности очистить все окаменелости и детально их изучить. Рассказывают, что Хоттон распознал важность одной находки (обнаруженной Питером Кролером, ассистентом куратора коллекции в Смитсоновском музее) и носил её в кармане вместе с клочком бумаги, на котором было написано «лягушечка». Но Хоттон умер в 1999 году, не успев изучить находку и опубликовать результаты.
Спустя пять лет несколько молодых учёных извлекли этот образец из запасников и потратили немало часов на тщательное препарирование и полную очистку ископаемого (в образце Хоттона она просматривалась частично). Наконец, в 2008 году окаменелость Хоттона была полностью описана и освещена в печати, через 14 лет после того, как её нашли. Соавторами статьи были Джейсон С. Андерсон из Университета Калгари (учившийся у Кэрролла и Рейша), а также Роберт Рейш, Стюарт Сумида из Калифорнийского университета в Бейкерсфилде (ученик Вона) и Надя Фрёбиш из Берлинского музея естествознания (ученица Кэрролла). Они назвали животное
Gerobatrachus hottoni (древняя лягушка Хоттона), хотя СМИ, растиражировавшие новость об этом открытии, окрестили существо лягомандрой.
Образец представлял собой практически полный скелет длиной всего 11 см. Животное лежало на спине; его бёдра, хвост и плечевые кости частично не сохранились (рис. 11.5). Когда видишь эту окаменелость, в первую очередь бросается в глаза сочетание «саламандрового» тела и широкой плоской лягушачьей морды (отсюда и лягомандра). Существо обладает и многими другими чертами лягушек — это и скелет, и череп, и особенно большая барабанная перепонка. Важнее всего, что зубы животного сидят на маленьких ножках с чётко выраженным основанием (педицеллярные зубы). Такая черта уникальна для всех ныне живущих и некоторых вымерших амфибий.

 Рис. 11.5. Gerobatrachus hottoni
Рис. 11.5. Gerobatrachus hottoni (
А) окаменелость; (
Б) реконструкция живой лягушки (иллюстрация (
А) предоставлена Diane Scott и Jason Anderson; иллюстрация (
Б) предоставлена Nobumichi Tamura)
Окаменелости, не попадающие в современные группы и расположенные прямо между ними, — настоящие переходные формы, иногда (ошибочно) именуемые недостающими звеньями. Геробатрахус — самая настоящая переходная форма, связывающая лягушек и саламандр. Древнейшая известная саламандра — это
Karaurus sharovi, жившая в конце юры (около 150 млн лет назад) и найденная в Казахстане. Древнейшая известная лягушка —
Triadobatrachus massinoti, живший
в начале триаса (240 млн лет назад) на Мадагаскаре (рис. 11.6; см. рис. 11.3). Триадобатрахус похож на современных лягушек: у него широкая морда и длинные перепончатые лапки, но, в отличие от лягушек, в позвоночнике имелся длинный туловищный отдел с 14 позвонками. У всех современных лягушек этот отдел гораздо короче и содержит четыре–девять позвонков. У триадобатрахуса (даже у взрослых особей) сохранялся короткий хвост, чего не наблюдается ни у одной современной лягушки. Его задние лапки были больше, чем у любой саламандры, но далеко не такими крупными, как у современных лягушек. Поэтому триадобатрахус хорошо плавал, но не умел прыгать. Благодаря всем этим чертам, а также многим другим, триадобатрахус — идеальное ископаемое переходное звено между современными лягушками и более примитивными формами, такими как геробатрахус (лягомандра).
 Рис. 11.6.
Рис. 11.6. Реконструкция примитивной триасовой лягушки триадобатрахуса (иллюстрация предоставлена Nobumichi Tamura)
Геробатрахус, живший 290 млн лет назад, намного старше любых представителей эволюционной линии лягушек и саламандр. Он обладает настолько примитивными чертами, что его нельзя отнести ни к лягушкам, ни к саламандрам. Это доказывает, что лягушки и саламандры не были созданы как отдельные «роды», а развивались от общих предков, одним из которых является геробатрахус.
Лучше один раз увидеть!
Насколько мне известно, лягомандра не экспонируется ни в одном музее. Но большие окаменелости пермских амфибий Техаса, в том числе эриопс и диплокаулус, есть в Американском музее естественной истории (Нью-Йорк), Денверском музее природы и науки, Музее естественной истории под открытым небом (Чикаго), Музее сравнительной зоологии Гарвардского университета (Кембридж, Массачусетс), Национальном музее естественной истории Смитсоновского института (Вашингтон, округ Колумбия), а также в Музее естественной истории им. Сэма Ноубла в Оклахомском университете (Норман, штат Оклахома).
Дополнительная литература
Anderson, Jason S., Robert R. Reisz, Diane Scott, Nadia B. Fröbisch, and Stuart S. Sumida. A Stem Batrachian from the Early Permian of Texas and the Origin of Frogs and Salamanders //
Nature. — 2008. — May 22. — P. 515–518.
Bolt, John R. Dissorophid Relationships and Ontogeny, and the Origin of the Lissamphibia //
Journal of Paleontology. — 1977. — Vol. 51. — P. 235–249.
Carroll, Robert. The Rise of Amphibians: 365 Million Years of Evolution. — Baltimore: Johns Hopkins University Press, 2009.
Clack, Jennifer A. Gaining Ground: The Origin and Early Evolution of Tetrapods. — Bloomington: Indiana University Press, 2002.
12. Черепаха с половиной панциря
Происхождение черепах
Одонтохелис
Понаблюдайте за черепахой. Она двигается вперёд, лишь когда высовывается из панциря.
Джеймс Брайан Конант
Черепахи до самого низа
Когда Уильям Джеймс прочитал лекцию о космологии и строении Солнечной системы, к нему обратилась щупленькая старушка. «Мистер Джеймс, ваша теория о том, что Солнце — центр нашей системы, а Земля — шар, который вокруг него вращается, звучит очень убедительно, но она ошибочна. У меня есть теория получше», — сказала старушка. «Какая же, мадам?» — вежливо осведомился Джеймс. «Такая: мы живём на земной коре, а эта кора — не что иное, как спина гигантской черепахи». Не желая развенчивать бредовенькую теорию с привлечением всей массы фактов, которой он располагал, Джеймс решил деликатно переубедить оппонентку, указав ей на определённые нестыковки в её взглядах. «Если ваша теория верна, мадам, — спросил он, — на чём стоит эта черепаха?» «Вы умны, мистер Джеймс, это хороший вопрос, — парировала старушка. — Но у меня есть ответ. Извольте: первая черепаха стоит на второй, гораздо более крупной, расположенной прямо под ней». — «Но на чём же покоится эта вторая черепаха?» — не отступал Джеймс. На что собеседница ликующе ответила: «Полно, мистер Джеймс, там же черепахи до самого низа!».
Существует много вариантов этой истории: одни приписывают философу Бертрану Расселу, другие — философу и психологу Уильяму Джеймсу, писателю Генри Дэвиду Торо, знаменитому скептику Джозефу Баркеру, философу Дэвиду Юму, а также таким учёным, как Артур Эддингтон, Лайнус Полинг и Карл Саган. Они восходят к древнему индийскому мифу о том, что мир зиждется на спине колоссальной черепахи. Например, Бертран Рассел говорил на лекции в 1927 году:
Если у всего должна быть причина, то и у Бога она должна быть. Если может существовать что-либо, не имеющее причины, будь то мир или Бог, то такой взгляд не выдерживает критики. Он будет равноценен индуистским верованиям, что мир покоится на спине у слона, а слон стоит на черепахе. Когда же индусов спрашиваешь, на чём стоит черепаха, они предлагают сменить тему.
Все эти трактовки связаны с проблемой бесконечной регрессии («черепахи до самого низа»), умалчивающей, на чём покоится последняя черепаха. Споры об основополагающих причинах продолжаются веками.
Метафорически эта история ставит перед нами и другой вопрос: если проследить палеонтологическую летопись черепах до самого основания, какое существо мы там найдём? Какое животное — не черепаха как таковая, а переходная форма, чьими ближайшими родичами являются именно черепахи? Как можно было бы создать такую «получерепаху»? Это обычный приём креационистов, пытающихся опровергнуть палеонтологию. Они указывают на большинство ископаемых черепах, подчёркивая, что всё это «просто черепахи» или «представители черепашьего рода», а не переходные формы, связывающие черепах с другими рептилиями. Даже когда предъявляешь им анатомические факты, свидетельствующие, что у древних черепах были те или иные очень примитивные черты, отсутствующие у черепах современных, они настаивают, что это просто черепаха. Не могут себе представить получерепаху. Как у ископаемого животного может быть половина черепашьего панциря, если большинство черепах нуждается для защиты как в его верхней половинке (карапаксе), так и в нижней (пластроне)?
К счастью, благодаря многолетним исследованиям, кульминация которых наступила в 2008 году, палеонтологи обнаружили экземпляры, демонстрирующие основные переходные этапы от обычной рептилии к настоящей черепахе.
Промежуточные черепахи
Прежде чем мы доберёмся до черепахи, которая в самом низу, обратимся к эволюционной истории черепах. Большинство людей полагают, что все черепахи похожи, однако начитывается 455 родов и более 1200 видов этих животных. Многие находятся под угрозой исчезновения из-за того, что люди на них охотятся, разрушают среду обитания и торгуют ими в зоомагазинах. С учётом ограничений, которые на черепах накладывают «доспехи», они могут вести разный образ жизни. Существуют морские черепахи, проводящие всю жизнь и океане, а также пресноводные и сухопутные.
Современные черепахи относятся к двум разным группам. Знакомые всем пресноводные, морские и сухопутные — это скрытошейные. Их легко распознать: когда они прячут голову под козырёк панциря, шея S-образно изгибается, причём шейные позвонки складываются в вертикальной плоскости прямо под карапаксом (рис. 12.1). Снаружи выглядит так, словно черепаха втянула голову прямо в панцирь. Кроме характерных поворотов головы, скрытошейным присущи и иные специализированные черты шейных и челюстных мускулов, не так давно открытые Юджином Гаффни из Американского музея естественной истории.
Вторая группа — бокошейные черепахи. Когда они втягивают голову под панцирь, их шея изгибается в горизонтальной плоскости, причём голова и шея втискиваются прямо под козырёк карапакса (см. рис. 12.1). Бокошейные черепахи — очень специализированная и своеобразная группа, включающая всего около 17 родов и 80 видов. Большинство из них встречаются на материках, когда-то входивших в состав древнего континента Гондваны (это Африка, Австралия и Южная Америка). Палеонтологическая летопись бокошейных не такая полная, как у скрытошейных, но в конце мела и начале кайнозоя разнообразные виды этих черепах встречались и в Гондване, и на древнем северном континенте — в Лавразии. Их нынешнее распространение в пределах Южного полушария связано с более поздним обеднением такого разнообразия.
 Рис. 12.1.
Рис. 12.1. Скрытошейные черепахи (вверху) изгибают шею S-образно в вертикальной плоскости и целиком втягивают голову под панцирь; бокошейные черепахи (внизу) изгибают шею вбок, затягивая голову и шею под козырёк панциря (рисунок Mary P. Williams)
Некоторые бокошейные черепахи в самом деле очень странные. Например, матамата (рис. 12.2), которая живёт на дне рек и озёр в бассейнах Амазонки и Ориноко в Южной Америке. Её панцирь покрыт маскировочными бугорками и бороздами. Ноздри матаматы вытянуты так, что напоминают дыхательную трубку. Поэтому черепаха может затаиться под водой и дышать, высунув лишь кончик носа. Когда рыбки или другая мелкая живность проплывают слишком близко от матаматы, она неожиданно разевает рот, выпрастывает огромную глотку и мгновенно засасывает добычу! Челюсти этой черепахи сильно редуцированы, не дают ей возможности кусать или жевать — только глотать добычу целиком, после того как откачает изо рта и глотки лишнюю воду с помощью сильных шейных мышц.
 Рис. 12.2.
Рис. 12.2. Матамата — современная бокошейная черепаха с редуцированной беззубой челюстью, широким ртом и плоской головой. Она не кусает добычу, а всасывает, широко раскрывая рот и захватывая её объёмистой глоткой (из фонда Wikimedia Commons)
Большинство ископаемых черепах относительно мелкие, примерно таких же размеров, как современные. Хотя на изолированных островах существуют гигантские экземпляры, например на архипелаге Галапагосы к западу от Эквадора и на островах Альдабра в Индийском океане. Крупнейшие из ныне живущих — морские черепахи, которые вырастают до гигантских размеров благодаря подъёмной силе, которую испытывают в своей водной стихии. Самая большая из них — кожистая черепаха (четвёртая по величине среди рептилий), её представители могут достигать более 2,2 м в длину и весить до 700 кг. Черепаха получила такое название, поскольку её костный панцирь почти полностью редуцировался и спинной скелет покрыт лишь тонкой крепкой шкурой. Избавившись от панциря, кожистая черепаха стала сравнительно лёгкой и не очень быстро тонет. При этом кожа достаточно толстая, чтобы отвадить большинство хищников (впрочем, немногие хищники решатся охотиться на взрослую кожистую черепаху).
В геологической истории встречаются по-настоящему исполинские черепахи. Самой крупной морской черепахой был архелон (в переводе с греческого «царь черепах»), плававший в мелких внутренних морях на территории нынешнего западного Канзаса вместе с другими водяными рептилиями — плезиозаврами, ихтиозаврами и мозазаврами (рис. 12.3). Крупнейшие экземпляры архелона достигали более 4 м в длину и около 5 м в размахе плавников. Весила такая черепаха более 2200 кг. Как и у многих морских черепах, у архелона был почти открытый спинной скелет и четыре зазубренные пластины на брюхе. Подобно современной кожистой черепахе, архелон, вероятно, имел вместо панциря толстый слой кожи.
 Рис. 12.3.
Рис. 12.3. Наиболее полный скелет архелона — гигантской морской черепахи, плававшей в меловом периоде в морях Канзаса (фотография предоставлена Естественнонаучным музеем Пибоди Йельского университета (Нью-Хейвен, штат Коннектикут))
Вымершие гигантские сухопутные черепахи не вырастали до таких размеров, тем не менее по сравнению с ними все современные наземные черепахи кажутся лилипутами. Одним из крупнейших был колоссохелис, достигавший более 2,7 м в длину, столько же в ширину, весивший тонну и более. Ископаемые останки колоссохелиса открыли в Пакистане в 1840-е годы, а затем обнаружили в разных регионах, от Европы до Индии и Индонезии. Возраст этих окаменелостей — от нескольких миллионов до 10 000 лет, то есть последние из них относятся к концу последнего ледникового периода. Вероятно, колоссохелис выглядел как увеличенная копия галапагосской черепахи.
Ещё крупнее был карбонемис, найденный в болотных отложениях Колумбии, которым около 60 миллионов лет. По размеру он сравним с автомобилем Smart (более 1,7 м в длину) и мог съесть почти любую тварь, какую повстречает, даже крокодила. Это был один из самых крупных обитателей Земли в палеоценовую эпоху — ещё больше, пожалуй, лишь титанобоа, обнаруженная в тех же отложениях. Это крупнейшая в истории змея, достигавшая 14 м в длину (см. главу 13). Карбонемис, подобно большинству южноамериканских черепах, являлся представителем пеломедузовых бокошейных, обычных для Южной Америки.
Крупнейшей пресноводной черепахой всех времён был ещё один южноамериканский монстр под названием ступендемис, обнаруженный в болотных отложениях формации Урумако в Венесуэле (возраст — 5–6 млн лет), также найденный в Бразилии (рис. 12.4). Подобно карбонемису, он относился к пеломедузовым бокошейным черепахам; похож на современную черепаху аррау (
Podocnemis expansa), но гораздо крупнее. Обладал колоссальным панцирем: 3,3 м в длину и 1,8 м в ширину.
Такие гигантские экземпляры дают представление об исключительном эволюционном разнообразии водных и сухопутных черепах. Возникает вопрос: какие черепахи расположены ниже современных скрытошейных и бокошейных, то есть более примитивны?
 Рис. 12.4.
Рис. 12.4. Панцирь ступендемиса, выставленный в Естественнонаучном музее Химедзи (префектура Хёго, Япония) (фотография из фонда Wikimedia Commons)
Первая пресноводная черепаха
Древнейшая из известных пресноводная черепаха (до 2008 года — самая древняя из известных черепах) называлась проганохелис. Она была невелика (около метра в длину) и крайне примитивна, устроена проще, чем любые современные скрытошейные или бокошейные. У неё имелась длинная шея, покрытая прочными шипами (следовательно, голова не втягивалась под панцирь). Известно несколько полных или почти полных скелетов проганохелиса. Первые нашли в верхнетриасовых отложениях в Германии, они имеют возраст около 210 млн лет. Позднее таких черепах обнаружили также в Гренландии и Таиланде.

 Рис. 12.5.
Рис. 12.5. Проганохелис, древнейшая пресноводная черепаха. (
А) ископаемый экземпляр и панцирь; (
Б) реконструкция внешнего облика ((
А) фотография из фонда Wikimedia Commons; (
Б) предоставлено Nobumichi Tamura)
Для человека, не разбирающегося в анатомии или зоологии, проганохелис кажется самой обычной черепахой. Но при внимательном изучении выясняется, что он сильно отличался от более поздних экземпляров. Хотя у него и был карапакс, в верхней части панциря проганохелис имел гораздо больше пластин, чем любая более поздняя ископаемая или ныне живущая черепаха. Особенно много пластин шло по краю панциря — они защищали лапы. У проганохелиса имелся длинный хвост с жёсткой шипастой оболочкой, который заканчивался «булавой». Эта черепаха жила бок о бок с первыми динозаврами и, таким образом, конкурировала с многочисленными хищниками. Череп проганохелиса (по сравнению с любыми ныне живущими черепахами) сильно напоминал череп более примитивных рептилий. Хотя у него был клюв, как у современных черепах, на нёбе оставались зубы. То есть это последняя в истории зубастая черепаха. Поскольку животное обладало и клювом, и зубами, вероятно, оно было всеядным — питалось как животными, так и растениями. Шея проганохелиса не складывалась, он не мог втянуть под панцирь свою большую «армированную» голову (в отличие от скрытошейных и бокошейных черепах).
Итак, мы добрались до древнейшей из известных пресноводных черепах. Проганохелис определённо был черепахой и имел панцирь, хотя по большинству других признаков напоминал примитивную рептилию и сильно отличался от более поздних черепах. Креационисты долго отказывались видеть его особенности, считая «просто черепахой», и твердили, что представить черепаху без панциря невозможно. Наконец, в 2008 году все вопросы о происхождении черепах удалось разрешить.
Черепаха с половиной панциря
Китайские палеонтологи месяцами разрабатывали важное местонахождение окаменелостей: Гуаньлиньскую биоту близ деревушки Синьпу в провинции Гуйчжоу (регион в восточной части центрального Китая, к западу от Гонконга, отделённый ещё одной провинцией от границы с Вьетнамом). Чёрные сланцы пачки пород Ваяо в формации Фалань откладывались в бассейне реки Наньпаньцзян на протяжении позднего триаса (около 220 млн лет назад). Этот бассейн был с трёх сторон окружён возвышенностями, но в его юго-западной части располагался лиман, относившийся к древнему океану Тетис, некогда простиравшемуся от Средиземноморья до Индонезии. Чёрные сланцы — типичные залежи, формирующиеся в глубоких стоячих водах. В них очень медленно протекают процессы гниения и разложения, поэтому любые окаменелости изумительно сохраняются. Хотя вода здесь была глубока и бедна кислородом, до земли тоже было недалеко, о чём свидетельствует окаменевший плавник и останки сухопутных животных. Вероятно, некоторые из этих существ плавали на морском мелководье или в заливах, вдававшихся в бассейн Наньпаньцзян.
За много лет Гуаньлиньская биота преподнесла массу удивительных окаменелостей морских рептилий, а также морских беспозвоночных (особенно аммонитов и гигантских «морских лилий», также называемых криноидеями). По ним можно проследить изменения, происходившие в мировом океане в конце триаса. Что касается рептилий, здесь попадаются почти полные скелеты «рыбоящеров» (ихтиозавров) до 10 м в длину (см. главу 15) и так называемых плакодонтов — пресмыкающихся, питавшихся моллюсками. Здесь есть представители и другой группы морских рептилий — талаттозавров. Ранее считалось, что Гуаньлиньская биота включает 17 специфических родов, но последние исследования позволили сократить этот список до восьми родов и видов.
Открывая новые виды рептилий, китайские учёные собрали очень интересную коллекцию окаменелостей, информацию о которой опубликовали в 2008 году. Взяв за основу один полный и несколько частично сохранившихся скелетов, они назвали странное животное
Odontochelys semitestacea (переводится как «зубастая черепаха с половиной панциря»). Сложно представить себе более выразительное переходное звено между черепахами и другими рептилиями (рис. 12.6). Одонтохелис позволяет ответить на вопрос, как из беспанцирной рептилии могла развиться черепаха в панцире. У него на брюхе имелся полноценный костный щит (пластрон), на спине — лишь крепкие рёбра, и никакого панциря! Другими словами, сначала у черепахи образовалась нижняя половина панциря, потом карапакс. Одонтохелис — настоящая «черепаха с половиной панциря».

 Рис. 12.6.
Рис. 12.6. Одонтохелис (
А) наиболее полная из известных окаменелостей; на спине животного — зачаточный карапакс (слева), на брюхе — полноценный пластрон (справа); (
Б) реконструкция внешнего облика (иллюстрация (
А) предоставлена Li Chun; иллюстрация (
Б) предоставлена Nobumichi Tamura)
Кроме этой замечательной черты, у одонтохелиса была и другая, не менее удивительная — полный комплект зубов на челюстях. Это последняя черепаха, обладавшая зубами, а не беззубым клювом. Опять же, можно проследить эволюционный переход от обычных рептилий к черепахам по зубам: у получерепахи одонтохелиса имелись обычные зубы, как у всех пресмыкающихся; у проганохелиса уже не было челюстных зубов, но оставались зубы на нёбе; более поздние черепахи лишены зубов.
Одонтохелис ставит точку и в другом давнем споре. Десятилетиями одни учёные полагали, что черепаший карапакс развился из сросшихся крошечных пластин ороговевшей кожи (остеодерм), а другие считали, что панцирь образовался прежде всего благодаря разрастанию спинных рёбер. Одонтохелис подтверждает правоту второй группы: его широкие спинные рёбра начинают соединяться и срастаться в панцирь, а поверх рёбер или между ними нет остеодерм. Эти данные подтверждают эмбриологические исследования черепах, в ходе которых прослеживается развитие карапакса по онтогенетическим изменениям спинных рёбер; остеодермы в данном процессе не участвуют.
Наконец, одонтохелис отвечает ещё на один вопрос: в какой среде первоначально развивались черепахи. Большинство более поздних ископаемых черепах, тот же проганохелис, найдены в отложениях, сформировавшихся на суше. Поэтому палеонтологи думали, что черепахи изначально были сухопутными животными. Но древнейшая известная черепаха одонтохелис — однозначно водный организм; она жила в открытом океане и, возможно, заплывала в устья и дельты рек по всему миру. По пропорциям своих передних конечностей одонтохелис напоминал многих пресноводных черепах, населявших небольшие (в том числе проточные) водоёмы.
Под пирамидой черепах
Одонтохелис — настоящее переходное ископаемое звено, соединяющее обычных рептилий и истинных черепах. Но от каких эволюционных ветвей пресмыкающихся произошли черепахи? Традиционно считалось, что они — потомки самой примитивной группы рептилий, анапсид, не имевших особых отверстий в черепе, характерных для более высокоразвитых сородичей. Такое мнение существует более ста лет и по-прежнему является самым распространённым.
Однако в последние 20 лет оно оспаривается благодаря новым источникам данных: молекулярному секвенированию ДНК и белков, характерных для всех рептилий. Ряд таких исследований позволяет отнести черепах к диапсидам — группе, включающей ящериц, змей, а также крокодилов и птиц. Одни исследования сближают черепах с ящерицами, другие — с крокодилами и птицами.
Правда, новейший анализ, проведённый группой учёных из Йельского университета и Тюбингенского университета в Германии даёт убедительные доказательства, что черепахи — самые примитивные из ныне живущих рептилий. Они ведут к эунотозавру, ископаемому животному из Южной Африки, впервые описанному в 1892 году Гарри Говиром Сили (рис. 12.7). Это существо довольно часто встречается в среднепермских отложениях, сформировавшихся около 270 млн лет назад, но полные скелеты с хорошо сохранившимися черепами попадаются редко. Внешне эунотозавр напоминал крупную жирную ящерицу, но имел важные особенности скелета. Самая замечательная из них — сильно расставленные широкие и плоские спинные рёбра, почти соединённые друг с другом и образовавшие цельный панцирь. Эта особенность и другие анатомические черты убедили многих учёных, что черепахи происходят от эволюционной ветви примитивных рептилий. Палеонтологи утверждали, что молекулярный анализ ошибочен, и виной тому проблема, называемая «эффект притяжения длинных ветвей»: изолированная группа, на раннем этапе отходящая от эволюционного древа, приобретает генетические паттерны, осложняющие её классификацию.

 Рис. 12.7.
Рис. 12.7. Эунотозавр — примитивная пермская рептилия с широко расставленными рёбрами, древнейшее звено эволюции черепах; (
А) частично сохранившийся экземпляр с хорошо заметными изогнутыми рёбрами, образующими зачаточный панцирь; (
Б) родословное древо эунотозавра и других примитивных черепах, демонстрирующее переход от обычных рептилий к черепахам (фотография (
А) предоставлена B. Rubidge, Институт эволюционных исследований Университета Витсватерсранд (Йоханнесбург, ЮАР); (
Б) перерисовано Э. Протеро; оригинал взят из
Tyler R. Lyson et al. Transitional Fossils and the Origin of Turtles //
Biology Letters. — 2010. — Vol. 6)
Поиск продолжается, и вопрос, какая рептилия породила черепах, по-прежнему открыт. Именно так обычно идёт научный процесс, пока не появляются чёткие и неопровержимые факты (одним из них стала первая публикация об одонтохелисе). Будьте начеку — возможно, к моменту выхода этой книги будет найден другой ответ!
Лучше один раз увидеть!
Насколько мне известно, одонтохелис нигде не экспонируется, но в Американском музее естественной истории (Нью-Йорк) можно посмотреть на многих других ископаемых черепах: колоссохелиса, ступендемиса, проганохелиса. В Естественнонаучном музее Пибоди Йельского университета (Нью-Хейвен, штат Коннектикут) есть крупнейший и самый знаменитый экземпляр архелона; другие экземпляры этой черепахи выставляются в Американском музее естественной истории и Музее естествознания (Вена). Отпечатки ступендэмиса также имеются в Естественнонаучном музее Химедзи (префектура Хёго, Япония) и в Естественнонаучном музее города Осака. В Музее естествознания Штутгарта экспонируются некоторые окаменелости проганохелиса, найденные в Германии.
Дополнительная литература
Bonin, Franck, Bernard Devaux, and Alain Dupré. Turtles of the World. / Translated by Peter C. H. Pritchard. — Baltimore: Johns Hopkins University Press, 2006.
Brinkman, Donald B., Patricia A. Holroyd, and James D. Gardner, eds. Morphology and Evolution of Turtles. — Berlin: Springer, 2012.
Ernst, Carl H., and Roger W. Barbour. Turtles of the World. — Washington, D. C.: Smithsonian Institution Press, 1992.
Franklin, Carl J. Turtles: An Extraordinary Natural History 245 Million Years in the Making. — New York: Voyageur Press, 2007.
Gaffney, Eugene S. A Phylogeny and Classification of the Higher Categories of Turtles //
Bulletin of the American Museum of Natural History. — 1975. — Vol. 155. — P. 387–436.
Laurin, Michel, and Robert R. Reisz. A Reevaluation of Early Amniote Phylogeny //
Zoological Journal of the Linnean Society. — 1995. — Vol. 113. — P. 165–223.
Li, Chun, Xiao-Chun Wu, Olivier Rieppel, Li-Ting Wang, and Li-Jun Zhao. An Ancestral Turtle from the Late Triassic of Southwestern China //
Nature. — 2008. — November 27. — P. 497–501.
Orenstein, Ronald. Turtles, Tortoises, and Terrapins: A Natural History. — New York: Firefly Books, 2012.
Wyneken, Jeanette, Matthew H. Godfrey, and Vincent Bels. Biology of Turtles: From Structures to Strategies of Life. — Boca Raton, Fla.: CRC Press, 2007.
13. Ходячие змеи
Происхождение змей
Хаасиофис
И сказал Господь Бог жене: что ты это сделала? Жена сказала: змей обольстил меня, и я ела.
И сказал Господь Бог змею: за то, что ты сделал это, проклят ты пред всеми скотами и пред всеми зверями полевыми; ты будешь ходить на чреве твоём, и будешь есть прах во все дни жизни твоей;
и вражду положу между тобою и между женою, и между семенем твоим, и между семенем её; оно будет поражать тебя в голову, а ты будешь жалить его в пяту.
Бытие, 3:13–15
Батюшки, живые змеи!
Если и есть среди животных те, на кого человек всегда остро реагирует, — это, безусловно, змеи. Они — одни из самых ненавистных и устрашающих животных, хотя в действительности большинство змей полезны человеку, так как истребляют грызунов и других вредителей. И всё же люди испытывают перед змеями сильный, зачастую иррациональный страх, который может перерасти в парализующую фобию (офидиофобию). Холодные немигающие глаза змей, трепещущие языки и ползание на брюхе многих нервирует.
Главный фактор этого практически всеобщего страха перед змеями заключается в том, что некоторые из них ядовиты. Десять наиболее распространённых змей в Австралии крайне опасны, поэтому страх оправдан. В тропической Азии и Африке ядовитые змеи тоже составляют значительный процент. Правда, в США распространены лишь три ядовитых вида: гремучая змея, медноголовый щитомордник и водяной щитомордник. Они соседствуют с более многочисленными безвредными змеями, которых мы часто убиваем. Большинство людей отказывают змее в праве на жизнь, тем более не стремятся изучить и понять её. Исключением являются те из нас, кто интересуется естествознанием; есть даже такие, чьё увлечение рептилиями превращается в серьёзный интерес, а иногда и в научную карьеру в области герпетологии.
Вероятно, поскольку мы сосуществовали со змеями на протяжении долгой эволюционной истории, змеи оставили глубокий след в человеческой культуре, часто фигурируют в мифах и легендах. В Древнем Египте кобра украшала корону фараона. У Медузы, одной из горгон в древнегреческой мифологии, змеи росли на голове вместо волос. Гераклу довелось убить Лернейскую гидру, отрубив ей девять змеиных голов, причём на месте каждой отсечённой головы вырастала новая. Греки также высоко ценили медицинские свойства змей, поэтому жезл кадуцей — символ исцеления, обвит двумя змеями. Змеи почитаются в индуизме и буддизме. Например, шея индуистского бога Шивы увита змеями, бог Вишну изображается спящим на семиглавой змее или среди её колец. Кроме того, змеи играли важную роль в центральноамериканской мифологии и религии. Китайцы издавна их почитали, а также считали деликатесом в традиционной кухне. Змея — один из 12 знаков китайского зодиака. Наконец, в третьей главе Книги Бытия змей в Эдемском саду склоняет Еву вкусить плод с Древа познания добра и зла. Даже в наше время есть христианские секты, практикующие ритуал змееношения (большинство его участников умирают от ядовитых укусов).
Независимо от вашего субъективного отношения к змеям, это, бесспорно, одна из самых успешных и разнообразных групп животных на Земле. Несмотря на специализированный хищнический образ жизни (все змеи питаются исключительно живой добычей), насчитывается 2900 видов этих животных, объединённых в 29 семейств и множество родов. Они распространены от Северного полярного круга в Скандинавии и далее на юг. Встречаются на всех континентах, кроме Антарктики, и даже на высоте 4900 м в Гималаях. Живут и ниже уровня моря, в тёплых прибрежных водах от Индийского океана до Западной Пацифики. На многих островах змеи не водятся: Гавайи, Исландия, Ирландия, Новая Зеландия, большинство островов в южной части Тихого океана. Но не потому, что их оттуда изгнал святой Патрик или кто-то другой. Скорее всего, змеям просто не удалось попасть на эти острова с ближайших материков, даже когда уровень моря снизился на пике последнего Великого оледенения и большинство сухопутных животных перебрались на отдалённые острова. Некоторые из этих островов (например, Ирландия) были практически полностью покрыты ледяными щитами, тогда как другие (Гавайи) просто находились слишком далеко.
Змеи обладают многими примечательными чертами, некоторые из них уникальны. Так, их череп состоит из ряда мелких костных распорок, сочленённых эластичными связками и сухожилиями (рис. 13.1). Поэтому змеи могут широко разевать пасть и целиком захватывать живую добычу, а потом медленно перемалывать челюстями тело, пока жертва не будет полностью проглочена. Ещё змеи умеют задерживать дыхание, пока добыча не пройдёт через глотку, и неделями медленно переваривают такую пищу целиком. Всё это время они часто находятся в состоянии оцепенения и прячутся, пока не закончится сложный процесс переваривания твёрдого непережёванного скелета. Можно наблюдать, как ком плоти продвигается по змеиному телу по ходу пищеварения.
 Рис. 13.1.
Рис. 13.1. Змеиный череп с изящными костями (из фонда Wikimedia Commons)
Хотя некоторые змеи видят хорошо, большинство из них различают лишь размытое изображение вокруг себя и обычно реагируют в первую очередь на движение; есть и слепые змеи. Не полагаясь на зрение, большинство из них пробуют воздух трепещущим языком, пользуясь вомероназальным органом, расположенным на верхнем нёбе и интерпретирующим улавливаемые запахи. Кроме того, на морде у многих змей есть специальные теплочувствительные ямки, помогающие им обнаруживать теплокровных животных (как хищников, так и добычу). Змеи утратили наружные уши, поэтому большинство из них «слушают» вибрацию, отдающуюся в нижней челюсти. Хотя бы поэтому «заклинание змей» — вымысел: поскольку выпрямляющаяся змея должна припадать нижней челюстью к земле, чтобы что-нибудь услышать, она реагирует на движения заклинателя, а не на звуки флейты.
Вслед за черепом у змеи идут от 200 до почти 400 позвонков. Для сравнения: у человека всего 33 позвонка, а у большинства хвостатых животных — до 50. Практически всё тело змеи состоит из многочисленных рёбер, покрытых перекрещивающимися мышечными связками, с помощью которых змея двигается и извивается при ползании. Большая часть змеиного тела — очень длинный туловищный отдел («грудная клетка») и короткий хвост. Обычно у змеи два лёгких, но левое может быть сильно редуцировано или отсутствовать, так как объём тела ограничен. Самые примитивные змеи (особенно удавы и близкие им виды) имеют рудименты бедренных и тазовых костей. Они утратили функцию конечностей, но используются во время брачных игр и поединков. Эти рудименты свидетельствуют о том, что предки змей были четвероногими животными.
При столь специфическом строении тела змеи могут иметь очень разные размеры. Самая маленькая — барбадосская узкоротая змея, всего 10 см в длину, вполне может обвиться вокруг небольшой монетки. Большинство змей вырастают до метра и более — они достаточно велики, чтобы одолеть свою типичную добычу: грызунов, других мелких млекопитающих и птиц (иногда и других змей). Настоящие гиганты — сетчатый питон и анаконда, два огромных удава. Анаконда — змея-пловец, уволакивающая добычу под воду и выдавливающая из неё воздух. Она может достигать 6,6 м в длину и весить до 70 кг. Сетчатый питон не такой массивный, но бывает ещё длиннее — до 7,4 м. Обе змеи так велики, что могут заглатывать и крупную добычу: коз, овец, телят, капибар. Однако они несравнимы с гигантами прошлого.
Титанобоа, недавно открытая в палеоценовых отложениях на территории Колумбии (возраст — 60–58 млн лет) обнуляет рекорды современных змей, той же анаконды (рис. 13.2). Хотя она известна по сотням позвонков и фрагментам черепа, размеры костей настолько велики, что, по оценкам учёных, змея могла достигать длины в 15 м (примерно со школьный автобус) и весить около 1135 кг. Титанобоа жила примерно через 5 млн лет после вымирания динозавров. В тропических болотах Колумбии рядом с ней обитали огромные крокодилы, черепахи и иные колоссальные рептилии. Вероятно, она вырастала до таких размеров благодаря отсутствию крупных хищных млекопитающих, которые ещё не развились, а также гигантских динозавров. Соответственно, ниша суперхищников была занята огромными рептилиями: змеями, крокодилами, черепахами.

 Рис. 13.2.
Рис. 13.2. (
А) Джонатан Блоч сравнивает большой позвонок титанобоа с более мелким позвонком анаконды; (
Б) реконструкция титанобоа в натуральную величину ((
А) фотография Jeff Gage, Флоридский музей естественной истории; (
Б) фотография предоставлена Смитсоновским институтом)
Титанобоа побила прежний рекорд, принадлежавший гигантофису — огромной змее из вымершего гондванского семейства матсоид, известной по эоценовым отложениям в Египте и Марокко (жила 40 млн лет назад). Гигантофис достигал 10,7 м в длину, то есть был намного больше крупнейшей анаконды или сетчатого питона. Ещё одна змея из семейства матсоид называется вонамби и жила в Австралии в эпоху последнего ледникового периода. Она имела 6 м в длину и была одной из огромнейших рептилий и крупнейших хищников материка. Однако голова у неё была маленькая, и она не могла есть дипродонтов — родственников вомбата, сравнимых по размеру с носорогом, либо гигантских кенгуру, населявших Австралию во времена ледникового периода. Хотя с почти любой другой дичью гигантофис справлялся. Эта змея вымерла около 50 тыс. лет назад, вместе с многими представителями сумчатой австралийской мегафауны.
Откуда взялись змеи
Змеи демонстрируют чудеса адаптации и эволюционного успеха, причём с тех времён, когда с лица Земли исчезли динозавры. Но откуда они взялись? Как четвероногая рептилия превратилась в змею? Где переходные формы, подтверждающие такую эволюцию?
На самом деле потеря ног — простейшая часть трансформации. Это произошло со многими группами четвероногих животных, и все они развивались независимо. К безногим рептилиям относятся не только змеи, но и группа существующих ныне амфисбен, а также некоторые ящерицы, в том числе отдельные сцинки, австралийские чешуеноги, веретеницы и желтопузики. Среди амфибий выделяются безногие (
Apoda), у которых развились червеобразные тела, а также сирены, сохранившие укороченные передние конечности и утратившие задние. Кроме того, минимум две вымершие группы амфибий, аистоподы и лисорофиды, тоже стали безногими. Практически все эти животные зарывались в землю, поэтому без конечностей им было, очевидно, проще пробираться через грунт или мягкий ил. Есть простая причина, по которой легко утратить конечности. Развитием зачатков конечностей и самих лапок управляет определённый набор Hox-генов и Tbx-генов: стоит им перестать отдавать команду отращивать конечности — и те исчезают.
Тем не менее представляется маловероятным обнаружить ископаемую змею, которая находилась бы на этапе потери конечностей. Как правило, змеи плохо фоссилизируются, поскольку их тело состоит из сотен тонких позвонков и рёбер, а они ломаются и расчленяются. Известны единичные экземпляры по частично сохранившимся или полным скелетам. Большинство ископаемых змей обнаружены по нескольким позвонкам, и внешний облик животного выстраивается по скудным данным о форме позвоночника.
Несмотря на эти препятствия, палеонтологическая летопись содержит много замечательных окаменелостей, позволяющих отследить переход от четвероногих ящериц к безногим змеям. На первом этапе находим множество фрагментарных окаменелостей из юрского периода. Далее следует
Adriosaurus microbrachis — ископаемое животное, найденное в 2007 году в среднемеловых породах (возраст — около 95 млн лет) на территории Словении (рис. 13.3,
А). Это название переводится как «адриатическая ящерица с маленькими передними лапками». Адриозавр был очень тонкотелой длинной морской ящерицей с полностью функциональными передними лапками, но неработающими рудиментарными задними конечностями.

 Рис. 13.3.
Рис. 13.3. Промежуточное ископаемое звено адриозавр: у этого животного были крошечные передние лапки, но полноценные задние конечности: (
А) скелет; (
Б) реконструкция внешнего облика (иллюстрации предоставлены M. W. Caldwell)
Далее встречаем разнообразных змей, у которых уже нет передних конечностей, но по-прежнему сохраняются крошечные нефункциональные задние. Например,
Najash rionegrina была земляной сухопутной змеёй; она описана в 2006 году по окаменелостям в возрасте около 90 млн лет, найденным в формации Канделерос на территории Аргентины (Нахаш — древнее библейское наименование змея, жившего в Эдемском саду). У
Najash сохранялся тазовый пояс — позвонки, сочленяющиеся с крестцом, а также рудиментарные задние конечности, в которых имелись бедренные и берцовые кости.
Ещё более специализированными и змееподобными являются экстраординарные ископаемые пресмыкающиеся, найденные в позднемеловых морских породах на территории Израиля и Ливана. Из этих ископаемых лучше других сохранился
Haasiophis terrasanctus (рис. 13.4). Имя этому животному — Змея Хааса из Святой Земли — дали в честь австрийского палеонтолога Георга Хааса, обнаружившего его местонахождение и успевшего описать окаменелость до своей смерти в 1981 году. Хаасиофиса нашли в известняках близ местечка Эйн-Ябруд на территории Иудейских гор, что в районе города Рамалла на Западном берегу реки Иордан. Возраст окаменелости — 94 млн лет. Она представляет собой почти полный скелет (88 см в длину), в котором не хватает лишь кончика хвоста. Череп и большинство позвонков выглядят примерно так, как у большинства других примитивных змей. Однако здесь присутствовали и задние конечности — очень маленькие, но с бедренной костью, обеими берцовыми и частью ступни. В отличие от задних конечностей
Najash, тазовые кости
Haasophis очень маленькие и уже не связаны с позвоночником; они полностью рудиментарны и бесполезны.
Haasophis и многие другие змеи мелового периода, видимо, имели вертикальный плавник и хвост, напоминавший весло, как современные морские змеи.

 Рис. 13.4.
Рис. 13.4. Двуногая змея хаасиофис: (
А) полностью сочленённый скелет с сохранившимися рудиментарными задними конечностями (крупные тёмные блоки — пробковые вставки, благодаря которым на образец ничего нельзя положить); (
Б) фрагмент рудиментарных задних конечностей (иллюстрации предоставлены M. Polcyn, Южный методистский университет)
Чуть более крупная змея из Эйн-Ябруда называется пахирахис; её Хаас описал в 1979 году. Хотя данные окаменелости более фрагментарны, чем у хаасиофиса, на метровом теле также имеются крошечные рудиментарные задние конечности. Рёбра и позвонки пахирахиса очень толстые и плотные, что, вероятно, помогало ему плавать на большой глубине в меловых морях.
Третья змея из морских ближневосточных известняков —
Eupodophis descouensi, найденная на территории Ливана (недалеко от Эйн-Ябруда) в породах возрастом около 92 млн лет (рис. 13.5). Родовое название этого животного означает «настоящая змея с ногами», а видовое дано в честь французского палеонтолога Дидье Декуэна. Это существо имело 85 см в длину, примерно столько же, сколько хаасиофис, но его конечности были редуцированы ещё сильнее, чем у двуногих змей мелового периода: хаасиофиса и пахирахиса.
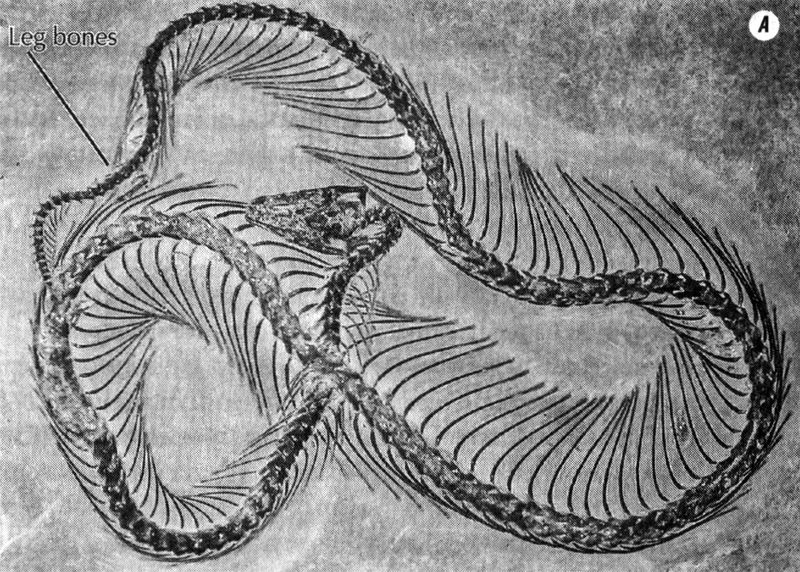
 Рис. 13.5.
Рис. 13.5. Двуногая змея эуподофис: (
А) полный скелет с сохранившимися рудиментарными задними конечностями; (
Б) фрагмент позвоночника, на котором также заметны рудиментарные задние конечности (иллюстрации предоставлены M. W. Caldwell)
Таким образом, не только рудиментарные задние конечности нескольких вымерших позднемеловых морских змей, но и рудиментарные тазовые и бедренные кости (иногда с крошечными отходящими от них «веточками») примитивных ныне живущих змей, в частности удавов и их родичей, — немые, но убедительные свидетельства происхождения змей от четвероногих существ.
Но каков общий предок всех змей? Первые идеи об этом высказал один из пионеров палеонтологии и герпетологии Эдвард Дринкер Коуп, заметивший, что анатомически змеи во многом схожи с варанами, например с австралийскими и комодскими (тем более с мозазаврами — морскими ящерами мелового периода). Анатомические факты тоже, видимо, подкрепляют родство змей с варанами, а последние данные молекулярной биологии — нет; они неоднозначны. Одни молекулярные последовательности позволяют считать змей ближайшими родичами варанов, другие не позволяют считать их родственниками какого-либо из ныне существующих семейств ящериц.
Точка зрения, согласно которой змеи потеряли конечности, переселяясь в море,
вероятно, подкрепляется многими окаменелостями морских змей из меловых пород восточного Средиземноморья (Словения, Израиль, Ливан). Согласно этой версии, утрата ушных раковин и появление цельных прозрачных змеиных век может быть обусловлены адаптацией к плаванию, а не к норному образу жизни.
Другая зоологическая школа развивает идею о том, что змеи могли произойти от земляных, а не от плавающих ящериц — таких, как калимантанский земляной безухий варан. Сторонники этой идеи считают, что прозрачные веки должны были предохранять змеиные глаза от грубого крупного песка при зарывании в землю, а ушная раковина могла исчезнуть, потому что без неё грязь меньше попадает в ушную полость. «Сухопутные» адаптационные черты
Najash согласуются с данной точкой зрения, хотя он жил несколько позднее, чем морские змеи хаасиофис, пахирахис и эуподофис. Самой примитивной из всех известных змей является кониофис, имевший ящеричью голову и змеиное тело. Правда, его окаменелость слишком плохо сохранилась, чтобы по ней можно было судить о конечностях животного. Тем не менее он был сухопутным, а не морским. Водяная ящерица адриозавр — ещё более примитивный родич змей. Она имела четыре лапы и плавала в океане.
Соответственно, тайна ближайших родичей змей ещё не раскрыта. Именно так развивается наука: противоречия наподобие вышеупомянутых очень важны для научного процесса, только так мы можем тщательно изучать все факты и не отбрасывать имеющиеся версии. Независимо от того, как в итоге разрешится спор, есть факт: многие окаменелости обладают чертами, иллюстрирующими переход от четвероногости к двуногости и далее к безногости. Это подтверждает, что змеи произошли от четвероногих предков.
Лучше один раз увидеть!
Реконструкция титанобоа в натуральную величину экспонируется в музее Университета штата Небраска (город Линкольн).
Дополнительная литература
Caldwell, Michael W., and Michael S. Y. Lee. A Snake with Legs from the Marine Cretaceous of the Middle East //
Nature. — 1997. — April 17. — P. 705–709.
Head, Jason J., Jonathan I. Bloch, Alexander K. Hastings, Jason R. Bourque, Edwin A. Cadena, Fabiany A. Herrera, P. David Polly, and Carlos A. Jaramillo. Giant Boid Snake from the Paleocene Neotropics Reveals Hotter Past Equatorial Temperatures //
Nature. — 2009. — February 5. — P. 715–718.
Rieppel, Olivier. A Review of the Origin of Snakes //
Evolutionary Biology. — 1988. — Vol. 25. — P. 37–130.
Rieppel, Olivier, Hussan Zaher, Eitan Tchernove, and Michael J. Polcyn. The Anatomy and Relationships of Haasiophis terrasanctus, a Fossil Snake with Well-Developed Hind Limbs from the Mid-Cretaceous of the Middle East //
Journal of Paleontology. — 2003. — Vol. 77. — P. 536–558.
14. Царь рыбоящеров
Крупнейшая морская рептилия
Шонизавр
She sells sea-shells on the sea-shore.
The shells she sells are sea-shells, I’m sure.
For if she sells sea-shells on the sea-shore
Then I’m sure she sells sea-shore shells.
Английская скороговорка
Она продаёт ракушки на морском берегу
В конце XVIII века приморский городок Лайм-Реджис на Дорсетском побережье в Южной Англии снискал славу популярного летнего курорта для богатых, где леди и джентльмены могли поплескаться в волнах и насладиться прохладным морским бризом. В то время было модно собирать ракушки, окаменелости и иные диковины. Все считали окаменелости оригинальными вещицами, встречающимися в горных породах, которые интересно называть и обозначать. Однако они не открывали ничего такого, что не было бы описано в Книге Бытия. Ещё никто не знал о динозаврах (их начали открывать в 1820–1830-е годы), равно как и о большинстве других вымерших земных организмов. На самом деле, большинство людей (особенно учёные) отрицали возможность вымирания, так как Бог оберегал каждого воробышка и, естественно, не допустил бы исчезновения каких-то своих созданий. По выражению Александра Поупа в его поэме «Опыт о человеке»: «Так видит Бог игру Вселенной всей, // Где обречён герой и воробей». К 1795 году скромный английский инженер и прокладчик каналов по имени Уильям Смит стал замечать, что на территории Британии окаменелости встречаются в строго определённом порядке, но лишь спустя двадцать лет люди поняли значение этого открытия.
В Лайм-Реджисе жил бедный плотник-краснодеревщик Ричард Эннинг с женой Молли, еле-еле сводившие концы с концами. В семье было много детей, но почти все они умерли в юном возрасте — обычное дело в те времена, поскольку медицина оставалась неразвитой, и детские болезни грозили смертью. Старшая дочь погибла в возрасте четырёх лет — на ней случайно загорелась одежда. Спустя пять месяцев после этой трагедии, в 1799 году, родилась Мэри Энн. Когда ей был год и три месяца, в деревушке ударом молнии убило нескольких женщин, в том числе ту, которая держала Мэри на руках. Сама девочка осталась невредимой. Мэри недолго ходила в церковную школу, где научилась читать и писать. Но в те годы женщины-простолюдинки редко получали образование, поэтому, повзрослев, она стала выбираться с отцом и старшим братом Джозефом (из всех братьев и сестёр выжили лишь они двое) на долгие вылазки, чтобы собирать окаменелости в прибрежных скалах. Эти скалы относились к нижнеюрским отложениям (возраст — от 210 до 195 млн лет), принадлежавших к формации голубой лейас. В породах было полно «каменных змей» (аммонитов), «чёртовых пальцев» (белемнитов), «чёртовых ногтей» (устриц
Gryphaea) и позвонков. Многие жители городка собирали окаменелости, чтобы летом продавать их богатым туристам и поправлять своё бедственное положение в суровые годы начала XIX века. Вдобавок семья Эннингов сталкивалась с дискриминацией на религиозной почве, так как они не состояли в англиканской церкви (были так называемыми «диссентерами»), поэтому были исключены из многих сфер общественной жизни.
Новая беда грянула в ноябре 1810 года, когда Ричард Эннинг, страдавший от последствий туберкулёза и нескольких опасных падений при сборе окаменелостей на скалах, умер в возрасте 44 лет. Молли, Джозеф и Мэри (ей тогда было всего одиннадцать) оказались вынуждены постоянно искать окаменелости, чтобы заработать себе на хлеб. Первая удачная находка подвернулась год спустя, когда Джозеф обнаружил удивительный череп длиной более 1,2 м, вмурованный в скалу, и выпилил его оттуда. Затем Мэри нашла оставшуюся часть скелета. Сначала они приняли эту находку за «ископаемого крокодила», поскольку у существа была длинная морда. Впоследствии её перекупали друг у друга богатые коллекционеры.
В 1814 году этот образец описал Эверард Хом, но учёный не разобрался в находке (рис. 14.1). Он классифицировал её как рыбу, поскольку существо было водоплавающим, а его позвонки напоминали рыбьи, однако признал, что тварь обладает и многими «рептильными» признаками. Хом решил, что перед ним «недостающее звено» из «великой цепи существ», пролегающей между рыбами и рептилиями. Однако он не считал, что одни из этих существ произошли от других (до появления этой идеи оставалось 40 лет). Затем, в 1819 году Хом решил, что перед ним — переходное звено между ящерицей и саламандрой-протеем, и назвал существо протеозавром.
 Рис. 14.1.
Рис. 14.1. На рисунке Уильяма Конибэра изображена первая известная окаменелость ихтиозавра (из книги
William Conybeare. Additional Notices on the Fossil Genera Ichthyosaurus and Plesiosaurus //
Transactions of the Geological Society of London, 2nd ser., 1 (1822))
В 1817 году Чарльз Кёниг (при рождении носивший имя Карл Дитрих Эберхард Кёниг), смотритель отдела естественной истории в Британском музее, неофициально назвал эту окаменелость ихтиозавром (от греческих слов, означающих «рыба» и «ящерица»), так как понял, что существо имеет признаки как рыб, так и ящериц. К маю 1819 года ему удалось приобрести это ископаемое для Британского музея, где оно находится по сей день. В 1822 году британский геолог Уильям Конибэр официально описал этот образец и назвал его, а также многие другие образцы ихтиозаврами. Теперь данное наименование относилось ко всем похожим экземплярам, найденным впоследствии. Название «протеозавр» за ненадобностью было упразднено.
Тем временем Джозеф Эннинг стал меньше заниматься сбором окаменелостей, так как поступил в ученики к драпировщику; одна Мэри продолжала кормить семью этим трудом. Большинство лучших своих находок она сделала в ветреные зимние месяцы, когда неспокойное море размывало скалы и на новой поверхности склонов оказывались свежие окаменелости. Эти месяцы были и самыми опасными, так как скала могла обрушиться в любой момент, а волны — смыть собирателя, если тот неправильно подсчитает время отлива. Вот как писала об Эннинг бристольская газета Mirror в 1823 году:
Эта неутомимая женщина годами ежедневно отправлялась на поиски важнейших окаменелостей при каждом отливе, проходила по много миль под нависающими скалами Лайма, чьи обрушившиеся массы — и есть цель её походов; ведь только эти скалы содержат ценнейшие осколки былого мира, которые в момент падения должны были застыть здесь, постоянно рискуя быть раздавленными теми фрагментами, что остались в подвешенном состоянии выше них, либо рассыпаться под ударом возвращающегося прилива. Именно трудам этой женщины мы обязаны практически всеми изящными образцами ихтиозавров, хранящихся сегодня в великих коллекциях.
Несколько раз Эннинг была на волосок от гибели. В октябре 1833 года её чуть не похоронило заживо под оползнем, унёсшим жизнь её бессменного спутника — чёрно-белого терьера по кличке Трей (рис. 14.2). Позднее, в том же году Мэри писала подруге Шарлотте Мёрчисон: «Ты, возможно, посмеёшься, если я скажу, что гибель моего старого верного пса довольно потрясла меня. Та скала, которая рухнула на него и мгновенно убила, прямо у меня на глазах, буквально под ногами... можно сказать, что мгновением раньше та же судьба могла постигнуть и меня».
 Рис. 14.2.
Рис. 14.2. Единственный известный портрет Мэри Эннинг, на котором она изображена с киркой, мешком для сбора образцов, в громоздком платье. Рядом лежит терьер Трей, впоследствии погибший под оползнем во время работы на скалах (иллюстрация из фонда Wikimedia Commons)
Но её усердие было вознаграждено. В 1823 году она нашла первый цельный образец длинношеего плезиозавра (см. рис. 15.5), который шокировал всё английское научное сообщество. Год спустя Мэри нашла ископаемого птерозавра — это был первый экземпляр, найденный за пределами Германии. Она собирала многочисленных ископаемых рыб, описанных другими учёными, а также множество аммонитов и других моллюсков. Эннинг обнаружила доказательства того, что у белемнитов, чьи раковины по форме напоминали пулю, имелся чернильный мешок — таким образом, эти существа оказались ископаемыми головоногими. Она поняла, что вещество, которое люди называют безоаровым камнем, на самом деле представляет собой фоссилизированные экскременты. Эту идею Уильям Бакленд позже выдал за собственную и изложил её в публикации, где назвал такие окаменелости «копролитами». Хотя Эннинг и была сравнительно малообразованной, она читала все научные статьи, какие могла отыскать, причём часто самостоятельно их переписывала и перерисовывала подробные иллюстрации. В 1824 году леди Гарриет Сильвестр так писала о ней:
Самое поразительное в этой молодой женщине — она столь досконально разобралась в науке, что лишь только найдя новую кость, уже знает, какому существу эта кость принадлежит. Она закрепляет кости на раме, покрытой цементом, а затем срисовывает их отпечатки... Это подлинно чудесный образец божественной благодати, что бедная, невежественная девушка оказалась столь одарённой, что чтением и практикой смогла обрести достаточные знания, чтобы беседовать с профессорами и другими учёными мужами на научные темы, причём все эти собеседники признают, что она понимает науку лучше, чем кто-либо в этом королевстве.
К 1826 году, когда Эннинг было всего двадцать семь, она скопила достаточно денег, чтобы открыть собственный магазин, куда обращались почти все известные геологи и палеонтологи, желавшие её посетить и приобрести найденные ею окаменелости. Среди этих учёных, в частности, были Луи Агассис, Уильям Бакленд, Уильям Конибэр, Генри де ла Беш, Чарльз Лайелл, Гидеон Мантелл, Родерик Мёрчисон, Ричард Оуэн и Адам Седжвик. Американские коллекционеры создавали собственные музеи, где выставляли окаменелости Мэри Эннинг, а представители монарших семей из нескольких стран покупали у неё лучшие образцы.
Несмотря на высокое мнение о Мэри Эннинг, британские джентльмены, заложившие основы современной геологии в начале XIX века, не принимали эту женщину на равных из-за её неблагородного происхождения и диссентерских религиозных взглядов. Позже она перешла в англиканскую церковь, чтобы избавиться от этой помехи. Все её великолепные находки описали покупавшие их богатые господа, почти не отмечавшие заслуг Мэри по сбору и препарированию окаменелостей. Ни одна из её идей не была опубликована при жизни, поскольку сама она не имела возможности напечататься. Мэри Эннинг умерла в 1847 году от рака груди, но к тому времени члены Британского геологического сообщества начали отдавать ей должное. Они собирали деньги, чтобы облегчить ей последние месяцы жизни и оплатить расходы на погребение, установили в её родной церкви витраж в её честь, а также произносили траурные речи о ней на своих собраниях (такой чести удостаивались лишь члены общества). Чарльз Диккенс даже написал о ней статью. Многие полагают, что стихотворение-скороговорка «Она продаёт ракушки на морском берегу...» рассказывает именно о Мэри Эннинг.
В настоящее время Эннинг считается не только первой и величайшей женщиной-палеонтологом, но и одной из основоположниц палеонтологии. Её открытия изменили представления о мире, в котором она родилась. К 1830-м люди стали серьёзно задумываться о подоплёке, связанной с вымиранием ихтиозавров и плезиозавров, рассуждать о тёмном и гибельном «допотопном мире», в морях которого плавали такие чудовища (рис. 14.3). Несколько лет спустя к этой драме добавились динозавры. Хотя в 1820-е и ранее барон Жорж Кювье показал, что мамонты, мастодонты и гигантские ленивцы должны были вымереть, именно вымирание таких крупных животных, как ихтиозавр и плезиозавр, наконец сподвигло учёных отказаться от буквальной интерпретации Книги Бытия. Они с ужасом взирали на причудливый допотопный мир, присматриваясь к зловещему блеску в глазах ихтиозавра. Некоторые, в частности Лайелл, полагали, что Земля развивается циклично и вымершие животные когда-нибудь возродятся (рис. 14.4). Но в конечном итоге большинство учёных были вынуждены отказаться от идеи совершенного творения неизменного мира и усомниться в реальности Всемирного потопа. Сама того не желая, Мэри Эннинг, набожная и скромная женщина, старавшаяся прокормиться сбором и продажей окаменелостей, заложила основы колоссальной революции в научном мышлении, прежде чем безвременно скончалась в возрасте 47 лет.
 Рис. 14.3.
Рис. 14.3. Ихтиозавр и два плезиозавра дерутся на побережье Лайм-Реджис. Рисунок Генри де ла Беша (1830). Это одно из первых изображений известной доисторической сцены, считается первым произведением в жанре палеоарт. Литографии этой сцены поступали в продажу, чтобы собрать деньги для Мэри Эннинг. В начале XIX века плезиозавры и ихтиозавры были широко известны, а динозавры ещё не открыты (из книги Henry De la Beche, Duria Antiquior — A More Ancient Dorset (London, 1830))
 Рис. 14.4.
Рис. 14.4. «Ужасные перемены» — сатирический рисунок, сделанный Генри де ла Бешем в 1830 году. Профессор Ихтиозавр читает лекцию о странном существе, жившем после последнего творения (показывает человеческий череп). Чарльз Лайелл был одним из последних геологов, отрицавших возможность массового вымирания и считавших, что история Земли циклична, а вымершие виды через какое-то время воплощаются в следующих ипостасях. В конечном итоге Лайеллу пришлось признать, что палеонтологическая летопись свидетельствует о последовательных изменениях и что вымершие виды не возрождаются (из книги Henry De la Beche, Duria Antiquior — A More Ancient Dorset (London 1830))
«Рыбоящеры»
Открытия Мэри Эннинг отворили дверь в мир удивительной группы животных—ихтиозавров. В начале XIX века динозавры были известны лишь по фрагментам зубов и челюстей и, следовательно, оставались малоизучены, пока в 1880-е годы не нашли их полные скелеты (см. главу 17). Зато найденные окаменелости ихтиозавра часто были полными или почти полными. Поэтому натуралисты быстро удостоверились, что эти животные действительно были рептилиями, хотя формой тела сильно напоминали китов или дельфинов благодаря так называемой конвергентной эволюции. Большинство ихтиозавров имели длинную узкую заострённую морду и множество острых конических зубов, которыми ловили плавающую добычу. У многих имелись большие глаза — видимо, чтобы лучше видеть в мутной воде; причём у некоторых видов глаза были защищены оболочкой из мелких костей, окружавших глазное яблоко и зрачок, так называемым склеротическим кольцом. На костях ихтиозавров, живших в конце юрского периода, есть следы кессонной болезни. Это свидетельствует о том, что животные могли нырять на большую глубину и часто страдали из-за необходимости надолго задерживать дыхание; при этом, когда они поднимались к поверхности воды, их кровь насыщалась пузырьками азота.
Голова ихтиозавра плавно переходила в туловище, как у многих водных животных, приобретающих обтекаемую форму тела из-за постоянного плавания. По последним оценкам, ихтиозавр мог плавать со скоростью 2 км/ч — чуть медленнее самых быстрых современных дельфинов и китов. На дельфинообразном теле ихтиозавра был спинной плавник (аналогичные плавники есть у дельфинов и рыб), который имел хрящевую, а не костную основу, и заметен только у экземпляров, у которых сохранились мягкие ткани. Передние конечности ихтиозавра превратились в ласты, состоявшие из десятков мелких костных дисков, сформировавшихся в результате «умножения» и «деления» костей пальцев на множество крошечных фрагментов. Задние конечности ихтиозавра превратились в гораздо более миниатюрные плавники (у китов и дельфинов они полностью утрачены), которые, вероятно, не слишком помогали для разгона при плавании. Задняя часть тела переходила в «рыбий» хвост с плавниками, ориентированными в вертикальной плоскости, то есть ихтиозавр плавал, двигая хвостом из стороны в сторону (как делает большинство костистых рыб).
Последние задние позвонки резко изгибались книзу в виде «завитка», поддерживавшего нижний хвостовой плавник; верхний хвостовой плавник поддерживался не костью, а хрящом. На заре изучения ихтиозавра учёные ломали голову над тем, зачем понадобился такой «завиток» хвостовых позвонков, и полагали, что это может быть механический дефект, возникающий, например, из-за усыхания и скукоживания сухожилий. Однако Ричард Оуэн верно заключил, что это остаток двухлопастного хвостового плавника. Его догадка подтвердилась в конце XIX века, когда в Германии обнаружили удивительное местонахождение Хольцмаден. На здешних окаменелостях сохранились тёмные отпечатки мягких тканей. Таким образом, палеонтологи впервые смогли оценить, для чего служила верхняя половина хвостового плавника у ихтиозавра, а также рассмотреть контуры спинного плавника (не имеет костной поддержки и, следовательно, обычно не видим).
Нам многое известно о палеобиологии ихтиозавра, поскольку имеются многочисленные и хорошо сохранившиеся, полностью сочленённые скелеты, порой с очертаниями мягких тканей и даже с содержимым желудка. Считается, что большинство ихтиозавров кормились как дельфины или киты: быстро ловили длинной зубастой пастью плавающую добычу (кальмаров, белемнитов, аммонитов, рыбу и т. п.). Это подтверждает и содержимое их желудков. У некоторых ранних ихтиозавров были затупленные размалывающие зубы для поедания моллюсков, другие имели беззубые «клювы» и, как считается, всасывали свою пищу (сегодня так питаются многие рыбы). Подтверждается, что крупные особи ели более мелких ихтиозавров. Многие хищники нападали на ихтиозавров, оставляя отметины на их мордах и костях. У некоторых ихтиозавров была короткая нижняя челюсть, а верхняя превращалась в длинный бивень, позволявший животному нападать на косяки рыбы (так делают меч-рыба и рыба-парусник).
С самого начала учёные спорили, как полностью водное животное могло выходить на сушу, особенно для откладывания яиц (подобно морским черепахам)? Учитывая, что плавники ихтиозавра были слишком малы, чтобы с их помощью животное могло вылезти на берег и ползти по песку. Затем в Хольцмадене были найдены окаменелости, запечатлевшие выход детёныша хвостом вперёд из полости, которая могла представлять собой родовые пути самки. Эти находки подтвердили давнюю догадку учёных: ихтиозавры были живородящими, размножались внутриутробным оплодотворением и никогда не откладывали яиц (точно так же беременные китихи и дельфинихи проводят всю жизнь в море, вынашивая детёнышей до их первого вздоха).
Короче говоря, ихтиозавр — удивительный пример конвергентной эволюции с дельфином. Строение его тела напоминало дельфинье, но эта была рептилия, а не млекопитающее, причём обладавшая некоторыми фундаментальными отличиями. Откуда взялись такие специализированные существа?
Происхождение ихтиозавров
Как и в случае с плезиозаврами (см. главу 15), мы располагаем превосходной серией ископаемых переходных звеньев, демонстрирующих происхождение ихтиозавра (рис. 14.5). Первым идёт нанчаньгозавр, живший в раннем триасе на территории Китая. У него было типичное рептильное тело, хотя уже имелась удлинённая морда, как у ихтиозавров. Когда это животное впервые описали, палеонтологи не были уверены, к какой группе его отнести, поскольку он обладал многочисленными примитивными рептильными признаками, но по форме черепа близок ихтиозаврам.
 Рис. 14.5.
Рис. 14.5. Происхождение ихтиозавра от более примитивных рептилий на протяжении триасового периода (© Ryosuke Motani)
Затем идёт утатсузавр, живший в раннем триасе на территории Японии. У него было обтекаемое торпедообразное тело, как у ихтиозавра, но передние и задние конечности примитивные, ещё не превратились в плавники. Длинная «ихтиозавровая» морда, но без нижнего завитка на хвостовом плавнике, а лишь плавный изгиб, позволяющий предположить, что в верхней части хвостового плавника имелись небольшие лопасти. Следующее звено — чаухозавр, живший в начале триаса на территории Китая. Череп у него был как у настоящего ихтиозавра, с коротким носом, простыми зубами и большими глазами. Но его крепкие лапы только начинали приобретать первые признаки типичных ихтиозавровых ласт, а на хвосте появился едва заметный завиток, поддерживающий верхнюю лопасть хвостового плавника.
Ещё более специализированным был миксозавр, обитавший в среднем триасе на территории Германии и в других местах (см. рис. 14.5). Это классическое переходное звено на полпути между развитыми ихтиозаврами и их более примитивными предками. Его тело во всём напоминало дельфинье; он имел длинную морду, большие глаза и спинной плавник, как большинство ихтиозавров. Передние и задние конечности — уже настоящие плавники, но количество пальцев на них не увеличилось. Хвост ещё более заметно загнут книзу, чем у чаухозавра, а в верхней части хвостового плавника осталась совсем небольшая лопасть. Калифорнозавр, живший в позднем триасе, специализирован ещё заметнее, у него сильнее видоизменились передние плавники и появляются первые признаки редукции задних плавников, а также ещё выразительнее становится изгиб хвоста книзу. Вероятно, на хвостовом плавнике у него была верхняя лопасть, хотя об этом сложно судить, поскольку нет хорошо сохранившихся экземпляров животного.
Все эти промежуточные звенья подводят нас к полноценному ихтиозавру юрского периода, примером которого является офтальмозавр (см. рис. 14.5): длинная зубастая морда; компактный череп с большими глазами, защищёнными склеротическими кольцами; полностью обтекаемое тело со спинным плавником; большие передние ласты с дополнительными костями в пальцах; маленькие задние ласты, также с дополнительными костями и резкий изгиб книзу, характерный для хвостового отдела позвоночника и указывающий, что верхняя и нижняя лопасть хвостового плавника были полностью симметричны. Именно такую тварь Мэри Эннинг извлекла на свет в 1811 году, а сегодня эту эволюционную линию можно проследить вплоть до существ, практически не напоминающих ихтиозавра.
Киторептилии триасового периода
До сих пор мы рассматривали «средних» ихтиозавров, большинство из которых имели не более 3–5 м в длину. Но существовали ихтиозавры, по размерам сравнимые с китами. Самым эффектным из них был шонизавр.
Первого ископаемого шонизавра обнаружили близ посёлка Берлин, на юге штата Невада, где теперь располагается знаменитый парк «Берлин-Ихтиозавр» (рис. 14.6). Это место находится в каньоне Уэст-Юнион на высоте 2133 м в Шошонских горах. Путь туда на автомобиле занимает шесть часов (на север) от Лас-Вегаса или три часа (на восток) от Рено. Настоящий медвежий угол. В состав парка входит не только местонахождение окаменелостей, но и городок Берлин, в настоящее время заброшенный. Первые рудокопы, жившие в этих местах, знали об ископаемых аммонитах и двустворчатых моллюсках, а некоторые находили огромные кости и даже строили из костей ихтиозавра очаги! В 1928 году Симон Мюллер из Стэнфордского университета догадался, что эти кости принадлежали ихтиозавру, но у него не было средств, чтобы их собирать или изучать.

 Рис. 14.6.
Рис. 14.6. Парк «Берлин-Ихтиозавр» близ Гэббса, штат Невада: (
А) входная площадка с барельефом шонизавра в натуральную величину; (
Б) обширное ископаемое ложе, усыпанное костями, внутри главного здания (фотографии предоставлены Lars Schmitz)
Прошло ещё 24 года, и Маргарет Уит из города Фаллон, штат Невада, собрала несколько никого не интересовавших окаменелостей и отправила их Чарльзу Л. Кэмпу, работавшему в Палеонтологическом музее Калифорнийского университета в городе Беркли. Кэмп заинтересовался и сам посетил залежи окаменелостей в 1953 году, после чего решил организовать серьёзные раскопки и хорошенько изучить ископаемые. После этой поездки Кэмп писал в своём полевом дневнике:
Си Мюллер говорит, что находил эти останки ихтиозавров в 1929 и 1930 годах, тогда пытался заинтересовать нас ими. Затем миссис Уит рассказала мне о них в прошлом сентябре и упомянула, что позвонки были очень крупными (до 1 фута
[4] в диаметре и 21 фунт
[5] весом). Мы взобрались по южному склону... и осмотрели материал, который миссис Уит смогла очистить своей кисточкой. Это был ряд из шести или более позвонков в твёрдом известняке, и ещё многие такие позвонки находились ниже. Позвонки были просто чудовищные — крупнее, чем у каких-либо ранее известных ихтиозавров, и более поздние по возрасту, чем позвонки среднетриасового цимбоспондила (описанного Лейди).
Каждое лето с 1954 по 1957 год Кэмп вместе с Сэмюэлом Уэллсом и группой сотрудников музея разворачивали серьёзные работы на этом месте (вторую серию экспедиций Кэмп предпринял в 1963–1965 годах). Им удалось извлечь один почти полный скелет, который сегодня выставлен в музее штата Невада в Лас-Вегасе (рис. 14.7,
А). Но большую часть ложа они оставили нетронутым, только очистили и обработали кости, чтобы они были видны лучше, чем ранее, в основном засыпанные землёй.

 Рис. 14.7.
Рис. 14.7. Шонизавр: (
А) полностью собранный скелет, выставленный в музее штата Невада в Лас-Вегасе; (
Б) реконструкция внешнего вида. ((
А) фотография автора; (
Б) иллюстрация предоставлена Nabumichi Tamura)
За работой Кэмп и его команда наблюдали несколько ослепительных вспышек и слышали, как рвались ядерные заряды на Невадском испытательном полигоне всего в 240 км южнее. Вот как Кэмп описывал 15 мая 1955 года, когда была взорвана бомба мощностью 28 килотонн:
Сегодня в 5 утра состоялся 14-й ядерный взрыв, в 200 милях
[6] отсюда. Я сидел в постели и видел фиолетово-розовую вспышку, продлившуюся долю секунды. Примерно через 13 минут до нас донёсся низкий громоподобный рокот, и земля дрогнула. Волна накатывала двумя или тремя крещендо. Через 3–5 минут послышался другой, приглушённый шум, будто где-то далеко, но очень сильно рычали львы.
К счастью, радиоактивные осадки от этих испытаний сносило к востоку, а не к северу, поэтому палеонтологи не пострадали. Кэмп дожил до 82 лет и в 1975 году умер от рака; за всю жизнь явно не подвергался сильному облучению. А вот постоянным жителям городка Джордж в штате Юта повезло меньше.
Концентрация скелетов в этом месте шокирует: здесь находится не менее 40 особей. Кэмп сначала полагал, что ящеров могло вынести на берег во время отлива, подобно тому как сегодня океан выбрасывает китов. Но более поздние исследования, проведённые Дженнифер Хоглер, показали, что эта часть формации Ланинг (верхний триас (около 215–217 млн лет)) представляет собой глубоководные отложения. Причина же, по которой многочисленные утонувшие скелеты ихтиозавров были нетронуты, по-прежнему остаётся загадкой. Отсутствие беспозвоночных-обрастателей, относительно неповреждённые кости и цельные скелеты позволяют предположить, что в очень глубоких местных стоячих водах не могли существовать не только падальщики, но и другие организмы.
По размерам шонизавр был сравним с большим китом — около 15 м в длину; с длинной беззубой мордой (зубы имелись только у молодых особей). Некоторые учёные полагают, что ему было несвойственно стремительно плавать в погоне за добычей (рис. 14.8; см. также рис. 14.7,
Б). Он всасывал пищу, плавая в океане; именно так поступают большие киты и китовые акулы, питающиеся планктоном. У шонизавра было круглое объёмное тело, относительно крупные грудные и брюшные плавники, целиком состоявшие из огромных круглых фаланг, возникающих, когда кости пальцев превращаются в ласты (гиперфалангия). У него явно не имелось спинного плавника и, подобно многим другим триасовым ихтиозаврам, на хвосте — лишь небольшая верхняя лопасть и небольшой загиб книзу на кончике хвостовых позвонков; не такой резкий завиток, как у ихтиозавров юрского периода.
 Рис. 14.8.
Рис. 14.8. Сравнение размера ихтиозавров Shonisaurus popularis и S. sikanniensis
Кэмп потратил немало времени, чтобы завершить изучение этих окаменелостей. Он назвал животное шонизавром в честь Шошонских гор и живущего там индейского племени шошонов; видовое имя выбрал тривиальное —
popularis (обыкновенный). В конце 1950-х Уит, Кэмп и Уэллес осознали, что гигантское существо заслуживает статуса главной окаменелости штата Невада. По-настоящему эффектный шонизавр встречается только в Неваде, и он более харизматичен, чем большинство найденных здесь окаменелостей. Лоббирование длилось несколько десятков лет, и высокий статус шонизавра был официально прописан в 1984 году.
В 2004 году ныне покойная Бетси Николс обнаружила и описала ещё более крупного верхнетриасового шонизавра (210 млн лет), найденного в формации Пардоне на территории Британской Колумбии (рис. 14.9). Это животное, названное
Shonisaurus sikanniensis, достигало более 21 м в длину — крупнее большинства современных китов! У этого шонизавра тоже была беззубая морда, большое грузное тело с длинными узкими грудными и брюшными плавниками, не было спинного плавника, а хвостовой плавник имел сверху маленькую лопасть. Со времени открытия
Shonisaurus sikanniensis некоторые учёные классифицируют его как представителя шастазавров — гораздо более мелких ихтиозавров, живших в нижнем триасе и обнаруженных в Калифорнии. Однако новейший анализ, проведённый в 2013 году, подтверждает версию о том, что перед нами — представитель гигантского вида шонизавров.
 Рис. 14.9.
Рис. 14.9. Гигантский шонизавр (или шастазавр?) из Британской Колумбии, выставленный в Королевском Тирреловском палеонтологическом музее в Драмхеллере (провинция Альберта) (фотография предоставлена Королевским Тирреловским палеонтологическим музеем)
Лучше один раз увидеть!
Окаменелости, найденные Мэри Эннинг, экспонируются в Естественнонаучном лондонском музее, а также в других, например в Музее наук о земле им. Седжвика в Кембриджском университете. Во многих музеях США есть великолепные экземпляры ихтиозавров; таковы, в частности, Американский музей естественной истории (Нью-Йорк), Музей естественной истории им. Карнеги (Питтсбург), Музей естественной истории под открытым небом (Чикаго) и Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия). Во многих немецких музеях представлены ископаемые ихтиозавры, добытые в Хольцмадене. В частности, это Музей естествознания в Берлине (музей Гумбольдта), Музей природы Зенкеберг (Франкфурт-на-Майне), Мюнхенский палеонтологический музей и Государственный музей естествознания (Штутгарт).
Чтобы попасть в парк «Берлин-Ихтиозавр» в штате Невада, едете по магистральному шоссе 361 штата Невада до Гэббса. Сначала поворачиваете к востоку, на шоссе 844, ведущее к Грэнтсвиллу, оттуда — дальше на восток по грунтовой дороге, пока не доберётесь до места. Практически полный скелет
Shonisaurus Popularis выставлен в Музее штата Невада, в заказнике Спрингс близ Лас-Вегаса. Гигантский
Shonisaurus sikanniensis экспонируется в Королевском Тирреловском палеонтологическом музее в Драмхеллере (провинция Альберта).
Дополнительная литература
Callaway, Jack, and Elizabeth L. Nicholls, eds. Ancient Marine Reptiles. — San Diego: Academic Press, 1997.
Camp, Charles L. Child of the Rocks: The Story of Berlin-Ichthyosaur State Park. — Nevada Bureau of Mines and Geology Special Pubhcation 5. Reno: Nevada Bureau of Mines and Geology, with Nevada Natural History Association, 1981.
Ellis, Richard. Sea Dragons: Predators of Prehistoric Oceans. — Lawrence: University Press of Kansas, 2003.
Ending, Shelley. The Fossil Hunter: Dinosaurs, Evolution, and the Woman Whose Discoveries Changed the World. — New York: Palgrave Macmillan, 2009.
Hilton, Richard P. Dinosaurs and Other Mesozoic Animals of California. — Berkeley: University of California Press, 2003.
Howe, S. R., T. Sharpe, and H. S. Torrens. Ichthyosaurs: A History of Fossil Sea-Dragons. — Swansea: National Museum and Galleries of Wales, 1981.
Wallace, David Rains. Neptune’s Ark: From Ichthyosaurs to Orcas. — Berkeley: University of California Press, 2008.
15. Гроза морей
Величайший морской монстр
Кронозавр
В мезозойскую эру не было настоящих морских змеев, но плезиозавры походили на них как две капли воды. Плезиозавр — рептилия, вернувшаяся в воду (видимо, эта идея показалась им удачной). Поскольку плезиозавры едва ли разбирались в плавании, они перелопачивали воду всеми четырьмя лапами, а не разгонялись хвостом, как более смышлёные морские животные (тот же ихтиозавр, пользовавшийся ластами в качестве балансира и руля). Всё этот плезиозавр неправильно делал! Он был слишком медлителен, чтобы ловить рыбу, — так достраивал себе шею позвонок за позвонком, пока она не стала длиннее всего тела... Пугать в море было некого, кроме рыбы, да и незачем. И душа к этому делу не лежала. Поскольку плезиозавр был так нескладен, жилось ему трудно. Приходилось выползать на берег откладывать яйца и всё такое. (Ихтиозавры — они всю жизнь проводили в воде и рожали живых малюток. Это легко делается, если знаешь как.)
Уилл Каппи. «How to Become Extinct»
Океаны в глуши
Сегодня австралийская глушь — полупустыня, покрытая сухим колючим кустарником, заросли которого тянутся на сотни километров. Редкие ливни обрушиваются стеной, и биллабонги (сухие старицы) стремительно превращаются в озёра. Большинство здешних растений приспособились бурно расти в течение считанных недель, когда достаточно влажно, а затем переносить засуху основную часть года. Высокие эвкалипты дают небольшую тень, но они постоянно истекают соком и сбрасывают длинные узкие листья, а также большие полоски коры. Вся экосистема приспособлена к засухе. Леса жарко пылают во время пожаров, которые случаются всё чаще, а сочная растительность вспыхивает, как пакля. Животные из австралийской глуши тоже хорошо приспособлены к засухе: это касается и крупнейших травоядных, кенгуру, и норных вомбатов, и коал, живущих в листве эвкалиптов.
Сложно представить, что этот выжженный ландшафт когда-то выглядел иначе, но горные породы на большей части Австралии убедительно свидетельствуют, что раньше тут были совсем иные условия. Эти породы — известняки, откладывающиеся на дне мелких морей, которые покрывали большую часть Австралии и большинства других континентов. В середине эры динозавров (начало мелового периода, 125–100 млн лет назад) на Земле царил глобальный парниковый климат. Грандиозные извержения подводных вулканов, спровоцированные мантийными суперплюмами, накачивали атмосферу колоссальными объёмами углекислого газа. Благодаря высокой концентрации парниковых газов в атмосфере на нашей планете было гораздо теплее, чем когда-либо. Учёные полагают, что в воздухе могло быть до 2000 единиц углекислого газа на миллион (ppm), тогда как сегодня это значение составляет более 400 ppm. Естественно, лёд на такой тёплой планете не мог существовать подолгу, поэтому не было ни полярных шапок, ни горных ледников, вообще никакого льда (к сожалению, режиссёры многих современных фильмов о динозаврах, вероятно, не знают об этом факте, и на фоновых пейзажах часто мелькают заснеженные горы).
Кроме того, основные континенты стремительно удалялись друг от друга, хотя незадолго до того образовывали единый суперконтинент Пангею. Быстрый спрединг морского дна не только приводил к насыщению атмосферы парниковыми газами, но оказывал и другие эффекты. При очень быстром спрединге общий объём срединноокеанического хребта гораздо выше, так как этот хребет горячий и более широкий, чем при медленном спрединге. Напротив, при сравнительно медленном спрединге хребет долго остывает, поэтому его склоны более обрывистые, а сам хребет тоньше. Поскольку срединноокеанические хребты были распластаны, сами океаны были мельче, и их воды распространялись на свободное место — заливали континенты. Кроме того, существованию сравнительно мелких океанов и общему подъёму уровня моря способствовало то, что вокруг подводных вулканов скапливались гигантские плато из лавы, а чем теплее вода, тем сильнее она расширяется (из-за последнего фактора уровень моря растёт и сегодня).
В результате мелкие моря подтопили почти все континенты раннего мелового периода. Некоторые территории были затоплены ещё в конце юрского периода, когда началось глобальное потепление. Под водой оказалась большая часть не только Австралии, но и Европы. Мелкие моря, покрывавшие Европу, кишели новыми формами планктона — микроскопическими водорослями, так называемыми кокколитофоридами. При отмирании этого фитопланктона крошечные известковые оболочки водорослей опускались на дно, скапливались там и спрессовывались в виде огромных массивов камня, который мы именуем известняком или мелом. Сегодня эти меловые моря превратились в сушу. Речь идёт не только о таких знаменитых местах, как белые скалы Дувра, но и о побережье Северной Франции, Бельгии и Голландии.
На территории Северной Америки тоже было огромное мелкое море, простиравшееся на месте современных Великих Равнин. Оно соединяло Мексиканский залив и тёплый Арктический океан. Почти все штаты и провинции на территории равнин — Техас, Оклахома, Небраска, Южная и Северная Дакота, Альберта и Саскачеван — покрыты огромными пластами сланцев, скопившихся в мелководных морях, а также известняком и мелом. На меловых отложениях Ниобрара в Западном Канзасе можно собрать огромное количество морских окаменелостей, в том числе гигантских морских рептилий, огромных рыб и морских черепах, а также разнообразных беспозвоночных, от аммонитов до двустворчатых моллюсков более 1,7 м в поперечнике.
Морское чудовище на краю света
Более века назад об этом ещё никто не знал. В 1899 году человек по имени Эндрю Кромби из городка Хьюэнден в австралийском штате Квинсленд обнаружил вблизи от своего дома обломок кости с шестью коническими зубами. В конце концов этот фрагмент оказался в музее Квинсленда, директор которого, Хебер Лонгмэн, в 1924 году назвал животное, которому принадлежала эта кость,
Kronosaurus queenslandicus (родовое название состоит из имени Кронос и греческого слова, в переводе означающего «ящерица»; видовое дано в честь региона, где его нашли). В греческой мифологии Кронос был одним из титанов. Он сверг своих родителей, Урана и Гею, а затем съел всех своих детей, кроме одного, чтобы те не могли лишить его власти. Рея, супруга Кроноса, спасла новорождённого сына Зевса, подсунув Кроносу вместо ребёнка камень омфал, завёрнутый в пелёнки. Возмужав, Зевс одолел Кроноса и заставил изрыгнуть проглоченных детей, ставших первыми олимпийскими богами и богинями. После этого Зевс заточил Кроноса в Тартаре. Очевидно, подбирая название для животного, Лонгмэн хотел подчеркнуть его титанические размеры. Впоследствии учёные из Квинслендского музея вернулись на то место, где Кромби нашёл первую кость, и добыли новый материал, в том числе частично сохранившийся череп кронозавра.
Узнав об этом огромном существе, сотрудники Музея сравнительной зоологии при Гарвардском университете снарядили экспедицию в эти места. Группу возглавил Уильям Шевилл, молодой аспирант-палеонтолог, закончивший обучение в Гарварде в 1927 году. Он и ещё шесть человек прибыли на раскопки в 1931 году. Шевилла описывали как очень сильного человека (в экспедицию он отправился в возрасте примерно 25 лет). Например, он прихватил с собой трёхкилограммовую кирку для откалывания известняка и, по рассказам, на ходу подбрасывал её и ловил. Впоследствии Шевилл стал экспертом по эхолокации и коммуникации китов, работал в Океанографическом институте Вудс-Хол. Путешественникам поручили собирать для музея любые образцы, имеющие естественнонаучную ценность. Томас Барбур, директор музея, выразился так: «Мы будем рассчитывать на экземпляры кенгуру, вомбата, тасманийского дьявола и тасманийского волка». Группа вернулась в Гарвард через год, доставив более 100 экземпляров окаменелостей млекопитающих и многие тысячи насекомых.
После возвращения первой гарвардской экспедиции в США Шевилл, оставшийся в Австралии, мобилизовал местных специалистов и организовал экспедицию по изучению нижнемеловых отложений в районе Ричмонда и Хьюэндена. Как сообщает австралийский палеонтолог Джон Лонг, Шевилл пригласил к участию сотрудников Австралийского музея (музей предложением не заинтересовался) и Квинслендского музея (в данном случае не нашлось средств на такое
предприятие или сотрудников, которые могли бы помочь).
В 1932 году группа добралась до долины Грампиан и Хьюэндена, где нашли переднюю часть черепа небольшого кронозавра. Затем от местного фермера Ральфа Уильяма Хаслэма Томаса учёные узнали, что на своём обширном участке Арми Даунз (8100 га) он нашёл какие-то огромные кости. Те явно пролежали в земле много лет, но были слишком тяжёлыми, чтобы их куда-то носить или собирать. В лучшем случае можно было отколоть зуб-другой молотом и долотом. Поэтому до прибытия гарвардцев этими костями никто особенно не интересовался. Учёные разбили лагерь под развесистой баугинией и регулярно охотились. Однажды вечером их навестила семья местных жителей — узнать, не нужно ли свежего мяса. Палеонтологи ответили: «Нет, спасибо, мяса у нас хватает» (питались кенгурятиной, поджаренной на жире эму, приправленной острым сыром и патокой).
Кости сидели в толстых и твёрдых известняковых конкрециях, большинство из них удалось извлечь, только пустив в ход динамит. Ассистент Шевилла по прозвищу «Маньяк» был спецом по взрывному извлечению из породы костей и их раскалыванию на более мелкие куски для удобной транспортировки. Большинство костей, остававшихся на поверхности, разрушились в результате выветривания; сохранились лишь те, что залегали в глубоких нодулях. Не хватало некоторых фрагментов задней части черепа, а также большей части позвоночника и рёберных, тазовых, а также плечевых костей.
В конце концов исследователи укомплектовали 86 деревянных ящиков с окаменелостями общим весом более 5,5 т, которые отправили в Бостон на пароходе
Canadian Constructor 1 декабря 1932 года. Затем тяжёлые блоки в кожухах поступили в лаборатории для разделки проб, расположенные на цокольном этаже. Там их начинали медленно обрабатывать гарвардские умельцы: Джим Дженсен по прозвищу Динозавр и Арни Миллер. Толстые известняковые конкреции предстояло долго и методично стачивать, а некоторые куски камня были настолько твёрдыми, что поддавались лишь отбойному молотку.
Сначала обработали череп; однако на титаническую работу по очистке всего скелета у учёных просто не было сил. В 1953 году окаменелостью заинтересовался один богатый спонсор: фамильной страстью его семейства было выслеживание морских змеев. Он выделил музею достаточно денег на обработку, и оставшуюся часть скелета удалось привести в порядок за три года. В 1959 году практически полный скелет кронозавра выставили на всеобщее обозрение в Гарварде (рис. 15.1). Ральфа Томаса, которому на тот момент исполнилось 93 года, пригласили в Гарвард на торжественное открытие экспозиции со скелетом — через 27 лет после того, как он впервые показал эту окаменелость музейщикам. Томас и Шевилл в слезах обнялись: каждый считал, что товарищ не пережил Вторую мировую войну.
 Рис. 15.1.
Рис. 15.1. Собранный скелет кронозавра, рядом с которым (для сравнения) стоит Рут, жена Альфреда Ромера. Экспонируется в Музее сравнительной зоологии при Гарвардском университете (фотография предоставлена библиотекой им. Эрнста Майра, Музей сравнительной зоологии в Гарвардском университете)
Сегодня в городе Ричмонд (штат Квинсленд) есть небольшой музей под названием «Уголок кронозавра». Перед входом там стоит бетонный макет кронозавра в натуральную величину — так могло выглядеть это животное в начале мелового периода (рис. 15.2).
 Рис. 15.2.
Рис. 15.2. Уголок кронозавра. Ричмонд, штат Квинсленд, Австралия (фотография предоставлена музеем «Уголок кронозавра»)
С тех пор как кронозавра открыли в Австралии, он был найден ещё в одном месте — в Колумбии. В 1977 году крестьянин из Моникиры, возделывая поле, наткнулся на огромный валун. Присмотревшись к нему, пахарь заметил окаменелость. Он написал об этом в колумбийские научные организации и стал её извлекать. Оказалось, что в камне был почти полный скелет кронозавра — одна из лучших окаменелостей, найденных в Колумбии. В 1992 году палеонтолог Оливер Гампе описал это животное как новый вид,
Kronosaurus boyacensis.
Царь морских чудовищ
Кронозавр был поистине удивительным существом. Его череп имел почти 3 м в длину (рис. 15.3), передние ласты достигали 3,3 м, а общая длина составляла около 12,8 м. Однако новейшие исследования показывают, что палеонтологи, реконструируя недостающие фрагменты скелета, могли составить слишком много позвонков. Вероятно, общая длина животного ближе к 10 м. Экземпляр из Музея сравнительной зоологии занимает в галерее целую стену, от одного его вида захватывает дух (см. рис. 15.1)! Сын Джима Дженсена рассказывает, что, подвешивая скелет на стену, тот воспользовался системой занавесок и другими ухищрениями, мастерски спрятав железные стержни и подпорки, приваренные к животному. Дженсен хотел изобразить кронозавра будто парящим в воздухе или плывущим в воде — как живое, динамичное существо. И ему удалось это сделать.
 Рис. 15.3.
Рис. 15.3. Реконструкция головы и тела кронозавра (иллюстрация предоставлена Nobumichi Tomura)
Кронозавр был крупнейшим представителем большого отряда морских рептилий, известных под общим названием «плезиозавры», к которым относятся два подотряда — плиозавровые и плезиозавровые. Строение тела у всех плезиозавров схоже, разные только головы и шеи. Эти животные активно плавали и бороздили моря мелового периода, загребая огромными передними и задними ластами. У плезиозавра был мощный плечевой и тазовый пояс, состоявший из нескольких костных брюшных пластин, к которым крепились сильные плавательные мускулы. Между плечевым и тазовым поясом располагалась сеть брюшных рёбер (
gastralia), дополнительно поддерживавших и усиливавших живот. У многих экземпляров в той части грудной клетки, где должен был находиться желудок, обнаружили гладкие камни: вероятно, кронозавры глотали их и использовали в качестве балласта. Кроме того, в желудках квинслендских кронозавров сохранились окаменелые остатки пищи, подтверждающие, что животные питались морскими черепахами и мелкими плезиозаврами. В тех же отложениях встречаются окаменелости огромных аммонитов и гигантских кальмаров; они тоже почти наверняка входили в рацион огромного хищника. Вдобавок в этих же отложениях были найдены плезиозавры-эромангазавры, на костях которых есть глубокие рубцы, вероятно, оставленные зубами кронозавра.
Если вы смотрели популярный сериал «Прогулки с динозаврами», вероятно, помните показанного там крупного европейского плезиозавра лиоплевродона. Это существо изобразили как 25-метрового монстра, охотящегося на динозавров и почти всех остальных обитателей юрского периода. Такое огромное существо по размеру вполне сравнимо с крупнейшими китами — например с синим (рис. 15.4).
 Рис. 15.4.
Рис. 15.4. Сравнение размеров плезиозавра лиоплевродона, кронозавра, большой белой акулы и синего кита. Также показаны явно преувеличенные в размерах «хищник Х» и гигантский лиоплевродон из телефильмов (рисунок Mary P. Williams)
К сожалению, как известно большинству палеонтологов, телевизионщики часто перевирают факты ради драматизма в сюжете. Я тоже знаю об этом не понаслышке, так как был консультантом на многих фильмах о доисторических хищниках, даже появлялся в кадре. Что бы я ни говорил сценаристам и продюсерам, они игнорировали мои слова, желая сделать ленту зрелищнее. Как только сценарий попадает в анимационную студию, о науке можно забыть! В большинстве случаев вся графика фантастичная. Имея лишь кости доисторических животных, мы не можем восстановить цвет их шкуры, не знаем, как именно они двигались или какие звуки издавали. «Истории» из этих документальных фильмов, описывающие контакты между древними животными, иерархию в их сообществах и т. д., также строятся на чистом вымысле (на основе исследований современных животных). К сожалению, обычно широкая аудитория знакомится только с подобным суррогатом палеонтологии, и у зрителей складывается ложное впечатление, что палеонтология нужна лишь для съёмки эффектных фильмов о вымерших животных, для демонстрации их масти, повадок, голоса. И ничто из показанного не подкреплено реальными научными данными.
На самом деле нет ни одного полностью сохранившегося ископаемого лиоплевродона, который мог бы иметь при жизни такие размеры. Среди имеющихся окаменелостей — несколько черепов и челюстей, а также другие отдельные кости. Крупнейший полный скелет, выставленный в Музее геологии и палеонтологии в городе Тюбинген, имеет длину всего 4,5 м. Новые методы оценки размера тела по черепу свидетельствуют, что крупнейшие черепа принадлежали животным около 5–7 м в длину, что близко не сравнимо с новыми оценками длины кронозавра (около 10 м).
В 2009 году на History Channel вышла сенсационная передача о доисторическом животном, которое авторы назвали «хищник Х» (см. рис. 15.4). Фильм был основан на открытии окаменелостей крупного плиозавра на арктическом архипелаге Шпицберген. В этой документальной ленте утверждалось, что животное имело длину 15 м и весило 5 т. Эту недостоверную информацию продублировали в 2011 году в одной из серий фильма «Планета динозавров». Обе ленты широко разрекламировали в других СМИ: упоминание о «величайшем хищнике всех времён» всегда привлекает внимание.
Когда экземпляры были, наконец, беспристрастно изучены и описаны, выяснилось, что их размеры сильно преувеличены. Окаменелости состоят лишь из нескольких фрагментов челюстей, нескольких позвонков и кусков ласт. Да, они большие, но размер животного нельзя уверенно оценить по столь скудному материалу. Телевизионщики, изначально раскручивавшие «хищника Х», снизили оценку его размеров до 12,8 м — примерно как у кронозавра. В настоящее время «хищник Х» получил официальное название
Pliosaurus funkei.
Мы можем говорить уверенно лишь о длине и размере кронозавра, поскольку его экземпляры достаточно полные. Всё остальное — чистые спекуляции и слухи, пока не будут найдены более полные окаменелости крупного плиозавра.
Морские длинношеи
Другая ветвь плезиозавров — более известные плезиозавроиды. Их самым знаменитым представителем является эласмозавр. В отличие от кронозавра и других плиозавровых, имевших большую тяжёлую морду и короткую шею, плезиозавроиды развивались в противоположном направлении: у них была исключительно длинная шея и крошечная голова. С тех пор как Мэри Эннинг открыла плезиозавра, первого представителя плезиозавроидов (рис. 15.5), были найдены многие другие подобные животные. Эти существа оказались не менее длинными, чем плиозавровые, но определённо не такими массивными. Как бы то ни было, они велики. Одним из крупнейших являлся эласмозавр; известны полные экземпляры этого существа длиной до 14 м, вес которых должен был составлять около двух тонн. В отличие от плиозавровых, плезиозавры, видимо, плавали медленно, неторопливо перебирая всеми четырьмя ластами.
 Рис. 15.5.
Рис. 15.5. Длинношеий плезиозавр Rhomaleosaurus cramptoni, найденный в Кеттлнессе (графство Йоркшир); экспонируется в Лондонском музее естественной истории (фотография из фонда Wikimedia Commons)
С тех пор как открыли ископаемых плезиозавров, палеонтологи их реконструировали, изобразив с длинной и гибкой змеевидной шеей; голова якобы могла вертеться в любом направлении. До сих пор на большинстве реконструкций плезиозавры выглядят именно так. Но последние оценки веса головы и шеи животного плюс малое количество шейных мускулов и ограниченная подвижность шейных позвонков указывают на то, что шея была не очень пластичной. Исследования говорят, что шея плезиозавра, вероятно, была полуподвижной и не могла сильно изгибаться, скорее напоминала не змею, а удочку. Кроме того, плезиозавры могли по-лебединому поднимать её над водой.
Исходя из того что шея плезиозавра не могла изгибаться как угодно и он был способен хватать добычу лишь в одном направлении, палеонтологи рассуждают о способах питания, не требующих гибкой шеи.
По одной из версий, плезиозавры могли затаиваться глубоко на дне и выслеживать ничего не подозревающую плывущую добычу. Плезиозавр мог просто засунуть голову в косяк рыбы, стаю кальмаров или аммониттов и ухватить жертву до того, как налетит ударная волна от массивного тела и все кинутся врассыпную. Огромные глаза животных тоже подтверждают эту идею.
Другая версия такова, что плезиозавры кормились со дна, перепахивая шеей морской ил и захватывая при этом добычу. У большинства плезиозавров были длинные зубы-клинья, направленные вперёд; это обычная адаптация к загарпуниванию рыбы и другой водяной добычи. Некоторые плезиозавры, например
Cryptoclidus и
Aristonectes, имели сотни крошечных зубов-карандашиков. Это позволяет предположить, что они могли выхватывать небольшие кусочки съестного из стай планктона либо с морского дна.
Другие учёные не уверены, что шея плезиозавра была малоподвижной. Они подчёркивают, что на окаменелостях большей частью отсутствуют мягкие ткани (особенно межпозвоночные хрящи), а с таким количеством позвонков шея должна была оставаться достаточно гибкой. Определённо, ей было далеко до змеиного тела, и она не могла изгибаться в форме буквы S, но сторонники этой версии полагают, что плезиозавр всё-таки мог согнуть шею в довольно тугую дугу, чтобы схватить добычу. В таком случае сложные повадки, диктуемые гипотезой «малоподвижной шеи», представляются менее вероятными.
Крупное тело, ласты прямо под брюхом, отсутствие связок между костями задних конечностей и позвоночником, а также другие черты плезиозавра не позволяют предположить, что он выползал на берег и откладывал яйца в песчаные ямки, как это делают морские черепахи. Однако многие художники по-прежнему изображают плезиозавра неуклюже распластавшимся по камням; его плавники слишком коротки, и ползти он не может. Тот факт, что плезиозавр всю жизнь проводил в воде, подтверждает описание недавно найденной ископаемой самки с эмбрионом в утробе. Таким образом, плезиозавры были живородящими и выводили потомство в море.
Происхождение морских чудовищ
Как возникли такие удивительные животные — плезиозавры? К счастью, мы располагаем замечательной палеонтологической летописью их происхождения от рептилий, которые даже не напоминают плезиозавров.
Древнейшим родственником плезиозавров был клаудиозавр — рептилия, обнаруженная в мадагаскарских породах, относящихся к пермскому периоду (270 млн лет назад) (рис. 15.6). Он очень похож на других примитивных пермских рептилий, за исключением того, что у клаудиозавра есть определённые ключевые черты черепа и нёба, характеризующие его как раннего представителя синаптозавров (
Euryapsida), к которым относятся и плезиозавры, и ихтиозавры. Видимо, он был полуводным и не имел грудины, которая могла бы мешать плавательным движениям передних лап. Соответственно, мог двигать и передними и задними лапами одновременно, а не попеременно, как делает ящерица. Его конечности — длинные и с очень длинными пальцами, между которыми, вероятно, были перепонки. Многие учёные отмечали, что пропорции конечностей и детали скелета клаудиозавра сближают его с галапагосской морской игуаной.

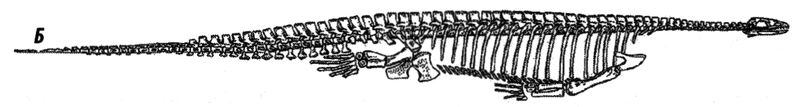

 Рис. 15.6.
Рис. 15.6. Ископаемые, позволяющие проследить переход от примитивных ящериц до настоящих плезиозавров: (
А) примитивная рептилия клаудиозавр, жившая в пермском периоде на Мадагаскаре; имела лишь некоторые признаки синаптозавров, но обладала короткой шеей, длинным хвостом, относительно крупными передними и задними конечностями, которые ещё не превратились в ласты; (
Б) нотозавр пахиплеврозавр, живший в триасовом периоде; имел сравнительно длинную шею, развитый хвост и конечности, приспособленные для плавания; (
В) примитивный настоящий плезиозавр — пистозавр, живший в триасовом периоде; имел удлинённую шею, удлинённый череп и сравнительно короткий хвост, конечности частично превратились в ласты; (
Г) более развитые плезиозавроиды: криптоклид (вверху) и гидротерозавр (внизу), с более длинными шеями, более маленькими головами, короткими хвостами и конечностями, полностью преобразованными в ласты (из книги
Robert L. Carroll. Vertebrate Paleontology and Evolution. — New York: Freeman, 1988. — Fig. 2-2, 12-4, 12-10, 12-12; иллюстрации предоставлены R. L. Carroll)
В триасовом периоде (250–210 млн лет назад) существовала большая группа примитивных водных рептилий под названием «нотозавры». Они тоже были сравнимы по размеру с большими ящерицами (менее метра в длину) и в основном похожи на клаудиозавра. Однако у них постепенно развивалась плезиозавровая длинная шея и вытянутая морда для ловли рыбы. В конечностях многие кости превратились в хрящи, что часто происходит у водных позвоночных. В плечевом поясе и тазовых костях нотозавров видны зачатки крепких пластинчатых костей, которые есть в плечевом и тазовом поясе плезиозавров.
Последним переходным звеном на пути к плезиозаврам была среднетриасовая рептилия пистозавр, найденная на территории Германии. У пистозавра имелся примитивный череп с простой мордой, зато нёбо было как у более высокоразвитых плезиозавров. Остальное тело — «переходный» вариант от ящериц к плезиозаврам: длинная шея, объёмистая утроба, хорошо развитые брюшные рёбра, а конечности походили как на ласты плезиозавра, так и на неспециализированные лапы нотозавров. Длинные кости передних и задних конечностей превратились в десятки дополнительных костей пальцев, которые, в свою очередь, стали дисковидными косточками в ластах плезиозавров.
Короче говоря, плезиозавры могут показаться странными и сильно специализированными, но мы можем проследить их родословную вплоть до обычных ящериц, ничуть не походивших на гигантских морских чудовищ.
Лох-несское чудовище?
Примерно с 1930-х не утихают слухи о том, что в шотландском озере Лох-Несс обитает чудовищная рептилия; позднее добавилась подробность: это доживший до наших дней плезиозавр. На поддержке мифа о лох-несском чудовище выросла целая индустрия, во многих телепередачах его пытаются подать в правдоподобном виде. Мы с Дэниэлом Локстоном продемонстрировали, что в реальности никакая рептилия в озере существовать не может (если, конечно, вы не думаете, что лох-несское чудовище — очень крупная рыба, например осётр). Тому есть разные причины, объясняемые сразу несколькими группами фактов:
•
Биологические. Климат в районе Лох-Несса слишком холодный, чтобы там могла долго существовать крупная холоднокровная рептилия. На самом деле в Шотландии обитает всего два вида ящериц и два вида змей, хотя на Земле царит сравнительно тёплый межледниковый климат. Знания лишь основ биологии достаточно, чтобы понять: в озере не может существовать всего
одно лох-несское чудовище — только их популяция, если ящеры действительно сохранились там на протяжении 65 млн лет, с тех пор как вымерли плезиозавры. Если бы популяция существовала, мы бы постоянно находили в районе озера множество костей и скелетов. На самом деле, в избытке встречаются кости, принадлежащие всем прочим животным, живущим и умирающим в Лох-Нессе или в любом другом крупном озере — но ни одного фрагмента костей Несси. Вдобавок озеро невелико и бедно ресурсами, чтобы поддерживать жизнедеятельность большой популяции хищных рептилий. Чем крупнее животное, тем больше охотничьи угодья, необходимые ему для пропитания, а озеро Лох-Несс маловато, чтобы там мог существовать хотя бы один такой монстр. Буквально каждый сантиметр дна озера неоднократно зондировали радаром и тралили, но ничего крупного не обнаружили.
•
Палеонтологические. Палеонтологическая летопись плезиозавров исключительно полная, равно как и палеонтологическая летопись морских позвоночных, живших в эру млекопитающих, после вымирания динозавров. Ни одна кость плезиозавра (а эти кости очень характерны и легко узнаваемы) не найдена в породах моложе 65 млн лет, притом что окаменелости других морских животных (акул, китов, морских львов, ламантинов) постоянно находят в таких местах, как холм Шарктус в Калифорнии (см. главу 9) и в скалах Колверт на берегу Чесапикского залива. Поскольку чем крупнее окаменелость, тем больше шансов, что она сохранится, это доказывает, что плезиозавры вымерли 65 млн лет назад.
•
Геологические. Лох-Несс представляет собой ледниковую долину, в которой ещё 20 тыс. лет назад лежал ледник толщиной в 1,6 км. Она была покрыта льдом на протяжении более 2,5 млн лет. Значит, чудовище пряталось в озере и миллионы лет было заперто под ледником, как по сценарию дешёвого фантастического фильма? Если нет, как оно туда попало? Если скрывалось в других местах, прежде чем переселиться в озеро, почему не найдены окаменелости? Кроме того, Лох-Несс находится в глубине острова и довольно высоко над уровнем моря, поэтому крупное морское животное не могло туда забраться. Тем более что плезиозавры не умели ползать по земле.
•
Культурные. Как продемонстрировали мы с Локстоном, плезиозавровая трактовка истории о лох-несском чудовище появилась сравнительно недавно. Она не встречается в расплывчатых древних свидетельствах о водном чудовище. В легендах оно именовалось «водяной лошадью» и ничуть не напоминало плезиозавра. На самом деле слух о плезиозавре пустил конкретный человек — Джордж Спайсер, увидевший плезиозавра в кинофильме о Кинг-Конге в 1933 году. С тех пор он и одна женщина по имени Олди Маккей заявляли, что видели чудовище, а газеты и прочие СМИ начали раздувать сенсацию.
• С момента появления этих репортажей стали распространяться многочисленные мистификации, подпитывавшие миф. Среди прочих подделок— «фотоснимок хирурга», классическое изображение Несси. После смерти мистификатора выяснилось, что он сфотографировал игрушечную субмарину, к которой приделал бутафорскую «голову». На других подобных снимках фигурировали, например, плывущие охапки сена, покрытые парусиной и верёвками. А знаменитый «плавник Несси» — просто зернистая фотография подводных пузырьков, претерпевшая изрядную обработку.
Короче говоря, существование лох-несского чудовища абсолютно невозможно с научной точки зрения и полностью опровергается любой из перечисленных групп фактов. Гипотеза подкрепляется лишь туманными «свидетельствами очевидцев», которые являются наислабейшими доказательствами в любом научном исследовании: человеческий глаз и мозг легко обманываются. Плезиозавры были удивительными существами. Было бы классно, если бы они до сих пор плавали в земных океанах, но, несмотря на живучесть мифа о лох-несском чудовище, плезиозавры действительно вымерли.
Лучше один раз увидеть!
Скелет
Kronosaurus queenslandicus по-прежнему является центральным экспонатом главного зала Музея сравнительной зоологии при Гарвардском университете (Кембридж, штат Массачусетс). В Австралии первые находки, связанные с кронозавром, экспонируются в Квинслендском музее (Южный Брисбен). Практически полный скелет
Kronosaurus boyacensis выставлен в том самом месте, где был найден: жители близлежащей деревни Вилла-де-Лейва выстроили прямо над ним Музей окаменелостей.
Во многих европейских музеях есть окаменелости плезиозавров. Так, находки Мэри Эннинг, сделанные в Лайм-Реджисе, выставлены в лондонском Музее естественной истории и Музее Лайм-Реджиса. Самый крупный череп
Pliosaurus kevani находится в Музее графства Дорсет (Дорчестер). Плезиозавры есть во многих немецких музеях, в частности в Музее естествознания в Берлине (музей Гумбольдта), Музее природы Зенкеберг (Франкфурт-на-Майне), Мюнхенском палеонтологическом музее и Государственном музее естествознания (Штутгарт). Единственный полный скелет лиоплевродона экспонируется в Музее геологии и палеонтологии города Тюбинген.
Во многих музеях США выставлены длинношеие эласмозавры. В частности, в Американском музее естественной истории (Нью-Йорк), Институте биоразнообразия и Музее естественной истории Канзасского университета (Лоуренс), Денверском музее природы и науки, Геологическом музее при Школе горнодобывающей промышленности и технологий (Рапид-Сити), Стернбергском музее естественной истории Университета Форт-Хейс (Хейс, штат Канзас). В Естественнонаучном музее округа Лос-Анджелес (город Лос-Анджелес) есть скелет эласмозавра, добытый в холмах Морено-Хиллс (меловой период) на территории Калифорнии и названный моренозавром. Скелет подвешен под потолком. Там же находится недавно описанный скелет беременной самки плезиозавра с эмбрионом в утробе. В Музее Отаго (город Данидин, Новая Зеландия) экспонируется плезиозавр, найденный в Новой Зеландии.
Дополнительная литература
Callaway, Jack, and Elizabeth L. Nicholls, eds. Ancient Marine Reptiles. — San Diego: Academic Press, 1997.
Ellis, Richard. Sea Dragons: Predators of Prehistoric Oceans. — Lawrence: University Press of Kansas, 2003.
Everhart, Michael J. Oceans of Kansas: A Natural History of the Western Interior Sea. — Bloomington: Indiana University Press, 2005.
Hilton, Richard P. Dinosaurs and Other Mesozoic Animals of California. — Berkeley: University of California Press, 2003.
Loxton, Daniel, and Donald R. Prothero. Abominable Science: The Origin of Yeti, Nessie, and Other Cryptids. — New York: Columbia University Press, 2013.
16. Чудовищный хищник
Крупнейший хищник
Гиганотозавр
Предлагаю отнести это животное к новому роду тираннозавр, подчеркнув тем самым, что он многократно превосходит по размеру любое плотоядное сухопутное животное, описанное по сей день... На самом деле, это животное — непревзойдённый образец эволюции крупных плотоядных динозавров; в общем, он удостаивается царственного и высокородного родового имени, которым я его нарекаю.
Генри Фэрфилд Осборн. «Tyrannosaurus and Other Cretaceous Carnivorous Dinosaurs»
Царь тиранических ящеров
Благодаря вековому пиару
Tyrannosaurus rex — пожалуй, самый известный и наиболее популярный динозавр. Он был открыт в бесплодных землях Хелл-Крик на территории штата Монтана известным собирателем окаменелостей Барнумом Брауном в 1900 году, а описан знаменитым палеонтологом Генри Фэрфилдом Осборном в 1905 году. Именно Осборн окрестил его запоминающимся именем, которое означает «царь тиранических ящеров»; не менее известно и сокращённое название
T. Rex. На самом деле, тираннозавр королевский — одно из немногих научных наименований, известных почти каждому (наверное, оно даже более известно, чем название нашего собственного вида
Homo sapiens). Браун нашёл всего пять скелетов, и к тому времени, как динозавр был описан и назван, в Американском музее естественной истории уже собрали и выставили один из самых зрелищных образцов (рис. 16.1) — четвёртый по счёту из обнаруженных Брауном. Описывая первый экземпляр королевского тираннозавра, Осборн упоминал в своей статье, что «это животное — непревзойдённый образец эволюции крупных плотоядных динозавров; в общем, он удостаивается царственного и высокородного родового имени, которым я его нарекаю».
 Рис. 16.1.
Рис. 16.1. Классический вариант сборки тираннозавра в Американском музее естественной истории; так он выглядел примерно с 1910-го по 1990-е годы. Позу «кенгуру» выбрали потому, что королевский тираннозавр казался медлительным ящером, таскавшим за собой хвост (изображение № 327524, предоставлено библиотекой Американского музея естественной истории)
Вскоре Осборн приобрёл желаемую известность в прессе, опубликовав 3 декабря 1906 года в New York Times статью о только что анонсированных новых образцах динозавра. Там он описывал тираннозавра как «самое огромное агрессивное животное, о котором нам только известно», «величайшего из королей в царстве животных», «непревзойдённого сухопутного воителя» и «царственного людоеда из джунглей». В другой статье New York Times королевского тираннозавра назвали «гладиатором седой древности» и «последней Великой рептилией, Королём всех пресмыкающихся».
Реконструкции тираннозавра, вскоре нарисованные одним из первых палеохудожников Чарльзом Р. Найтом, сделали ти-рекса самым прославленным динозавром. С тех пор он стал настоящей «иконой», появлялся почти во всех медиа. Первые фильмы, где он упоминался, были ещё немыми: «Призрак сонной горы» (1918) и «Затерянный мир» (1925; по мотивам романа Артура Конан Дойля, написанного в 1912 году; того самого Конан Дойля, придумавшего Шерлока Холмса). Затем последовали «Кинг-Конг» (1933), тетралогия о Парке юрского периода, два последних ремейка «Кинг-Конга», а также мультфильм и мультсериал «Земля до начала времён», где тираннозавра назвали острозубом. На телеэкране тираннозавр стал звездой детского сериала «Барни и друзья», а огромных кукольных тираннозавров провозили на платформах во время парадов. Кроме того, стали хорошо продаваться разные товары с изображениями тираннозавра. Появилась даже британская рок-группа T. Rex. Образ гигантского хищника, возвышающегося над другими динозаврами (и над посетителями музеев), впечатляет (см. рис. 16.1). Покойный Стивен Джей Гулд говорил, что в возрасте пяти лет испугался, увидев скелет тираннозавра в Американском музее естественной истории, но именно это зрелище вдохновило его стать палеонтологом.
Естественно, в течение 110 лет с момента открытия и описания королевского тираннозавра наука не стояла на месте. Сильнее всего изменились представления о том, какова была осанка этого существа. Когда Осборн руководил установкой первых окаменелостей, кости собирались таким образом, словно тираннозавр был большим двуногим ящером, тащившим хвост по земле (см. рис. 16.1). Такое представление о тираннозавре до сих пор отражено в большинстве игрушек, в книгах и иной продукции. Но в 1970-е и 1980-е годы палеонтологи открыли следы крупных плотоядных динозавров и не обнаружили никаких борозд от хвоста. Это значит, что королевский тираннозавр (как и практически все динозавры) вытягивал хвост назад и стоял, опираясь на бёдра и задние ноги. Многочисленные биомеханические исследования продемонстрировали, что такая поза довольно устойчива. Благодаря исследованию, за которым внимательно следил писатель и сценарист Майкл Крайтон, фильм «Парк юрского периода» помог популяризовать следующее представление о королевском тираннозавре: это быстрый и умный хищник, стоявший на задних лапах, как на ходулях, и вытягивавший хвост назад. Теперь данный образ постепенно проникает в культурный и коммерческий контекст, связанный с этим динозавром (рис. 16.2).
 Рис. 16.2.
Рис. 16.2. Современный вариант сборки королевского тираннозавра в Американском музее естественной истории. Как и у других двуногих динозавров, его тело ориентировано по горизонтали, а хвост отставлен назад, служит для балансировки туловища над задними лапами. На заднем плане видна старая сборка горгозавра, скелет которого не удалось переориентировать в современную позу (фотография из фонда Wikimedia Commons)
Наши знания о королевском тираннозавре расширяются, причём за счёт исследований, которые совсем недавно выглядели за гранью возможного. Например, моделирование силы укуса королевского тираннозавра даёт от 35 тыс. до 57 тыс. ньютонов для задних зубов: в три раза сильнее укуса большой белой акулы, в три с половиной раза сильнее укуса австралийского гребнистого крокодила, в семь раз сильнее, чем у аллозавра, и в 15 раз — чем у африканского льва. В более новом исследовании силу укуса оценили в диапазоне от 183 тыс. до 235 тыс. ньютонов, что эквивалентно укусу крупнейших особей гигантской акулы мегалодона (см. главу 9).
Его исполинский череп имел длину 1,5 м, но был просто нашпигован всевозможными отверстиями, впадинами и воздухоносными мешками, благодаря чему становился легче. В горизонтальном сечении кончик морды тираннозавра имел форму буквы U, что обеспечивало ему бо́льшую силу укуса, чем у хищных динозавров-теропод, обладавших V-образной мордой. Однако морда тираннозавра была узкой по сравнению с широким затылком, поэтому его глаза направлены вперёд: он имел превосходное бинокулярное зрение, дававшее стереоскопическую картинку и позволявшее точно оценивать расстояние. Огромные зубы (длиной до 30 см от корня до верхушки) были загнуты назад и по форме напоминали мясницкие ножи длиной с банан, имели зазубренные кромки для шинкования мяса, а также специальные усиливающие рёбра сзади. Передние зубы в поперечном сечении имели форму буквы D, поэтому вряд ли могли обломиться, если тираннозавр делал резкий укус и рывок назад.
Что они ели? На многочисленных костях динозавров имеются шрамы, которые мог оставить лишь тираннозавр, а у некоторых особей есть затянувшиеся раны, нанесённые сородичами. Более того, в морде и на шее у многих из них попадаются обломившиеся тираннозавьи зубы. Очевидно, королевские тираннозавры пожирали многих других динозавров и дрались между собой. Споры относительно рациона сводятся к тому, были ли эти животные чистыми хищниками либо падальщиками. Как и во многих спорах, при такой формулировке два варианта представляются взаимоисключающими, но природа всегда сложнее упрощённых дискуссий. Большинство крупных хищных млекопитающих (львы, тигры, ягуары, пумы) одновременно являются падальщиками. Поймать дичь настолько сложно, что им не до разборчивости, и они едят дохлятину, если находят, а за живой добычей охотятся, лишь не имея выбора. Есть даже образцы полуобглоданных скелетов тираннозавров с отметинами от зубов сородичей, что указывает на каннибализм.
На шее королевского тираннозавра был S-образный изгиб, как у большинства теропод. Тираннозавр имел более короткую шею, чем большинство других теропод, однако она была очень крепкой и мощной, что позволяло чудовищу с огромной силой ею размахивать, сбивая добычу с ног или вырывая куски плоти. Всем известно, какие крошечные передние лапки были у тираннозавра, и на них всего по два рабочих пальца, а не по три, как часто ошибочно изображают. На самом деле у большинства хищных динозавров всего по три рабочих пальца на каждой передней лапе, но в популярных источниках многих из них изображают с пятипалыми передними конечностями. Существует множество версий о том, какова могла быть функция крошечных лапок. Однако короткие и почти нефункциональные передние конечности были типичны для большинства высокоразвитых теропод. Правда, передние лапки тираннозавра уж совсем маленькие. Более того, чем крупнее становились череп и челюсти теропод, тем сильнее уменьшались их передние лапки. Это значит, что хищные динозавры специализировались в направлении мощного укуса, используя в качестве главного оружия сильные челюсти и шею. Передние конечности становились рудиментами, которые не использовали для удерживания агонизирующей добычи.
В течение некоторого времени окаменелости тираннозавров попадались сравнительно редко, было известно лишь несколько частично сохранившихся скелетов. Но в последние 20 лет братия коллекционеров бросила огромные силы на их поиски (особенно после того, как экземпляр под названием «Сью» продали на аукционе более чем за 8 млн долларов). Сегодня по частичным скелетам известно более 50 особей. У нас в распоряжении есть тираннозавры любого возраста — от малюток до подростков, молодых и очень старых особей. Таким образом, известно, что тираннозавр стремительно рос примерно до 14-летнего возраста, к которому достигал веса 1800 кг. Затем он поправлялся примерно на 600 кг в год, пока не матерел и его рост не замедлялся. Томас Хольтц говорил, что тираннозавры «жили быстро и умирали молодыми», поскольку наряду со стремительным взрослением для них была характерна высокая смертность. Млекопитающие, напротив, взрослеют медленнее и живут дольше.
При таком изобилии особей учёные долго пытались определить, кому принадлежат те или иные окаменелости — самцам или самкам. Предлагали ориентироваться на множество различий, но большинство из них не выдержали критики. Признаки, которые обычно связывают с половым диморфизмом, оказались обусловлены в основном географическими и популяционными факторами. Однако пол одного экземпляра не вызывает сомнений. В кости B-рекса, найденного в штате Монтана, сравнительно хорошо сохранились мягкие ткани, в том числе костно-мозговая, типичная для овулирующих птичьих самок.
Птицы произошли от теропод — близких родственников тираннозавров (см. главу 18). Хотя большинство окаменелостей тираннозавров — просто кости, на некоторых остались отпечатки шкуры, которая, вероятно, была покрыта перьями. Затем в формации Исянь на территории Китая нашли маленького тираннозавра
Dilong paradoxus, который, судя по всему, был покрыт нитеобразными перьями или пухом. Открытие
Yutyrannus huali в Китае доказало, что перья (в основном нитеобразные или пушистые) покрывали этого динозавра практически полностью. Два этих экземпляра свидетельствуют, что тираннозавра можно было бы реконструировать оперённым, а не голым и кожистым, как его обычно изображают (даже в «Мире юрского периода», четвёртой серии саги «Парк юрского периода» динозавры по-прежнему голые).
Благодаря наличию многочисленных экземпляров и подробным исследованиям, проведённым палеонтологами, королевский тираннозавр является самым известным хищным ящером. А какие ещё большие хищники тогда существовали?
Из Африки
В конце XIX века Германия считалась лидером почти во всех сферах научного знания, особенно в эмбриологии, анатомии и эволюционной биологии. Ведущие немецкие учёные были так авторитетны, что первые американские палеонтологи, в частности Генри Фэрфилд Осборн и Уильям Бэрримен Скотт, получали последипломное образование именно в Германии.
Немецкие археологи значительно продвинулись в египтологии. Во главе этого движения стоял легендарный Карл Рихард Лепсиус. Генрих Шлиман открыл древнюю Трою (территория современной Турции), а также провёл первые исследования микенской Греции. Огромный Пергамский музей в Берлине может похвастаться одной из лучших коллекций произведений искусства и артефактов из древней Олимпии, с острова Самос, из Пергама, Милета, Приены и Магнесии. Там же находятся грандиозные произведения искусства и архитектуры из древней Вавилонии и Ассирии, а также легендарный бюст египетской царицы Нефертити.
В те времена немецкая палеонтология была поистине ультрасовременной. Многие ведущие учёные, работавшие в области палеонтологии и в смежных науках с конца XVIII до начала XX века, были немцами. Среди самых известных следует назвать одного из пионеров палеоботаники Эрнста Фридриха фон Шлотгейма (1764–1832), легендарного исследователя и биолога Александра фон Гумбольдта (1769–1859), одного из первых геологов Леопольда фон Буха (1774–1853), эмбриолога и зоолога, одного из главных сторонников Дарвина — Эрнста Геккеля (1834–1919), Карла Альфреда фон Циттеля (1839–1904), автора популярного учебника по палеозоологии, а также влиятельного палеонтолога Отто Г. Шиндевольфа (1896–1971).
Некоторые из них работали на легендарных залежах окаменелостей в Германии, например на Зольнхофельнских известняках и Хольцмаденских сланцах. Но многие вели исследования за границей, как археологи и другие коллеги-учёные. Особенно активные работы развернулись в тех частях Африки, где находились германские колонии. Так, в период с 1909 по 1911 год на территории Германской Восточной Африки (современная Танзания) велись грандиозные раскопки в отложениях Тендагуру, откуда Вернер Яненш извлёк удивительно полный скелет жираффатитана (ранее этот динозавр именовался брахиозавром). Сегодня он экспонируется в Музее естественной истории (Музее Гумбольдта) в Берлине (см. рис. 17.5). Там же был найден скелет стегозавра (
Kentrosaurus), спина которого усажена шипами, а не пластинами, а также многие другие необычные динозавры.
Ещё одним известным немецким палеонтологом, работавшим в Африке, был барон Эрнст Штромер фон Рейхенбах (1870–1952). В 1910–1911 годах он руководил знаменитой экспедицией в Египет, в то же самое время Яненш работал в Германской Восточной Африке. После двух относительно малоуспешных вылазок из Каира (в ходе одной из них Рихард Маркграф, коллега фон Рейхенбаха, открыл на территории современной Ливии окаменелости одного из древнейших приматов,
Libypithecus) фон Штромер и Маркграф добрались до далёких пустынь на западе Египта и до оазиса Бахария на самой границе с Ливией. 18 января 1911 года Штромер наконец нашёл кости огромных динозавров. По его словам, он обнаружил:
... три большие кости, которые я попытался выкопать и сфотографировать. Конечность, лежавшая выше, сильно разрушена и фрагментарна, но имеет 110 см в длину и 15 см в толщину. Вторая кость, лежавшая ниже и сохранившаяся лучше, — вероятно, бедренная, имеет целых 97 см в длину, а в центральной части достигает 15 см в толщину. Третья находится слишком глубоко в земле, на её извлечение потребуется гораздо больше времени.
В течение нескольких следующих недель он нашёл в Бахарии и другие экземпляры, но к февралю 1911-го был вынужден вернуться в Германию. Несколько десятилетий Штромер описывал фрагменты костей удивительных динозавров и публиковал о них статьи. В частности, он рассказал о зауроподе египтозавре, гигантском крокодиле стоматозухе, о фрагментах некоторых теропод: бахариазавра, а также суперхищников кархародонтозавра и спинозавра.
Все окаменелости с оазиса Бахария хранились в Мюнхенском палеонтологическом музее, где сейчас располагается Баварская государственная зоологическая коллекция. К началу 1944 года союзники уже бомбили все крупные немецкие города, особенно те, где имелись важные военные объекты (шла подготовка к высадке в Нормандии, состоявшейся 6 июня 1944 года). Берлинский естественнонаучный музей с его бесценными экспонатами, в числе которых жираффатитан, «берлинский экземпляр» археоптерикса и множество удивительных ихтиозавров из Хольцмадена, едва не пострадал: во время одной из бомбёжек был полностью разрушен железнодорожный вокзал, находившийся по соседству. В ночь с 24 на 25 апреля 1944 года Королевские ВВС Великобритании провели мощную бомбардировку Мюнхена, в результате которой были полностью уничтожены все находки Штромера, а также бесценные исторические и прочие коллекции Мюнхенского палеонтологического музея. За одну ночь оказалась похоронена и перечёркнута многовековая работа биологов и палеонтологов, исчезли накопленные ими коллекции, причём не сохранилось никаких сведений о том, как выглядело большинство утраченных экземпляров, что они собой представляли. Из наследия Штромера остались лишь его собственные научные иллюстрации, сделанные в 1915 году, фотографии выставок, а также
несколько окаменелостей, найденных в последние годы.
Самой именитой окаменелостью, найденной в ходе экспедиции Штромера, был спинозавр. Благодаря фильму «Парк юрского периода 3» он известен всем фанатам динозавров. Его изображают как гигантского двуногого хищника с крокодильей мордой и «парусом» на спине (см. рис. 16.3 и 16.4) — настолько крупного, что он мог побороть и сожрать более мелкого тираннозавра. Штромер изобразил лишь несколько огромных стержней, поддерживавших этот спинной «парус», и нарисовал нижнюю челюсть, отдельные зубы, несколько рёбер и позвонков. Спинные стержни были очень длинными, до 1,65 м. Штромер описывал (но никогда не рисовал) элементы верхней челюсти, которые оказались навсегда утрачены. Экземпляры были действительно огромны (нижняя челюсть достигала 75 см в длину), но настолько фрагментарны, что истинный внешний вид спинозавра остаётся загадкой. В начале 1990-х годов экспедиция из Пенсильванского университета под руководством Питера Додсона, Мэтью Ламанны, Джошуа Смита и Кеннета Лаковары вернулась в оазис Бахария. Они нашли несколько новых экземпляров, в том числе зауропода
Paralititan stromeri, названного в честь Эрнста Штромера, но не добыли нового материала по спинозавру.

 Рис. 16.3.
Рис. 16.3. Спинозавр: (
А) известные кости (выделены тёмным цветом) на основании последних работ, выполненных Низаром Ибрахимом, Полом Серено и другими; (
Б) реконструкция внешнего вида ((
А) фотография предоставлена Paul Sereno и Michael Hettwer; иллюстрация (
Б) предоставлена Nobumichi Tomura)
После 1944 года несколько окаменелостей спинозавра открыли в Марокко и Тунисе. Недавно группа исследователей, в составе которой были Пол Серено и его студент-постдок Низар Ибрахим, нашли новые окаменелости спинозавра и широко анонсировали, что работают над новой реконструкцией. Они предполагают, что спинозавр имел относительно длинное поджарое тело и очень короткие передние и задние лапы — то есть совсем не напоминал увеличенную копию королевского тираннозавра, каким его изобразили в «Парке юрского периода 3» (см. рис. 16.3,
Б). Длинный узкий клюв животного был приспособлен не для поедания других динозавров, а для ловли рыбы. Кроме того, в средней части клюва имелись ноздри, а также каналы для нервов и кровеносных сосудов, помогавших следить за изменениями в давлении воды. Все эти факты подсказывают, что по образу жизни спинозавр напоминал крокодила, а не большинство двуногих динозавров-теропод.
Эти физические доказательства согласуются с результатами химического анализа костей, свидетельствующими, что в рационе спинозавра преобладала рыба и иные водные существа. Плотность костей конечностей также подсказывает, что большую часть времени спинозавр проводил в воде. Такие же кости у других водных животных, например у гиппопотама, очень плотные и служат балластом. Идут серьёзные споры о том, мог ли спинозавр с такими короткими и толстыми лапами выползать на сушу, однако он определённо не являлся быстрым сухопутным бегуном и хищником, который мог бы преследовать и убивать сравнительно крупных динозавров. Пальцы на его передних лапах длинные и изящные, приспособлены для ловли мелкой добычи, а нижняя часть ступни уплощена, что позволяло становиться на всю лапу (стопоходящий), а не на кончики пальцев (пальцеходящие), как большинство динозавров. Строение тонких и длинных пальцев на передних и задних лапах позволяет предположить, что между пальцами у спинозавра даже могли быть перепонки.
Наконец, у него на спине имеется огромный «парус», сформировавшийся между длинными выростами невральной дуги поверх позвоночника; поэтому спинозавр и получил такое название. У каждого палеонтолога есть своя подборка любимых теорий, объясняющих эту анатомическую особенность, но большинство сходятся на том, что это был не настоящий парус, помогающий быстрее плавать. Причём он настолько велик и заметен, что спинозавру было явно нелегко затаиться под поверхностью воды, как это делают крокодилы. Некоторые полагают, что парус одновременно служил солнечной батареей и радиатором, однако редкие динозавры нуждались в таком органе. Самое распространённое мнение таково: парус подобен рогам оленей и антилоп и в основном служил для узнавания сородичей и самоутверждения среди самцов.
Громкие заявления о свеженайденных окаменелостях спинозавра вызвали определённый скепсис среди палеонтологов, поскольку при реконструкции некоторых костей (особенно тазовых) могли быть допущены ошибки. Кроме того, реконструкция была сделана на основе фрагментов скелета, принадлежавших разным особям. Некоторые кости восстановили в виде цифровых копий с сохранившихся фотоснимков Штромера и изготовили на 3D-принтере.
В частности, следует со здоровым скептицизмом относиться к оценке размеров спинозавра. Если ориентироваться на его «облегчённый» профиль и короткие конечности, он и близко не мог быть таким массивным и крупным, как гигантские сухопутные хищники. Ибрахим, Серено и их коллеги утверждают, что ящер имел 15,2 м в длину, но если присмотреться к схеме найденных костей, выясняется, что среди них очень мало хвостовых (см. рис. 16.3,
А). Поэтому реконструкция хвоста (а следовательно, и общей длины динозавра) в лучшем случае спекулятивна. Предлагались разные оценки его размера, но при наличии столь скудного материала все они выглядят зыбкими. В 1926 году немецкий палеонтолог Фридрих фон Хюне, изучавший первые окаменелости спинозавра, предположил, что животное могло достигать 15 м в длину и весить 6 т. В 1988 году Грегори Пол тоже дал оценку длины в 15 м, но уменьшил массу до 4 т. Однако в 2007 году Франсуа Терьен и Дональд Хендерсон применили новые техники масштабирования и оценили длину в 12,6–14,3 м, вес — от 12 до 21 т. Таким образом, у них получился менее крупный, но более тяжёлый экземпляр, чем считалось ранее. Если крупнейший королевский тираннозавр имел длину 13 м и весил около 10 т, то спинозавр от него почти не отличался по размеру. Вероятно, он был не так велик, чтобы отбросить тираннозавра словно игрушку, как это показано в «Парке юрского периода 3». Поскольку все экземпляры слишком фрагментарны, в данном случае мы не можем ничего утверждать.
Если спинозавра нельзя считать крупнейшим хищным ящером с полной уверенностью, что насчёт другого большого теропода, найденного в Африке, кархародонтозавра? Его в 1924 году открыли французские палеонтологи Шарль Депере и Ж. Саворнин, которые извлекли несколько огромных зубов из нижнемеловых континентальных отложений в Алжире. Зубы были примерно такими, как у первого из обнаруженных динозавров, мегалозавра, поэтому учёные назвали свою находку
Megalosaurus saharicus. В 1914 году Штромер нашёл частично сохранившийся череп этого животного в оазисе Бахария, а ещё добыл множество зубов, когтей, разнообразных тазовых костей и костей конечностей. Когда в 1931 году дошли руки до описания этого материала, он переименовал динозавра в
Carcharodontosaurus saharicus, поскольку окаменелости совсем не походили на останки мегалозавра, найденного в Англии. Как следует из названия, огромные зубы по форме и размеру во многом были такими же, как у большой белой акулы (
Carcharodon carcharias). К сожалению, штромеровские окаменелости кархародонтозавра были уничтожены во время бомбардировки Мюнхена в 1944 году, как и материал по спинозавру и вся остальная коллекция.
Частично сохранившийся череп казался впечатляющим, но был неполным, а о скелете было известно совсем мало. Поэтому точная реконструкция и оценка размеров не представлялась возможной. Согласно первым расчётам, из всех хищных динозавров кархародонтозавр имел самый крупный череп, но основные элементы отсутствовали. Когда их нашли, длину черепа пересмотрели, уменьшив с почти 2 до 1,6 м. В соответствии с последними измерениями длина кархародонтозавра могла составлять 12–13 м, а вес — от 6 до 15 т. Таким образом, по размеру он сравним со спинозавром и крупным королевским тираннозавром. Но экземпляр настолько фрагментарен, что мы не можем с уверенностью сказать, какой из этих трёх динозавров был самым крупным (см. рис. 16.4).
 Рис. 16.4.
Рис. 16.4. Сравнение размеров основных теропод: огромные размеры кархародонтозавра сомнительны, поскольку его окаменелости слишком фрагментарны для такой реконструкции. Спинозавр изображён в соответствии с «длинноногим» вариантом реконструкции и не соответствует новым данным о коротконогих и приземистых экземплярах (рисунок Mary P. Williams)
Затем, в 1995 году, Пол Серено предпринял несколько экспедиций в Сахару. В формации Кем Кем на территории Марокко, близ алжирского региона, где Депере и Саворнин нашли первые зубы, Серено и его группа из Чикагского университета добыли более полный материал по черепу карходонтозавра (см. рис. 16.5). Финансирование для этой работы Серено получил от Национального географического общества, и его открытия — эта и другие замечательные окаменелости — сразу попали во все выпуски новостей. Его опасная экспедиция через смертоносную Сахару с её адской жарой, убийственными песчаными бурями, плохими дорогами и шайками бандитов да террористов стала сюжетом не одного документального фильма с тех пор, как он впервые описал её в 1996 году. В 2001 году бывший студент Серено, Ханс Ларссон, выполнил подробный анализ некоторых участков области уха и черепной коробки, которые были не известны по другим черепам. В 2007 году Серено и его студент Стивен Брузатте опубликовали описание ещё одного вида,
Carcharodontosaurus iguidensis, найденного в формации Эчкар на территории Нигера. Это животное имело примерно такие же размеры, как марокканские экземпляры. Поскольку первые окаменелости, найденные Штромером, не сохранились, Серено и Брузатте решили считать новый череп из Марокко заменой для типового экземпляра, или неотипом.
 Рис. 16.5.
Рис. 16.5. Череп кархародонтозавра, найденный в формации Кем Кем. Для сравнения показан человеческий череп. (Фотография предоставлена Paul Sereno и Michael Wettner)
Хотя, судя по большинству известных фрагментов скелета, кархародонтозавр имел телосложение обычного теропода, а его череп был своеобразен (см. рис. 16.5). Свод черепа сильно изогнут вверх, в боковой части есть очень широкие отверстия, примерно как у королевского тираннозавра и других крупных теропод. Поэтому, несмотря на размеры, огромный череп весил сравнительно мало. В пропорциях к телу мозг кархародонтозавра был примерно таким же, как у более мелкого аллозавра, его близкого родича. Кроме того, у него имелся большой зрительный нерв и широкие глазницы, то есть этот хищник полагался преимущественно на зрение.
Если экземпляры спинозавра и кархародонтозавра столь фрагментарны, что мы не можем с уверенностью судить, были ли они значительно крупнее королевского тираннозавра, кто же был крупнейшим из когда-либо живших хищников? Оказывается, этот гигант обитал не в Африке, а на другом континенте, отколовшемся от Гондваны, — в Южной Америке. Это был близкий родич кархародонтозавра, живший в раннем меловом периоде.
Крупнее королевского тираннозавра
В отличие от спинозавра и кархародонтозавра, динозавр, который, вероятно, был крупнейшим из всех теропод, известен по довольно полному скелету. Палеонтолог-любитель Рубен Дарио Каролини открыл его окаменелости в 1993 году в нижнемеловых отложениях на юге Аргентины. Родольфо Кориа и Леонардо Сальгадо в 1995 году назвали этого динозавра
Giganotosaurus carolinii: родовое название происходит от греческого выражения «большая южная ящерица», а видовое имя дано в честь Каролини. Многие неспециалисты произносят это название с ошибкой: ГИГАНТОзавр, а правильно: ГИГА-НОТО-завр.
В отличие от экземпляров крупных африканских теропод, скелет гиганотозавра почти полный (он сохранился примерно на 70%) и содержит большую часть черепа; нижнюю челюсть, таз и задние конечности, а также большую часть позвоночника. В нём не хватает передних конечностей и некоторых других фрагментов. Соответственно, оценка длины сделана по относительно полным черепам и скелетам, а также по реальным конечностям (не наугад).
Самый большой череп и челюсти в 1988 году в Аргентине нашёл Хорхе Кальво). Оказалось, что их длина — около 1,95 м; это больше, чем у каких-либо других теропод. Череп у гиганотозавра, как и у его близкого родича кархародонтозавра, довольно изящен и легко сложён: с высоким сводом и множеством отверстий, окружённых костными распорками. На верхней части морды и над глазами есть более огрублённые области. Затылок скошен вперёд под углом, поэтому челюстные суставы висят сзади и снизу от того места, где череп прикрепляется к шейным позвонкам.
 Рис. 16.6.
Рис. 16.6. Наиболее полный скелет гиганотозавра, выставленный в Муниципальном музее Эрнесто Бахманна в Вилья Эль Чокон (Аргентина) (фотография предоставлена R. Coria)
Учитывая облегчённую конструкцию черепа, маловероятно, что гиганотозавр мог сильно кусаться; по-видимому, мощность его укуса была в три раза ниже, чем у королевского тираннозавра. Расположенные на нижней челюсти широкие режущие зубы, напоминающие акульи, больше подходили для нанесения ударных ран, чем для укусов «книзу» в бульдожьем стиле; такие же зубы были у королевского тираннозавра. Следовательно, гиганотозавр мог нападать на более мелкую добычу, например на сравнительно небольшого андезавра (зауропода-титанозавра), диплодоков
Nopcsaspondylus и
Limaysaurus, а также на прочих игуанодонов и мелких хищных динозавров, известных под общим названием «дромейозавры», например, на велоцираптора и ещё более мелких животных. Они встречаются в тех же нижнемеловых аргентинских отложениях, что и гиганотозавр.
Исходя из размеров конечностей, почти полного позвоночника, общего вида черепа и скелета, крупнейшие особи гиганотозавра достигали около 14,2 м в длину и весили от 6,5 до 13,8 т. Они были существенно больше самых крупных королевских тираннозавров, имевших 13 м в длину и весивших 8 т. Итак, гиганотозавр — лучший претендент на звание величайшего хищника всех времён.
Лучше один раз увидеть!
Скелеты королевского тираннозавра выставлены во многих музеях мира, но самые известные находятся в Американском музее естественной истории (Нью-Йорк; это первый собранный скелет тираннозавра), Музее естественной истории им. Карнеги (Питтсбург; оригинальный типовой экземпляр Генри Фэрфилда Осборна), Денверском музее природы и науки (тираннозавр реконструирован в «танцующей» позе и буквально нависает над холлом), Музее естественной истории под открытым небом (Чикаго; выставлен «Сью», тот самый неоднозначный экземпляр, который ушёл с аукциона за 8 млн долларов), Музее Скалистых гор в Бозмене (штат Монтана; куратор этого музея, Джек Хорнер (Jack Horner), нашёл больше экземпляров тираннозавра, чем кто бы то ни было), Национальном музее естественной истории Смитсоновского института (Вашингтон, округ Колумбия), Музее естественной истории округа Лос-Анджелес (город Лос-Анджелес; три экземпляра тираннозавра, от малютки до почти взрослого) и в Палеонтологическом музее Калифорнийского университета (город Беркли).
Новая реконструкция спинозавра выставлена в штаб-квартире Национального географического общества (Вашингтон, округ Колумбия).
В Аргентине полные скелеты гиганотозавра экспонируются в Муниципальном музее Кармен Фунес (Пласа Уинкуль) и Муниципальном музее Эрнесто Бахманна (Вилья Эль Чокон). В США слепки с таких скелетов выставлены в Академии естественных наук Дрексельского университета (Филадельфия) и Музее естественной истории Фернбанк (Атланта).
Дополнительная литература
Brett-Surman, M. K., Thomas R. Holtz Jr., and James O. Farlow, eds. The Complete Dinosaur. 2nd ed. — Bloomington: Indiana University Press, 2012.
Carpenter, Kenneth. The Carnivorous Dinosaurs. — Bloomington: Indiana University Press, 2005.
Fastovsky, David E., and David B. Weishampel. Dinosaurs: A Concise Natural History. 2nd ed. — Cambridge: Cambridge University Press, 2012.
Holtz, Thomas R., Jr. Dinosaurs: The Most Complete Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. — New York: Random House, 2007.
Nordruft, William, with Josh Smith. The Lost Dinosaurs of Egypt. — New York: Random House, 2007.
Parrish, J. Michael, Ralph E. Molnar, Philip J. Currie, and Eva B. Koppelhus, eds. Tyrannosaurid Paleobiology. — Bloomington: Indiana University Press, 2013.
Paul, Gregory S. The Princeton Field Guide to Dinosaurs. — Princeton, N. J.: Princeton University Press, 2010.
17. Страна гигантов
Крупнейшее сухопутное животное
Аргентинозавр
В то время были на земле исполины.
Бытие, 6:4
В начале XIX века невероятный мир вымерших животных был почти не известен большинству людей. Кое-где могли найти несколько больших костей, но считалось, что они принадлежали библейским «исполинам», либо их просто отметали, не удостоив научного исследования. К 1810 году французский барон Жорж Кювье подробно описал ископаемых мамонтов и мастодонтов, незадолго до того открытых в ледниковых отложениях Европы и Северной Америки, и пришёл к выводу, что это вымершие животные, обитавшие в тёмном и ветреном «допотопном мире», реликты предыдущего акта творения, не упомянутого в Библии. Затем Мэри Эннинг стала находить окаменелости удивительных морских рептилий в юрских отложениях на территории Англии, и вскоре «допотопный мир» гигантских и страшных ихтиозавров и плезиозавров стал подпитывать фантазию художников, пытавшихся изобразить доисторическое прошлое (см. рис. 14.4). Однако ещё никто не представлял себе мир, в котором доминировали динозавры.
Лишь после многочисленных разрозненных находок динозавровых костей пришло понимание, что это останки гигантских вымерших рептилий, а не какие-нибудь «драконьи кости» (как их долго называли китайцы) либо кости исполинских людей, упомянутых в Библии. В 1676 году большую кость нашли в известняках Тейнтон в каменоломне Стоунсфилд близ Оксфорда (среднеюрские отложения). Годом позже Роберт Плот, профессор химии из Оксфордского университета, опубликовал книгу «The Natural History of Oxfordshire» («Естественная история Оксфордшира») и сам её проиллюстрировал. Именно тогда появилось первое изображение кости динозавра в научной литературе (рис. 17.1). Плот верно определил, что перед ним — нижний конец бедренной кости, и решил, что она могла принадлежать римскому боевому слону или библейскому гиганту.
 Рис. 17.1.
Рис. 17.1. Оригинальный рисунок Роберта Плота и первая в мире иллюстрация, связанная с динозавром, позднее названная Scrotum humanum (на самом деле это кончик бедренной кости динозавра-теропода, вероятно мегалозавра) (рисунок из фонда Wikimedia Commons)
В 1763 году Ричард Брукс написал книгу, в которой повторно опубликовал иллюстрацию Плота и назвал её
Scrotum humanum («Мошонка человека»), поскольку она отдалённо напоминает пару гигантских окаменевших мужских яичек. В 1970 году шли споры о том, считать ли злосчастное
Scrotum humanum валидным названием первого известного динозавра и не заменить ли им позднейший вариант
Megalosaurus. Международная комиссия по зоологической номенклатуре постановила, что образец слишком неопределённый, чтобы однозначно указать динозавра, которому он мог принадлежать. Кроме того, такое название позиционировалось не в качестве научного, а как подпись из двух слов.
Между 1815 и 1824 годами известный естествоиспытатель, преподобный Уильям Бакленд описал фрагменты челюсти и другие кости огромного хищного «ящера», обнаруженные недалеко от его дома в Оксфорде, и назвал это животное мегалозавром (в переводе с греческого «большая ящерица»). Затем в 1825 году доктор Гидеон Мантелл описал зубы и некоторые другие костные фрагменты огромной рептилии, найденные в нижнемеловых вельдских отложениях в Тилгейт-Форест (графство Сассекс), и назвал это животное игуанодоном (игуанозубым).
К 1842 году, после этих и других открытий, британский натуралист Ричард Оуэн придумал слово «динозавры» (в переводе с греческого «ужасные ящеры») для обозначения всех экземпляров. Уже на следующий год Оуэн описал несколько зубов и других огромных костей другого существа, найденного в 1825 году близ Чиппинг-Нортона; его он назвал цетиозавром (китоящером). Поскольку материал был скуден, Оуэн предположил, что эти окаменелости принадлежали гигантской морской рептилии, напоминавшей крокодила, но Мантелл поправил его и указал, что они, вероятно, относятся к гигантскому сухопутному пресмыкающемуся, похожему на игуанодона или мегалозавра. Оуэн не согласился и не включил цетиозавра в число динозавров, когда в 1842 году предложил этот термин. Останков цетиозавра было недостаточно, чтобы в то время точно реконструировать это существо.
Однако в марте 1868 года близ Блетчингдона несколько рабочих нашли гигантскую бедренную кость зауропода, а вскоре были открыты другие крупные фрагменты конечностей и позвонки. Эти кости явно свидетельствовали, что цетиозавр не походил на большого крокодила, а был гигантской рептилией, шагавшей на четырёх ногах-колоннах (рис. 17.2). Скелет был недостаточно полон, чтобы распознать длинную шею и длинный же хвост, которые сегодня считаются характерными чертами зауропод, но был и остаётся одним из самых полных скелетов зауропод, найденных в Европе.
 Рис. 17.2.
Рис. 17.2. Кости конечностей из частично сохранившегося скелета цетиозавра, выставленные в Музее естественной истории при Оксфордском университете (фотография предоставлена M. Wedel)
Только в 1870–1880-е годы группы исследователей, работавшие вместе с Отниелом Чарльзом Маршем из Йельского университета и их коллегами, трудившимися с Эдвардом Дринкером Копом из Филадельфии, нашли почти полные скелеты зауропод в Колорадо и Вайоминге. Так, Марш добыл несколько удивительно полных экземпляров в местечке Комо Блафф в центральной части Вайоминга. Вскоре все полученные им экземпляры получили имена: апатозавр и атлантозавр — в 1877 году, морозавр и диплодок — в 1878 году, бронтозавр и барозавр — в 1890 году. А Коп в 1877 году назвал камаразавра и каулодона. Сегодня учёные считают, атлантозавр и бронтозавр — то же животное, что и апатозавр; морозавр и камаразавр тоже относятся к одному виду. Лишь апатозавр, диплодок, камаразавр и барозавр по-прежнему считаются валидными родами.
Окаменелостей накопилось так много, что к 1878 году Марш смог объединить их, включая цетиозавра, в группу, которую назвал зауроподами (в переводе с греческого «ящероногие»). К сожалению, по каждому из этих динозавров Марш опубликовал лишь небольшие статьи без иллюстраций, поэтому все гиганты оставались практически неизвестными до 1900 года.
Последний этап открытия «китовых рептилий» наступил, когда сотрудники музеев начали осознавать, что огромные скелеты, пылящиеся в запасниках, способны прославить их учреждения и собрать толпы посетителей. К 1905 году в Американском музее естественной истории в Нью-Йорке, Музее естественной истории им. Карнеги в Питтсбурге, Музее естественной истории им. Пибоди при Йельском университете в Нью-Хейвене (штат Коннектикут) собрали скелеты крупных зауропод, назвав их бронтозаврами — это невалидное имя придумал Марш.
К сожалению, слово «бронтозавр», укоренившееся в нашей культуре, — младший синоним названия «апатозавр», и его нельзя использовать. Марш назвал бронтозавра в 1890 году, ориентируясь на частично сохранившийся скелет взрослой особи крупного зауропода, найденного в Комо Блафф. В те времена было принято, что любая окаменелость, даже слегка отличающаяся от уже известных экземпляров, получала новое название. Затем это название закрепилось за скелетами, собранными в Американском музее и Музее им. Пибоди, где стало культовым — и бронтозавры попали на страницы всех книг о динозаврах.
Позже выяснилось, что Марш назвал апатозавром чуть хуже сохранившуюся и более молодую особь того же динозавра. В 1903 году Элмер Риггс более тщательно изучил образцы Марша и пришёл к выводу, что апатозавр и бронтозавр — одно и то же животное. Но по правилам Международной комиссии по зоологической номенклатуре верным считается название, которое дано первым. Поэтому среди палеонтологов принято считать, что наименование «бронтозавр» с 1903 года является младшим синонимом. Наиболее влиятельный палеонтолог своего времени, Генри Фэрфилд Осборн из Американского музея естественной истории, отказался признавать анализ Риггса и способствовал тому, чтобы название «бронтозавр» надолго задержалось в обывательском сознании после того, как от него отказались учёные.
К сожалению, популярная литература и прочие источники информации часто не успевают за научным прогрессом. Поэтому название «бронтозавр» оставалось распространённым и в 1980-е, и в 1990-е годы, когда в музеях начали пересобирать имевшиеся скелеты, устанавливая их в более реалистичных позах. Поскольку у экземпляра из Комо Блафф не было черепа, к скелетам в Американском музее и Музее им. Пибоди добавили черепа короткомордого брахиозавра. Джон Остром и Джек Макинтош показали, что у апатозавра был вытянутый череп, примерно как у диплодока. Наконец, жалобы палеонтологов стали настойчивыми, и авторы детских книг и новостей постепенно начали учитывать новые научные данные — через 90 лет после первых публикаций об апатозавре.
Осборн также поручил легендарному художнику Чарльзу Найту изобразить классическую реконструкцию «бронтозавра». Вскоре широкая общественность буквально запала на бронтозавров и прочих длинношеих и длиннохвостых динозавров-зауропод (см. рис. 17.3). Они появились в самых первых фильмах с покадровой анимацией, в том числе в «Затерянном мире» (1925), где движущихся динозавров изобразил легендарный Уиллис О’Брайен, опиравшийся на иллюстрации Найта. Вскоре зауроподы были повсюду: на политических карикатурах, в кино, на товарах и даже на логотипе компании Sinclair Oil.
 Рис. 17.3.
Рис. 17.3. Культовое изображение бронтозавра, сделанное Чарльзом Найтом в 1905 году. Бронтозавр показан как неповоротливый обитатель болот, таскавший за собой хвост. Сегодня такая идея полностью устарела (изображение № 327524, предоставлено библиотекой Американского музея естественной истории)
Казалось трудно себе представить, что 110 годами ранее никто, кроме ряда учёных, понятия не имел об этих существах.
Образ жизни древних гигантов
За прошлый век наука о зауроподах проделала большой путь. Выявлено минимум 90 родов, а может, и больше. Хотя вопросы о том, какие названия зауропод являются валидными таксонами, представляют некоторую проблему. Поскольку эти животные были огромными, многие их кости прочные, долговечные и хорошо сохраняются, независимо от того, как сильно был изломан или размыт скелет. Поэтому большинство поименованных зауропод известны лишь по считанным костям: обычно речь идёт о фрагментах хребта или позвонках, иногда — о конечностях. Есть много частично сохранившихся скелетов, но и они, к сожалению, часто успевали остаться без головы, прежде чем фоссилизировались (обычно черепа более лёгкие и хрупкие, чем другие кости). Лишь немногие зауроподы известны по достаточно полным скелетам, и именно они встречаются в музеях: это апатозавр, диплодок, брахиозавр, камаразавр, барозавр, маменчизавр и некоторые другие.
Зауроподы произошли от группы триасовых динозавров, которые назывались прозауроподами. Прозауроподы были классическими переходными звеньями, связывавшими гигантских чудовищ юрского периода с более ранними эволюционными ветвями динозавров (среди них попадались существа размером с цыплёнка). Такие прозауроподы, как платеозавр, имели до 10 м в длину и весили до 4 т, но по размерам не могли сравниться со своими потомками (см. рис. 17.4). Тем не менее у них начали удлиняться шея и хвост. Хотя платеозавр перемещался почти исключительно на двух ногах, конечности некоторых прозауропод (например, меланорозавра) позволяли им ходить на четырёх либо на двух конечностях. Они обладали хорошо развитыми хватательными пальцами, которых не было у их потомков со «слоновьими» ногами.
 Рис. 17.4.
Рис. 17.4. Скелет прозауропода платеозавра, найденный в триасовых отложениях в Германии (фотография автора)
К середине и концу юрского периода наступил настоящий век гигантов. Эти чудовища были не медлительными болотными ящерами, таскавшими за собой хвосты, какими их представляли в начале XX века. Первые исследователи были поражены их размерами, и им не верилось, что зауроподы могли выдерживать собственный вес на суше, поэтому «селили» их в болотах. На самом деле ряд важных экземпляров (рядом с некоторыми сохранились и следы), а также многочисленные качественные биомеханические исследования радикально преобразили наши представления о зауроподах и их палеобиологии. Прежде всего, следы свидетельствуют, что зауроподы ходили, вытянув хвост и отставив его назад, поскольку борозд от хвоста почти не сохранилось. Вдобавок анализ формации Моррисон и других горных пластов, изобилующих зауроподами, показывает, что они не жили на болотах, а хорошо адаптировались не только к прибрежным регионам, но и к гораздо более засушливым экосистемам. Зауроподы были отличными ходоками и отправлялись на большие расстояния в поисках пищи. Они кормились древесной листвой, которую без труда доставали с помощью длинной шеи. Наконец, в теле зауропода было так много воздушных мешков, что они просто не могли как следует погрузиться в воду, не говоря о том, чтобы нырять.
Их анатомия довольно примечательна. Голова зауропод была совсем миниатюрной для таких огромных животных, зубы у большинства простые, напоминали карандаши или лопатки. Многие учёные ломали голову над тем, как динозавр мог прокормить такую огромную тушу, имея столь примитивный жевательный аппарат (напротив, утконосые и рогатые динозавры развили целые зубные батареи из сотен зубов-жерновов, способных перерабатывать массу растительности). Некоторые палеонтологи полагали, что зауроподы питались без разбора практически всем, что могли найти: с равным успехом жевали листву деревьев и паслись на плотном ковре папоротников. Учтите, что никаких цветковых растений, в частности травы, не существовало вплоть до раннего мелового периода — к тому времени дни расцвета гигантских юрских зауропод остались далеко позади.
Каждый отдельный позвонок во всей шее, спине и хвосте — инженерное чудо; благодаря множеству костных распорок и связок они получаются лёгкими и при этом очень прочными, хорошо удерживаются вместе с помощью множества прочных сухожилий. Кости зауропод, как у многих других динозавров и птиц (особенно идущие вдоль хребта), полны воздушных мешков и потому относительно лёгкие. Некоторые новейшие исследования показывают, что зауроподы обычно не задирали голову высоко и надолго (хотя большинство реконструкций именно таковы), поскольку в таком случае для кровоснабжения головы требовалось бы очень высокое кровяное давление. Опыты для проверки этой идеи ставили на домашних животных, которых разводили с целью получить потомство с болезненно высоким кровяным давлением. Ещё одно новое исследование (пока не опубликованное) предполагает, что зауроподы были способны управлять своим кровяным давлением и могли снабжать кровью высоко поднятую голову, обходясь без огромного сердца. Подобно жирафам, они могли иметь в шейных кровеносных сосудах специальные клапаны, препятствовавшие резкому снижению кровяного давления и не дававшие динозавру упасть в обморок, когда он высоко поднимал голову.
Будучи крупнейшими сухопутными животными всех времён, зауроподы имели массивные конечности и пальцы, превратившиеся в короткие диски или костные столбики, как у слонов. Однако, в отличие от слонов, зауроподы ходили на кончиках пальцев (были пальцеходящими, как большинство динозавров), а не опирались при ходьбе на ступню (так делают стопоходящие, к которым относятся люди и слоны). Правда, ступни зауропод были отчасти пригодны как для пальцехождения, так и для стопохождения. Огромные кости зауроподовых ног, разброс их следов, а также колоссальная масса, которую им приходилось нести, исключают версию о том, что эти животные быстро двигались. Большую часть времени они бродили ровным медленным шагом, но, вероятно, могли немного пробежать (как современные слоны). Однако с такими огромными ногами зауроподы могли и шагом покрывать огромные расстояния.
Существует несколько основных ветвей зауропод, в том числе обладавшие очень длинной шеей и хлыстовидным хвостом диплодоковые (например, диплодок и апатозавр); высокие брахиозавры с удлинёнными передними ногами и шеями, как у жирафа; приземистые титанозавры с миниатюрными головами (особенно много в Африке и Южной Америке, но они жили и на остальных континентах, включая Антарктику) и другие. Расцвет большинства зауропод пришёлся на конец юрского периода, но некоторые группы, например титанозавры, были многочисленны в Южном полушарии на протяжении мелового периода (в Северной Америке к тому времени почти исчезли), вполне могли дожить до его конца.
Размер имеет значение!
Естественно, размер имеет значение для животных, которые были крупнейшими обитателями своей экосистемы и самыми огромными обитателями суши, когда-либо существовавшими на свете. Многие из них претендовали на звание «самого большого динозавра», но через несколько лет лишались его после открытия нового гиганта. Ситуация осложняется тем, что чем крупнее динозавр, тем меньше его костей сохраняется. Самый крупный и массивный динозавр, почти полный скелет которого нам известен, — знаменитый брахиозавр (сегодня именуемый жираффатитаном). Скелет находится в Музее естественной истории (Музее Гумбольдта) в Берлине (см. рис. 17.5) и был найден в отложениях Тендагуру на территории Германской Восточной Африки (современная Танзания) в период с 1909 по 1912 год. Этот грандиозный экземпляр составлен из элементов пяти частично сохранившихся скелетов (в основном принадлежавших молодым особям) и возвышается на несколько этажей (13,5 м) над галереей, а в длину достигает 22,5 м. Его масса должна была составлять от 30 до 40 т.
 Рис. 17.5.
Рис. 17.5. Самый полный собранный скелет крупного зауропода Giraffatitan brancai (он же — брахиозавр), найденный в отложениях Тендагуру в Африке и выставленный в Музее естественной истории (Музее Гумбольдта) в Берлине (фотография M. Wedel)
Попадались и более крупные кости динозавров — например, большая берцовая кость из той же коллекции на 13% больше, чем у собранного жираффатитана, — но попытка точно определить размеры животного, ориентируясь лишь на несколько позвонков или элементов конечностей, чревата проблемами (рис. 17.6). Так, Мэтью Уэдел и Ричард Сифелли обнаружили четыре шейных позвонка огромного титанозавра, которого назвали завропосейдоном (в греческой мифологии Посейдон — бог моря и землетрясений). Эти окаменелости были найдены в раннемеловых пластах на территории Оклахомы; кости так огромны, что сначала их ошибочно приняли за окаменевшие стволы деревьев, пока один учёный тщательно не очистил находку и не распознал кости динозавра. Сифелли обнаружил их в 1994 году и доставил в Музей им. Сэма Ноубла в Оклахомском университете, но только его студент Мэтью Уэдел присмотрелся как следует и понял, что у него в руках. Завропосейдон известен всего по четырём шейным позвонкам, но они поистине гигантские. Если бы можно было оценить его размеры, предполагая, что эти кости пропорциональны по величине позвонкам жираффатитана, завропосейдон мог достигать 17 м в высоту при вытянутой шее, а значит, был самым высоким динозавром. Его длина составляла около 34 м, вес — примерно 40 т.
 Рис. 17.6.
Рис. 17.6. Сравнение размеров зауропод, например амфицелии, завропосейдона и суперзавра — известны по слишком фрагментарным окаменелостям, чтобы можно было точно определить их величину (рисунок Mary P. Williams)
Если завропосейдон был самым высоким из когда-либо существовавших животных, то некоторые зауроподы были длиннее и массивнее его. Крупнейший экземпляр, по которому известно достаточно много костей для надёжной оценки размеров, — аргентинозавр, найденный в формации Уинкуль в Аргентине (где же ещё?), а именно — в Патагонии, на юге Аргентины (рис. 17.7). Эти окаменелости датируются поздним меловым периодом. Первые кости в 1987 году нашёл фермер, опять же ошибочно принявший их за окаменевшие древесные стволы. Затем собрали образцы, и в 1993 году Хосе Бонапарте и Родольфо Кориа дали динозавру официальное название
Argentinosaurus huinculensis. Имеющиеся фрагменты его скелета — часть позвоночника, тазовая область, несколько рёбер, бедренные кости и правая большая берцовая кость. Пусть костей мало, они огромны. Позвонки просто ошеломительны: каждый высотой более 1,59 м (рис. 17.8,
А), а большая берцовая кость имеет длину 1,55 м. Оценки размеров динозавра, основанные на этих фрагментарных окаменелостях, дают общую длину от 25 до 30 м и вес от 80 до 100 т, хотя новейшие исследования позволяют предположить, что животное могло весить около 50 т. Другая оценка, основанная на более мелком, но полном скелете сальтазавра (другой титанозавр), даёт длину около 30 м и вес от 60 до 88 т. Скелет, установленный в Муниципальном музее Кармен Фунес, имеет 40 м в длину и 7,3 м в высоту, то есть получился даже выше и длиннее первоначальных оценок (рис. 17.7). Таким образом, аргентинозавр может быть самым длинным и крупным сухопутным животным всех времён. Поэтому в настоящее время он считается рекордсменом.
 Рис. 17.7.
Рис. 17.7. Собранный скелет аргентинозавра, выставленный в Муниципальном музее Кармен Фунес в Пласа Уинкуль (Аргентина) (фотография предоставлена R. Coria)

 Рис. 17.8.
Рис. 17.8. Позвонки зауропод: (
А) гигантский позвонок аргентинозавра; (
Б) для сравнения: гораздо более мелкий позвонок жираффатитана (брахиозавра) (см. рис. 17.5) — крупнейшего динозавра, для которого известен и собран практически полный скелет ((
А) фотография автора; (
Б) фотография предоставлена M. Wedel)
Но постойте! Ряд огромных зауропод, живших 97–94 млн лет назад, одновременно с аргентинозавром, имеют почти такие же размеры. Среди них — паралититан из Египта, антарктозавр (рис. 17.9) и аргирозавр из Южной Америки. К сожалению, все они известны максимум по нескольким костям ног, поэтому сравнение их размеров с аргентинозавром в лучшем случае было бы приблизительным. Недавно объявили, что в Аргентине обнаружены ещё более крупные кости. Нашедшие заявили, что это останки «крупнейшего динозавра» (такие экземпляры всегда сопровождаются шумихой), но палеонтологи считают, что это лишь крупная взрослая особь аргентинозавра.
 Рис. 17.9.
Рис. 17.9. Огромные бедренные кости антарктозавра, крупнейшего из известных аргентинозавров. Они больше каких-либо найденных бедренных костей, принадлежавших динозаврам. Для масштаба сфотографирован мальчик Франсиско Новас (фотография предоставлена Fernando Novas)
В 2014 году из Аргентины сообщили о находке ещё одного гигантского зауропода. Первооткрыватели назвали его дредноутом, в честь огромных броненосных кораблей времён Первой мировой войны. «Дредноут» значит неустрашимый: действительно, такой корабль был огромен и хорошо вооружён, ни одно судно ему не угрожало. Скелет дредноута более полный, чем у других зауропод; утверждается, что он сохранился на 70%. Найденные кости относятся в основном к задней части животного и его передним конечностям, но сохранилось очень мало фрагментов шеи и головы. Поэтому длина дредноута оценивается чисто гипотетически. Опять же, первооткрыватели увлеклись раскруткой в СМИ, заявляя, что нашли «самого большого» динозавра весом в 59 т. Палеонтологи отмечают, что экземпляр недостаточно полон для оценки веса и определённо не годится для оценки длины. Многие учёные считают всех титанозавров, имевших разнообразные размеры и извлечённых из меловых пород Аргентины, представителями одного изменчивого рода и, может быть, нескольких видов, которые напрасно подразделили на десятки самостоятельных родов, стараясь привлечь внимание журналистов. Биологам известно, что такие крупные животные обычно живут небольшими популяциями и отличаются сильной внутривидовой изменчивостью; речь не идёт о десятках близкородственных родов, населяющих одну и ту же экосистему.
В настоящее время полагают, что существовали более массивные титанозавры, чем аргентинозавр. Один из них, брухаткайозавр (от санскритского выражения «большое тяжёлое тело» и греческого слова «ящерица»), был найден в нижнемеловых отложениях на территории Индии. Этот динозавр, описанный П. Ядагири и К. Айяасами в 1989 году, мог весить от 175 до 220 т, но впоследствии оценку снизили до 139 т. Если это так, брухаткайозавр был крупнее всех известных зауропод. Однако его окаменелости включали лишь фрагменты тазовых костей, часть бедренной кости и большой берцовой кости, а также предплечья и фрагментов нескольких позвонков. И всё же большая берцовая кость достигала 2 м в длину, то есть была на 29% больше, чем у аргентинозавра. То же касается бедренной кости. Большинство палеонтологов предпочитают не высказываться по поводу брухаткайозавра до тех пор, пока не будет найден более полный материал (что мало вероятно). К сожалению, оригинальные образцы были утрачены, когда здание, в котором они хранились, смыло муссонным наводнением. Поэтому вся информация, оставшаяся о брухаткайозавре, — самая первая публикация с простыми чертежами.
Если этот случай ещё не кажется вам предельно шокирующим и удручающим, послушайте историю о
Amphicoelias fragillimus. Описание этого динозавра основано на единственном позвонке, который в 1877 году нашёл один из пионеров палеонтологии Эдвард Дринкер Коп (он и дал название этому динозавру). Коп опубликовал рисунок с изображением данного экземпляра, и, если верить его измерениям, животное было просто колоссальным! Единственный позвонок, сохранись он полностью, имел бы 2,7 м в высоту. Если соотнести эту величину с размерами позвонков других зауропод, амфицелия могла достигать 40–60 м в длину и весить до 122 т, то есть превосходила бы по размеру любого динозавра, кроме брухаткайозавра (см. рис. 17.6). К сожалению, материал по амфицелии был утрачен вскоре после смерти Копа. Возможно, позвонок просто рассыпался в переполненном запаснике, поскольку консерванты тогда ещё не использовались, или образец кто-то нечаянно разбил при транспортировке коллекции, уже после смерти Копа. Таким образом, мало того, что два кандидата на звание крупнейшего сухопутного животного известны по некачественным окаменелостям, так ещё и сами
окаменелости утрачены! Поэтому пальма первенства по-прежнему у аргентинозавра — пока его не потеснят, найдя более убедительные материалы.
Живой динозавр в Конго?
Существует современная легенда о том, что в бассейне реки Конго по-прежнему обитает динозавр-зауропод. Это животное, известное под именем мокеле-мбембе, фигурирует во многих книгах, сообщениях СМИ, «документальных» телефильмах и даже в голливудской киноленте «Динозавр. Тайна затерянного мира» (1985). О нём рассказывали многие люди, поэтому оно почти так же знаменито, как лох-несское чудовище и снежный человек. Однако, если изучить вопрос подробнее, убеждаешься, что всё это — просто пыль в глаза. Мы с Дэниэлом Локстоном показали, что какие-либо физические доказательства в пользу реальности мокеле-мбембе отсутствуют. Большинство «доказательств» сводится к рассказам очевидцев из числа местных жителей, причём эти рассказы перевели и интерпретировали американские исследователи (почти всегда это миссионеры или современные креационисты, а не биологи). Такие свидетельства крайне недостоверны, поскольку многие аборигены не видят разницы между мифическими и реальными, с нашей точки зрения, животными. Кроме того, многие рассказы расплывчаты, противоречивы и бесполезны в научном отношении. Некоторые пытаются распознать в них описание стегозавра или трицератопса, а не зауропода. Кто-то даже описывает носорога, хотя это животное неизвестно жителям бассейна Конго, поскольку носороги обитают в саванне, а не в джунглях. Многие рассказы подозрительны, так как западные исследователи часто показывают туземцам изображения животного и спрашивают, соответствует ли картинка их наблюдениям — задают наводящие вопросы. Во многих культурах считается нормальным говорить гостю то, что он хочет услышать, из вежливости. Важнее всего, что, согласно последним психологическим исследованиям, «свидетельство очевидца» практически неприменимо в качестве показаний (даже в суде). Человек — плохая видеокамера. Мы с такой готовностью «видим» то, что хотели увидеть, и приукрашиваем изначально увиденное, исходя из собственных ожиданий, так легко выдаём воображаемое за действительное, что ни один учёный не примет свидетельство «очевидца» иначе, как его личный опыт (а возможно, обман зрения или галлюцинацию).
Кроме того, с этими свидетельствами и данными сопряжена масса проблем, из-за которых существование мокеле-мбембе представляется крайне маловероятным. Все фотографии и видеозаписи сделаны с такого большого расстояния и столь размыты, что невозможно понять, что на них запечатлено, и тем более считать их доказательствами существования животного. На снимках, которые удаётся распознать, обычно оказываются гиппопотамы, люди в каноэ либо другие нечёткие объекты без характерных черт. Популяционная экология подсказывает, что таким крупным животным, как зауроподы, нужны огромные территории для обитания, причём сама популяция должна быть велика, в ней должны быть и взрослые, и молодые особи. Однако мы располагаем лишь свидетельствами очевидцев и плохим видео — нет ни костей, ни скелетов, ни каких-либо других физических доказательств.
Идёт время, всё больше людей ищут и не находят мокеле-мбембе, поэтому гипотеза становится всё слабее. На самом деле «неизведанные джунгли» в бассейне Конго — миф. Профессиональные биологи постоянно путешествуют по бассейну Конго, но они никогда не видели мокеле-мбембе и не слышали рассказов о нём. Сообщения о динозавре убедительны только для легковерных миссионеров, не разбирающихся в биологии. На картах Google Earth все желающие могут рассмотреть любой регион из космоса и без труда заметить крупных животных. Если вбить в Google Earth координаты 10.903497 N, 19.93229 E, вы увидите слонов с неплохой детализацией. Определённо, крупное животное вроде мокеле-мбембе тоже давно заметили бы, если бы огромные стада этих ящеров бродили в бассейне Конго.
Сохранилась отличная палеонтологическая летопись зауропод, а их огромные кости хорошо фоссилизируются. Поэтому сам факт, что
ни одной ископаемой кости зауропод не найдено в отложениях моложе 65 млн лет, убедительно свидетельствует: зауроподы не дожили до наших дней (хотя есть немало пластов с подходящими экологическими условиями, в которых хорошо фоссилизируются крупные млекопитающие).
Наконец, в истории о мокеле-мбембе есть и кое-что совсем неправдоподобное. «Очевидцы», рассказывавшие о динозавре, описывали зауропода в таком виде, какой был популярен в 1905 году, когда всеобщее внимание было приковано к первым скелетам и изображениям. Но такие животные никогда не существовали. Научные исследования показали, что зауроподы не были медлительными и неповоротливыми тварями, таскавшими за собой хвост и прятавшимися в болотах, а вытягивали хвост горизонтально и жили на суше вдоль побережья, не в воде. В рассказах же о мокеле-мбембе это животное прячется в реке Конго и может затаиваться там часами. На самом деле зауропод не мог даже целиком уйти под воду, так как у него вдоль позвоночника было слишком много воздушных карманов. Они не умели нырять и тем более часами сидеть под водой.
С мифом о мокеле-мбембе связана одна странность: его поисками заняты священники-креационисты, а не профессиональные зоологи. Несколько лет назад меня попросили выступить в качестве «скептика» в одном из эпизодов сериала «Охотники на монстров», посвящённом мокеле-мбембе. Съёмка получилась поистине странной, поскольку большую часть времени продюсеры потратили на то, чтобы заставить меня говорить вещи, которые впоследствии можно было бы подать как аргументы в пользу существования этого существа. Они даже пытались поймать меня на слове на камеру, вручая бесформенный кусок гипса и рассчитывая, что я распознаю в нём «след динозавра». Когда я смотрел эту передачу, больше всего меня удивили два «охотника» на мокеле-мбембе, история о похождениях которых заняла основное время ленты. Они показали себя некомпетентными зоологами, понятия не имеющими, чем занимаются, и не умеющие обращаться со своим вычурным оборудованием; говорили какие-то странные вещи о крошечной норе в отвесном речном береге, будто гигантский зауропод мог зарыться в него, полностью спрятаться и оставить лишь небольшое отверстие для дыхания.
Позже я выяснил, что оба «исследователя» были священниками-креационистами, не имевшими систематического образования в области зоологии. Один из них, Уильям Гиббон, совершил много экспедиций в Конго, потратил кучу денег и не добился никаких результатов. Почему-то эти люди считают, что открытие крупного динозавра опровергло бы теорию эволюции — невзирая на горы доказательств, которые её подтверждают!
Охота на мокеле-мбембе — не бесполезные поиски живого ископаемого, интересные любителям. «Исследователи», тратящие на это своё время, руководствуются антинаучными целями, их данным и интерпретациям нельзя верить. Подобные поиски — часть глобальной деятельности креационистов, стремящихся дискредитировать доказательства теории эволюции и любыми средствами расшатать её научные основы. Эти потуги нельзя просто игнорировать или относиться к ним снисходительно. Напротив, они требуют пристального внимания научного сообщества, поскольку угрожают науке.
Лучше один раз увидеть!
Во многих музеях естественной истории по всему миру выставлены оригинальные слепки скелетов зауропод. В США такой оригинальный материал есть в Американском музее естественной истории (Нью-Йорк) — апатозавр и барозавр, Музее естественной истории им. Карнеги (Питтсбург) — апатозавр и первый диплодок, Национальном музее естественной истории Смитсоновского института (Вашингтон, округ Колумбия) — диплодок и камаразавр, Музее естественной истории округа Лос-Анджелес (Лос-Анджелес) — маменчизавр, а также в Музее естественной истории им. Пибоди при Йельском университете (Нью-Хейвен, Коннектикут) — апатозавр.
Практически полные скелеты жираффатитана (брахиозавра) и дикреозавра выставлены в Музее естественной истории (Музее Гумбольдта) в Берлине, а также в Чикаго, где слепки жираффатитана установлены прямо у выхода из Музея естественной истории под открытым небом и в международном аэропорту О’Хара. Позвонки завропосейдона экспонируются в Музее им. Сэма Ноубла при Оклахомском университете в городе Норман. Реконструированные скелеты аргентинозавра можно увидеть в Аргентине, в Муниципальном музее Кармен Фунес (Пласа Уинкуль) и в Аргентинском музее естественных наук Бернардино Ривадавия в Буэнос-Айресе. Слепок такого скелета также выставлен в Музее естественной истории Фернбанк (Атланта).
Дополнительная литература
Brett-Surman, M. K., Thomas R. Holtz Jr., and James O. Farlow, eds. The Complete Dinosaur. 2nd ed. — Bloomington: Indiana University Press, 2012.
Curry Rogers, Kristina, and Jeffrey A. Wilson, eds. The Sauropods: Evolution and Paleobiology. — Berkeley: University of California Press, 2005.
Fastovsky, David E., and David B. Weishampel. Dinosaurs: A Concise Natural History. 2nd ed. — Cambridge: Cambridge University Press, 2012.
Holtz, Thomas R., Jr. Dinosaurs: The Most Complete Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. — New York: Random House, 2007.
Klein, Nicole, Kristian Remes, Carole T. Gee, and P. Martin Sander, eds. Biology of the Sauropod Dinosaurs. Understanding the Life of Giants. — Bloomington: Indiana University Press, 2011.
Loxton, Daniel, and Donald R. Prothero. Abominable Science: The Origin of Yeti, Nessie, and Other Cryptids. — New York: Columbia University Press, 2013.
Paul, Gregory S. The Princeton Field Guide to Dinosaurs. — Princeton, N. J.: Princeton University Press, 2010.
Sander, P. Martin. An Evolutionary Cascade Model for Sauropod Dinosaur Gigantism — Overview, Update and Tests //
PLoS ONE. — 2013. — Vol. 8. — e78573.
Sander, P. Martin, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva-Marie Griebeler, Hanns-Christian Gunga, Jürgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas Tütken, Oliver Wings, and Ulrich Witzel. Biology of the Sauropod Dinosaurs: The Evolution of Gigantism //
Biological Reviews of the Cambridge Philosophical Society. — 2011. — Vol. 86. — P. 117–155.
Tidwell, Virginia, and Kenneth Carpenter, eds. Thunder-Lizards: The Sauropodomorph Dinosaurs. — Bloomington: Indiana University Press, 2005.
18. Перо в камне
Первая птица
Археоптерикс
И если бы весь крестец недосиженного цыплёнка <... > вдруг увеличился бы, окостенел и окаменел в неизменном виде, мы получили бы последнее переходное звено между птицами и рептилиями; причём ни одна черта такого существа не помешала бы нам отнести его к динозаврам.
Томас Генри Гексли. «Further Evidence of the Affinity between the Dinosaurian Reptiles and Birds»
Творения природы
Более 300 лет каменщики высекали куски прекрасного, затейливо-слоистого кремового известняка в карьерах Зольнхофен близ города Эйхштетт в Южной Германии. Эти удивительные породы были такими тонкозернистыми (без видимых окаменелостей, типичных для большинства известняков), что славились на весь мир в качестве сырья для изготовления литографий, протравливаемых кислотой. В них не было ни изъянов, ни примесей, ни каких-либо включений, способных испортить тонкие детали вырезанных вручную пластин. Многие великие произведения искусства созданы из этой породы. Она использовалась для тиснения самых первых литографий на заре существования печатных книг, в том числе для легендарных работ Альбрехта Дюрера и других художников. Благодаря абсолютно ровному цвету, отсутствию рисунка и зернистости эта порода до сих пор остаётся популярным строительным материалом; сегодня её можно заказать по Интернету.
К середине XIX века в Зольнхофене развернулись масштабные горные работы: множество людей трудились не покладая рук, чтобы найти неповреждённые известняковые обнажения, из которых можно было бы высекать большие плоские куски для изготовления печатных форм и строительного камня. Иногда, раскалывая плиты по плоскости напластования, они находили художества совершенно иного рода — прекрасно сохранившиеся окаменелости разных существ, в том числе разнообразных костистых рыб, отдельных ракообразных и мечехвостов, офиур. Там же встречались окаменелости динозавра компсогната, а также были впервые найдены хорошо сохранившиеся экземпляры птеродактиля, описанные натуралистами ещё в 1784 году. Каменотёсы не искали их специально, но когда случайно находили, это была приятная награда за долгие часы каторжного труда. Некоторые ископаемые были так красивы, что их продавали коллекционерам и богачам, собиравшим эти дары природы для удовольствия или из научного интереса.
Однажды в 1860 году каменотёс обнаружил в известняке удивительную вещь — чёткий отпечаток пера, очень напоминавший асимметричные маховые перья современных птиц. Спустя некоторое время данный образец попал в руки выдающемуся палеонтологу Кристиану Герману Эриху фон Майеру, который уже описал большинство зольнхофенских динозавров и птеродактилей, а также древнего динозавра платеозавра (см. главу 17). В 1860 году Майер, имея всего одно ископаемое перо, дал животному официальное научное название
Archaeopteryx lithographica («древнекрыл из литографических камней»).
Как повезло Дарвину
Спустя несколько месяцев почти полный скелет археоптерикса нашли в каменоломне близ Лангенальтхайма в Германии (рис. 18.1). Затем этой находкой расплатились за медицинские услуги с местным врачом Карлом Геберлейном. В образце почти отсутствовали шея и голова; это была мешанина из костей, но чётко просматривались отпечатки перьев вокруг скелета, напоминавшего динозавровый. Разные немецкие музеи хотели приобрести этот экземпляр, поэтому Геберлейн принял максимально щедрое предложение — 700 фунтов (около 72 тыс. долларов в пересчёте на современные цены и целое состояние по тем временам!), сделанное Британским музеем естественной истории. Этот образец стал известен как Лондонский экземпляр, поскольку до сих пор там и находится. Когда окаменелость попала в Лондон, её внимательно изучил выдающийся британский анатом и палеонтолог Ричард Оуэн. Он уже был известен как автор описаний многих окаменелостей и термина «динозавры». Оуэн взялся за работу и в 1863 году опубликовал подробное описание этого экземпляра. При всей его фрагментарности, учёный не мог проигнорировать тот факт, что кости существа были как у рептилий, хотя крылья оперены.
 Рис. 18.1.
Рис. 18.1. «Лондонский экземпляр» археоптерикса (фотография из фонда Wikimedia Commons)
Это открытие стало настоящим подарком судьбы для другого британского естествоиспытателя, Чарльза Дарвина. Его неоднозначная новая книга «Происхождение видов» была опубликована в 1859 году — всего двумя годами ранее. Несмотря на то, какие убедительные доказательства он приводил в пользу реальности эволюции, ему приходилось извиняться за отсутствие хороших «промежуточных» окаменелостей, которые подтверждали бы его теорию. Археоптерикс, который и «хронологически» отлично соответствовал данной теории, оказался первоклассным переходным звеном, подкреплявшим построения Дарвина; учёный был просто в восторге. Он и представить не мог такого идеального примера, демонстрировавшего, как рептилии могли превратиться в птиц, совершенно иной класс. В четвёртом издании «Происхождения видов» Дарвин победоносно заявлял:
... ещё не так давно палеонтологи держались того мнения, что весь класс птиц появился внезапно в эоценовый период, а теперь мы знаем, по свидетельству проф. Оуэна, что птица несомненно существовала в эпоху отложения верхнего зелёного песчаника; а совсем недавно в юрских сланцах Золенгофена была найдена странная птица Archaeopteryx, с длинным, как у ящерицы, хвостом, на каждом позвонке которого сидела пара перьев, и с крыльями, снабжёнными двумя свободно выступающими когтями. Это открытие едва ли не яснее всякого другого показало, как мало мы ещё знаем о древних обитателях Земли.
Однако Оуэн верил не в дарвиновскую эволюцию, а в трансмутацию — процесс, который описал сам. В 1863 году, характеризуя ископаемого археоптерикса, он старательно замалчивал или отвергал все напрашивавшиеся, явные связи между птицами и пресмыкающимися. Томас Генри Гексли, молодой и резкий учёный, блистательно отстаивавший эволюционные взгляды и получивший за это прозвище «бульдог Дарвина», раскритиковал Оуэна за его недоработку и потребовал признать очевидное. Он доказывал, что археоптерикс не только замечательно подходит на роль недостающего звена между рептилиями и птицами, но и — что гораздо важнее — по строению костей почти не отличается от динозавра. На самом деле оказалось, что ещё один экземпляр археоптерикса изначально был неверно описан как мелкий зольнхофенский динозавр компсогнатус, пока столетие спустя Джон Остром из Йельского университета не рассмотрел экземпляр внимательнее и не заметил на нём перья.
Всё больше экземпляров
Решающий довод в этом споре появился, когда крестьянин Якоб Нимайер в 1874 году нашёл близ Блюмберга в Германии наиболее хорошо сохранившийся из всех известных экземпляр археоптерикса (рис. 18.2). Чтобы выручить денег на покупку коровы, он продал эту удивительную окаменелость трактирщику Иоганну Дёрру. Тот, в свою очередь, продал экземпляр Эрнсту Отто Геберлейну — сыну того самого врача, который двенадцатью годами ранее продал окаменелость археоптерикса Британскому музею. Это самый знаменитый и чаще всего фотографируемый из 12 известных экземпляров: он сохранился почти полностью, при этом распластан в скале, поэтому все его перья хорошо видны, а голова и шея откинуты назад. Типичная поза умирающих животных, поскольку в агонии сокращается выйная связка, удерживающая голову и шею.
 Рис. 18.2.
Рис. 18.2. Берлинский экземпляр археоптерикса (фотография из фонда Wikimedia Commons)
Когда Геберлейн в 1877 году стал подыскивать покупателей на свою невероятную находку, многие научные институты стремились её приобрести. Интересовались не только британцы, но и йельский палеонтолог Отниел Чарльз Марш. Однако немцы не желали так просто делиться своим достоянием с иностранцами после того, как первый археоптерикс от них упорхнул. Берлинский музей естественной истории (Музей Гумбольдта), получив финансирование от Эрнста Вернера фон Сименса, чья компания по-прежнему является гигантом во многих отраслях, приобрёл этот экземпляр за 20 тыс. золотых марок (примерно 21 тыс. долларов в пересчёте на современные цены). Теперь данная окаменелость известна под именем Берлинский экземпляр. Её многократно изучали, и сегодня именно она является источником большинства наших знаний об археоптериксе. Берлинский экземпляр — ещё более удачный пример недостающего эволюционного звена, чем Лондонский, поскольку он хорошо сохранился и чётко демонстрирует смесь птичьих и динозавровых признаков.
Хотя окаменелости археоптерикса редки (примерно за 200 лет найдено всего 12 экземпляров), после официального объявления о Берлинском экземпляре в 1877 году нашлись и другие. Одну окаменелость (найдена в 1855 году, сейчас находится в Музее Тейлора в Харлеме (Нидерланды)) сначала ошибочно приняли за крыло птерозавра, а первый экземпляр, идентифицированный как археоптерикс, обнаружили в известняках позднее. В 1970 году Остром внимательнее рассмотрел Голландский экземпляр и осознал, что это кость из крыла археоптерикса, а не птерозавра; на ней были едва заметные отпечатки перьев. Другой экземпляр (в Юрском музее города Айхштетт) нашли в 1951 году близ Воркерсцелля в Германии. Он является одним из самых маленьких, но наиболее хорошо сохранившихся скелетов. Ещё одна окаменелость, найденная в 1992 году, в 1999-м была продана в Палеонтологический музей Мюнхена за 1,9 млн немецких марок (примерно 1,3 млн долларов в пересчёте на современные цены). Этот скелет тоже почти полный, хотя при фоссилизации он оказался свёрнут практически пополам. Торс ещё одного экземпляра (голова и хвост не сохранились) нашли в 1956 году близ Лангенальтхайма. Он долго экспонировался в Музее Максберга, пока его владелец, Эдуард Опич, не забрал окаменелость обратно. После смерти Опича найти экземпляр не удалось — его украли либо продали на чёрном рынке.
Два других фрагментарных экземпляра по-прежнему находятся у частных владельцев. Дейтингский экземпляр (из дейтингских отложений, чуть более молодых, чем зольнхофенские) показывали очень недолго. Другая окаменелость, временно переданная в пользование Музею Бюргермайстер-Мюллер в Зольнхофене, — просто крыло. Ещё один важный экземпляр длительный период времени находился в частном владении, пока не был подарен крошечному Музею «Центр динозавров» в отдалённом городе Термополис (штат Вайоминг). Это одна из самых полных окаменелостей, с хорошо сохранившейся головой и ступнями, но без нижней челюсти и шеи. Наконец, в 2011 году объявили об открытии двенадцатого экземпляра. Он находится у частного владельца и описан недавно.
Птица... или динозавр?
Как правильно понял Гексли ещё в 1860-е годы, скелет археоптерикса схож со скелетом динозавра — настолько, что один экземпляр ошибочно приняли за маленького зольнхофенского теропода компсогнатуса (рис. 18.3). Как и большинство динозавров (но не современных птиц), археоптерикс имел длинный костистый хвост, зубастый череп с большим количеством полостей, динозавровые (не птичьи) позвонки, длинную узкую лопатку, тазовую кость, напоминающую как более позднюю птичью, так и типично динозавровую, а также обладал уникальной динозавровой и в то же время птичьей специализацией конечностей. Больше всего поражает запястье. У всех птиц и у некоторых хищных динозавров, например у дромеозавров (велоцираптор, дейноних и им подобные), запястная кость имеет форму полумесяца и образована слиянием множества костей; такая черта уникальна для этих животных. Кость служит основным шарниром для движения запястья, благодаря чему дромеозавр мог вытягивать лапу и хватать добычу быстрым хлёстким ударом книзу. Именно такое движение птица совершает в полёте, когда машущее крыло идёт вниз. Археоптерикс имел те же три пальца (большой, указательный и средний), что и большинство динозавров, причём указательный палец был намного длиннее остальных. Кроме того, когти археоптерикса очень напоминали кости хищных динозавров.
 Рис. 18.3.
Рис. 18.3. Сравнение скелетов небольшого динозавра орнитолестеса, археоптерикса и голубя (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 12.6)
Задние конечности археоптерикса тоже обладали многими признаками, типичными для динозавров. Самые поразительные черты имеет голеностоп. У всех динозавров, птерозавров и птиц уникальное строение голеностопа, так называемая цевка: вместо типичного для позвоночных голеностопа, который находится между большеберцовой костью и первым рядом костей плюсны (как ваша лодыжка), у них срастаются первый и второй ряд костей плюсны — внутри цевки. Таким образом, верхний ряд костей плюсны почти не функционален, у многих птиц и динозавров он просто сливается с кончиком большеберцовой кости, образуя небольшой костный нарост. Когда в следующий раз будете обгладывать куриную или индюшачью ножку (это и есть берцовая кость), обратите внимание на несъедобный хрящик в верхней, менее мясистой части ножки — это реликт, свидетельствующий, что птицы произошли от динозавров! Кроме того, в передней части первого ряда костей плюсны есть костная шпора, отходящая от большеберцовой кости, — ещё одна уникальная черта некоторых динозавров и птиц. Наконец, элементы и структура пальцев ног, а также короткие большие пальцы характерны именно для хищных динозавров и птиц. У археоптерикса не было крупного «птичьего» противопоставленного большого пальца, благодаря которому он мог бы хвататься за ветки и карабкаться по ним. Однако последние исследования показывают, что на задних лапах археоптерикса всё-таки был небольшой «ударный коготь», который вы могли видеть у велоцирапторов в фильме «Парк юрского периода».
При наличии всех этих фактов, свидетельствующих, что археоптерикс — оперённый динозавр, почему мы считаем его птицей? На самом деле у него всего несколько уникальных «птичьих» черт, которые не встречаются у хищных динозавров: большой палец на ноге почти полностью отставлен назад; по краям зубов нет таких зазубрин, как на мясницком ноже; хвост сравнительно короток для динозавра, а передние конечности длиннее, чем у большинства других хищных динозавров. Все остальные черты археоптерикса, в том числе перья и сросшаяся ключица (вилочка), уже найдены у других динозавров. Некоторые считают, что перья археоптерикса были более развиты, чем у других динозавров, и имели асимметричную форму: стержень шёл не по центру, а по краю пера. Это значит, что археоптерикс мог летать, хотя и не так хорошо, как большинство современных птиц.
Птицы взлетают
Археоптерикс произвёл революцию — это первая переходная окаменелость, найденная после публикации дарвиновского «Происхождения видов». На его примере видно, как некоторые динозавры превратились в птиц. Но палеонтологическая летопись первоптиц стремительно дополняется, особенно последние 30 лет, когда огромное количество прекрасно сохранившихся окаменелостей найдено в Китае. Самые революционные открытия были сделаны в ляонинских отложениях на северо-востоке Китая и датируются ранним меловым периодом; эти залежи окаменелостей стали одними из важнейших в мире. В тонких озёрных сланцах окаменелости сохраняются поразительно хорошо: видны контуры тела, перья и мех. Также там есть полностью сочленённые скелеты, не потерявшие ни одной кости. Иногда удаётся определить даже цвет перьев и содержимое желудка.
В течение прошлого десятилетия каждые несколько месяцев объявляли о новом крупном открытии, сделанном в этих отложениях, переворачивавшем прежние представления о птицах и динозаврах. Самые удивительные из всех окаменелостей — явно нелетающие динозавры, ещё не превратившиеся в птиц, но имевшие развитые перья (рис. 18.4). Среди них такие невероятно хорошо сохранившиеся экземпляры, как синозавроптерикс, протархеоптерикс, синорнитозавр, каудиптерикс, крупный теропод бэйпяозавр и крошечный микрораптор.

 Рис. 18.4.
Рис. 18.4. Синозавроптерикс — нелетающий оперённый динозавр из ляонинских отложений, ещё не превратившийся в птицу: (
А) окаменелость; (
Б) реконструкция внешнего вида (илл. предоставлены M. Ellison и M. Norell, Американский музей естественной истории)
У большинства этих динозавров не было маховых перьев и других признаков, которые свидетельствовали бы о пригодности оперения для полёта. На окаменелостях просматриваются следы перьев, являвшихся обычной чертой хищных динозавров (а также большинства других ящеров, вероятно, даже птерозавров). Таким образом, перья развивались не для полёта, а, предположительно, для утепления и лишь потом превратились в летательное приспособление. В 2003 году Ричард Прам и Алан Браш опубликовали статью, где полностью переосмыслили происхождение перьев. Они показали, что перья — не видоизменившиеся чешуйки, как считалось ранее, а развиваются из примерно таких же эмбриональных зачатков, что и чешуя, однако их развитие контролируют иные гены (рис. 18.5). Перья первого типа — простые полые заострённые стержни, появившиеся у примитивных динозавров, а также имевшиеся у другой эволюционной ветви, к которой относятся трицератопсы и их родичи. Перья второго типа — пух без опахала, как у динозавра синозавроптерикса. У перьев третьего типа есть и пух, и стержень, но нет бородок, которые скрепляли бы их подобно застёжке-липучке. Такие перья были у
Yutyrannus и, скорее всего, у королевского тираннозавра. Перья второго и третьего типа имеются у крупного динозавра бэйпяозавра, то есть они были у наиболее развитых хищных динозавров, например у дромеозавров. У перьев четвёртого типа есть бородки, связывающие опахала в гладкую поверхность, но стержень идёт через центр пера. Такие перья наблюдаются у каудиптерикса, и это позволяет предположить, что данная черта была свойственна сравнительно высокоразвитым хищным динозаврам: орнитомимозаврам, овирапторам и дромеозаврам. Классическое асимметричное маховое перо, в котором стержень пролегает у переднего края опахала, появилось у археоптерикса, и именно поэтому многие учёные считают его одной из первых промежуточных форм между динозаврами и птицами — одним из тех, чьи перья стали меняться для настоящего полёта.
 Рис. 18.5.
Рис. 18.5. Эволюция перьев у динозавров и птиц (рисунок Карла Бьэлла на основе материала из статьи
Richard O. Prum and Alan H. Brush. Which Came First, the Feather or the Bird? //
Scientific American. — March 2003 и из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 12.9)
Следуя от археоптерикса по эволюционному древу птиц (рис. 18.6), мы подходим к раховахису (рис. 18.7,
А), жившему в меловом периоде на Мадагаскаре. Размером он был примерно с ворону, на задней лапе имел серповидный коготь, обладал длинным костистым хвостом, зубами и многими другими чертами динозавров, но в то же время и такими птичьими признаками, как сращивание задних позвонков с крестцом (синсакрум), отверстиями для кровеносных сосудов во всех позвонках, а также воздушными мешками, которые есть у современных птиц. На пальцах у рановахиса имелись маховые бугорки (небольшие костные выросты, к которым прикрепляются маховые перья). Это позволяет предположить, что он был оперён и умел летать (что неудивительно). Была у него и фибула (малоберцовая кость), не достававшая до цевки. У птиц фибула редуцировалась до крошечной костяной щепки, которую вы кусаете, обгладывая куриную или индюшачью ножку. У археоптерикса была полноценная фибула, как у динозавров.
 Рис. 18.6.
Рис. 18.6. Родословное древо птиц мезозойской эры (иллюстрация предоставлена L. Chiappe, Музей естественной истории округа Лос-Анджелес)

 Рис. 18.7.
Рис. 18.7. Птицы мелового периода: (
А) раховахис с Мадагаскара; (
Б) реконструкция синорниса из Китая ((
А) по
Catherine A. Forster et al. The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar //
Science. — 1998. — March 20; © 1998 Американская ассоциация по содействию научному прогрессу; (
Б) по
Paul C. Sereno and Rao Chenggang. Early Evolution of Avian Flight and Perching: New Evidence from the Lower Cretaceous of China. //
Science. — 1992. — February 14. — Fig. 2; © 1992 Американская ассоциация по содействию научному прогрессу)
Следующий виток эволюции — конфуциусорнис и его родичи. Это первые птицы, имевшие беззубый клюв, а также уникальный признак, характерный для всех высших птиц, — пигостиль, кость, образующуюся в результате слияния всех динозавровых хвостовых позвонков в единую «гузку». У этих более развитых птиц также увеличилось количество задних спинных позвонков, срастающихся в синсакрум, имелись более длинные кости, укреплявшие плечо и улучшавшие способность к полёту. Недавно путём эмбриологических экспериментов удалось разблокировать птичьи гены, подавляющие развитие хвостовой кости и гарантирующие, что эта кость остаётся короткой. Получилась курица с длинным костистым хвостом, как у её предков-динозавров.
Вслед за этой переходной формой выходим ещё на одну эволюционную ветвь, которая ведёт к вымершим энанциорнисовым, или противоптицам. Они так названы потому, что фоссилизированные кости конечностей ориентированы в противоположном направлении, чем у современных птиц, а плечевые кости находятся в странном состоянии (см. рис. 18.6). К ним относится иберомезорнис из мелового периода, найденный в местечке Лас-Хойяс в Испании; синорнис из Китая (см. рис. 18.7,
Б); гобиптерикс из Монголии; энанциорнис из Аргентины и многие другие. Все эти птицы были более специализированными, чем археоптерикс, рахонавис или конфуциусорнис, — в том смысле, что у них уменьшилось количество туловищных позвонков, имелась гибкая вилочковая кость, плечевой сустав был лучше приспособлен для полёта, кости верхней конечности слились в одну так называемую пряжку, а кости пальцев в основном слились в цельный элемент (ту часть куриного крылышка, которую мы никогда не едим).
Продолжая путь по родословному древу, добираемся до нескольких птиц мелового периода: вуруна с Мадагаскара, патагоптерикса из Аргентины, а также хорошо известных водоплавающих птиц гесперорниса и ихтиорниса, найденных в меловых отложениях Канзаса. Эти птицы имеют не менее 15 ярко выраженных специализированных эволюционных черт, среди которых потеря брюшных рёбер (гастралий); переориентация лобковой кости в то положение, какое она занимает у современных птиц (параллельно седалищной кости); уменьшение количества туловищных позвонков и многие другие признаки крыла и плеча, улучшавшие способность к полёту. Ихтиорнис ещё более близок современным птицам, так как имел киль — вырост грудины, к которому прикрепляются летательные мышцы, а также особое утолщение на плечевом суставе, повышающее гибкость крыла. Наконец, группа, к которой относятся все современные представители класса птиц, характеризуется полной утратой зубов и некоторыми другими анатомическими специализациями, в частности слиянием костей плюсны с образованием цевки.
* * *
С тех пор как был найден первый ископаемый археоптерикс, пройден большой путь. Открытие этого животного сыграло важную роль, подкрепив теорию эволюции Дарвина. На протяжении десятилетий он оставался центральной темой всех споров, связанных с происхождением птиц и полёта. Теперь археоптерикс — один из сотен удивительных экземпляров ископаемых птиц, живших в эпоху динозавров. Эти животные заставили нас полностью пересмотреть представления о динозаврах, особенно о птицах. Динозавры не вымерли, сейчас они скачут в птичьих клетках и летают в наших садах. Поэтому, когда в следующий раз увидите «оперённого динозавра», взмывающего в небо, восхититесь, как эволюция превратила ужасного хищника велоцираптора во множество удивительных птиц, от страусов до колибри. Все они — современные оперённые динозавры.
Лучше один раз увидеть!
Практически все старинные зольнхофенские каменоломни сейчас находятся в частной собственности, там нельзя ничего собирать без разрешения владельца. Поскольку почти за 200 лет было найдено всего 12 экземпляров археоптерикса, шансы найти ещё один крайне малы.
Большинство оригинальных экземпляров археоптерикса исключительно ценны, а некоторые до сих пор находятся в частных коллекциях и не выставлены на всеобщее обозрение. Например, окаменелость, которая ранее выставлялась в Максбергском музее, сегодня утрачена; Дейтингский экземпляр не экспонируется; вы нигде не увидите и окаменелость, описанную совсем недавно, — она у частного владельца. Точные слепки выставлены во многих музеях естественной истории, даже есть в продаже. В некоторых музеях, например в Американском музее естественной истории в Нью-Йорке, выставлены слепки не только Берлинского и Лондонского экземпляров, но и большинства общедоступных окаменелостей археоптерикса.
Насколько мне известно, в музеях есть следующие оригинальные окаменелости археоптерикса:
• Лондонский экземпляр — в Музее естественной истории в Лондоне (см. рис. 18.1);
• Берлинский экземпляр (за непробиваемым стеклом) в Музее естественной истории (Музее Гумбольдта) в Берлине (см. рис. 18.2);
• экземпляр из Термополиса — в Центре Динозавров штата Вайоминг (город Термополис);
• частично сохранившийся экземпляр — в Мюнхенском палеонтологическом музее;
• почти полный экземпляр — в Юрском музее (Айхштетт, Германия);
• образец крыла — в Музее Бюргермайстер-Мюллер (Зольнхофен, Германия);
• образец крыла — в Музее Тейлора (Харлем, Нидерланды).
Дополнительная литература
Chiappe, Luis M. The First 85 Million Years of Avian Evolution //
Nature. — 1995. — November 23. — P. 349–355.
Chiappe, Luis M., and Gareth J. Dyke. The Mesozoic Radiation of Birds //
Annual Review of Ecology and Systematic. — 2002. — Vol. 33. — P. 91–124.
Chiappe, Luis M., and Lawrence M. Witmer, eds. Mesozoic Birds: Above the Heads of Dinosaurs. — Berkeley: University of California Press, 2002.
Currie, Philip J., Eva B. Koppelhus, Martin A. Shugar, and Joanna L. Wright, eds. Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. — Bloomington: Indiana University Press, 2004.
Gauthier, Jacques, and Lawrence F. Gall, eds. New Perspectives on the Origin and Early Evolution of Birds. — New Haven, Conn.: Yale University Press, 2001.
Norell, Mark. Unearthing the Dragon: The Great Feathered Dinosaur Discovery. — New York: Pi Press, 2005.
Ostrom, John H. Archaeopteryx and the Origin of Birds //
Biological Journal of the Linnean Society. — 1976. — Vol. 8. — P. 91–182.
Ostrom, John H. Archaeopteryx and the Origin of Flight //
Quarterly Review of Biology. — 1974. — Vol. 49. — P. 27–47.
Padian, Kevin, and Luis M. Chiappe. The Origin of Birds and Their Flight //
Scientific American. — 1998. — February. — P. 28–37.
Prum, Richard O., and Alan H. Brush. Which Came First, the Feather or the Bird? //
Scientific American. — 2003. — March. — P. 84–93.
Shipman, Pat. Taking Wing: Archaeopteryx and the Evolution of Bird Flight. — New York: Simon & Schuster, 1988.
19. Почти млекопитающее
Происхождение млекопитающих
Тринаксодон
Из всех великих переходов между основными структурными уровнями среди позвоночных переход от примитивнейших амниот до примитивнейших млекопитающих сопровождается наиболее полной и непрерывной палеонтологической летописью, простирающейся от среднего пенсильванского до позднего триасового периода, и занимает от 75 до 100 миллионов лет.
Джеймс Хопсон. «Synapsid evolution and the radiation of non-eutherian mammals»
Протомлекопитающие
Один из наиболее полных и максимально документированных переходов в палеонтологической летописи — последовательность, рассказывающая о происхождении млекопитающих от самых примитивных амниот (рис. 19.1). Практически для каждого этапа найдены сотни отлично сохранившихся экземпляров. Систематическое название всех ископаемых протомлекопитающих — синапсиды. Это таксономическая группа, к которой относятся не только предки млекопитающих, но и сами млекопитающие. Палеонтологи больше не используют устаревший термин «зверообразные рептилии», так как эволюционная линия млекопитающих, первыми представителями которой были археотирис и протоклепсидропс, жившие в позднем карбоне, возникла одновременно с эволюционной линией рептилий (где рептилии — это черепахи, змеи, ящерицы, крокодилы и их родичи). Сколь угодно древние предки млекопитающих никогда не входили в класс рептилий. К сожалению, в начале карьеры мы легко заучиваем неверные термины, а потом неохотно с ними расстаёмся, поэтому ошибочная формулировка «звероподобные рептилии» по-прежнему часто встречается в книгах и документальных фильмах.


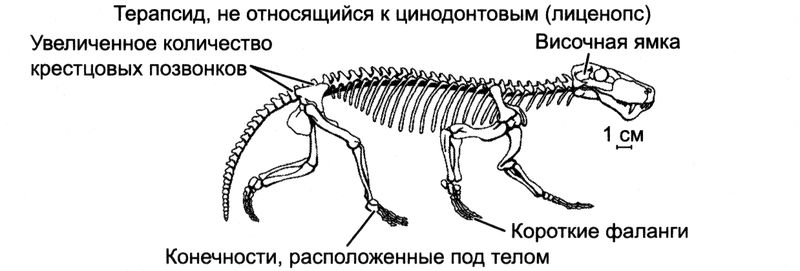
 Рис. 19.1.
Рис. 19.1. Эволюция скелета синапсид от примитивных пеликозавров (в частности, хаптодуса) до терапсид, не относящихся к цинодонтам (например, лиценопса) и цинодонтов (например, тринаксодона), до настоящих млекопитающих, представителем которых является мегазостродон (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 13.4)
Первые хорошо известные синапсиды были найдены в раннепермских красноцветных отложениях северного Техаса — там, где открыли лягомандру и много других важных окаменелостей (см. главу 11). Самые эффектные из этих существ имели спинные гребни, как, например, огромный хищник диметродон (см. рис. 19.1 и 19.2) и травоядный эдафозавр. Хотя эти животные часто присутствуют в детских книгах о динозаврах и среди иной продукции на данную тему, они не имеют с динозаврами ничего общего, а относятся к
нашим предкам! Обычно в подобных товарах, изображающих доисторических животных, «динозаврами» называют не только настоящих динозавров, а и многих других животных, в том числе ихтиозавров и плезиозавров, а также летающих рептилий или птерозавров. Хотя далеко не все доисторические животные являются таковыми. Динозавры имели ряд уникальных анатомических признаков, в том числе отверстие в вертлужной впадине, характерную переднюю конечность всего с тремя функциональными пальцами (большим, указательным и средним), редуцированный безымянный палец и мизинец, сустав в середине голеностопа.

 Рис. 19.2.
Рис. 19.2. Скелеты типичных синапсид: (
А) пеликозавр диметродон с гребнем на спине; (
Б) волкоподобный лиценопс, относящийся к горгонопсам (фотографии предоставлены R. Rothman)
В раннепермском периоде господствующим хищником на территории Техаса был диметродон. Известно множество почти полных скелетов этого животного, а также десятки черепов и частично сохранившихся скелетов (см. рис. 19.2,
А); это одни из самых распространённых окаменелостей в данных отложениях. Крупные особи вырастали от 1,7 до 4,6 м, их спинной гребень («парус») возвышался до 1,7 м над землёй, а вес животного составлял до 250 кг. У диметродона был узкий сжатый череп с длинными кривыми челюстями, усаженными грозным частоколом конических зубов-кинжалов. Зубы отличались по размеру: в передней части челюсти находились большие клыковидные зубы, далее, в направлении к задней части челюсти, они больше походили на простые конусы и уменьшались в размерах. На самом деле именно благодаря этому признаку Эдвард Дринкер Коп в 1878 году назвал животное диметродоном («зубы двух размеров»). Практически единственная черта черепа, роднящая диметродона с млекопитающими, если не считать специализированных зубов, — отверстие (височное окно) в нижней части головы. Нижнее височное окно — один из отличительных признаков синапсид; в изменённом виде оно присутствует у всех млекопитающих и, вероятно, служило точкой прикрепления для сильных челюстных мышц, которые могли вдаваться в это отверстие при жевании. У более поздних синапсид эта черта была очень важной.
Давно ведутся споры о том, для чего был нужен эффектный «парус» на спине диметродона и его травоядного родича эдафозавра, найденного в тех же отложениях. Список возможных толкований велик, но некоторые палеонтологи считают этот орган «радиатором», с помощью которого холоднокровный диметродон мог регулировать температуру тела: повёрнутый перпендикулярно солнечным лучам, «парус» должен был стремительно впитывать тепло, параллельно — остужать хозяина. Однако, поскольку большинство других синапсид того времени не имели терморегулирующего гребня, другие палеонтологи полагают, что он мог использоваться напоказ: позволял распознавать собратьев, а другим животным демонстрировал размеры и силу диметродона. Именно этой цели служат большие рога у современных антилоп и оленей.
Великая Кару
В сердце Южной Африки расположена огромная пустыня Кару. Как и в большинстве пустынь, там наблюдаются резкие перепады температур, а
также стремительно наступают влажные и засушливые периоды. В среднем в этой пустыне выпадает менее 250 мм осадков в год, причём большую часть влаги пустыня получает при немногочисленных штормовых ливнях, случающихся здесь во время короткого влажного сезона. Для южноафриканских колонистов, шедших на север от Кейптауна, пустыня была серьёзной преградой на пути к высокотравным степям Хайвельд в северной части страны. Флора Кару состоит в основном из суккулентов, таких как молочаевые, внешне напоминающие кактусы Нового Света, а также из алоэ, пустынных эфемер и других растений, приспособившихся к чередованию наводнений и засухи, а также к экстремальным перепадам температур. В Кару обитают млекопитающие, адаптировавшиеся к таким условиям: многие антилопы (в частности, спрингбок, символ Южной Африки), гну, страусы, немногочисленные слоны, носороги и гиппопотамы. В своё время здесь обитала и наполовину покрытая полосами лошадь квагга, напоминавшая зебру (истреблена). На неё охотились львы, леопарды, шакалы, гиены и другие хищники. С распространением ирригации эти территории достались овцам и коровам, полностью выедавшим скудную растительность и поставившим немногочисленные популяции диких животных на грань вымирания.
Кару важна и для нашего рассказа об истории жизни. Отложения супегруппы Кару начинаются с конгломерата Двика, относящегося к раннему карбону (310 млн лет назад), где расположены одни из древнейших ледниковых отложений Гондваны; далее идёт мощная последовательность пермских отложений (300–250 млн лет), относящихся к формациям Экка и Бофорт. К этому периоду относится крупнейшее в истории массовое вымирание, случившееся 250 млн лет назад и завершившееся к концу формирования формации Бофорт, в начале триаса (250–200 млн лет назад). Выше этих пермско-триасовых красноцветных отложений находятся другие триасовые породы из формации Стормберг. Наконец, ещё выше залегает отвердевшая юрская лава, излившаяся в эпоху вулканизма в Драконовых горах (около 180 млн лет назад). Формация Бофорт так богата важными позднепермскими и триасовыми окаменелостями, что именно по ней определяется геохронология этих периодов. В частности, именно в свите Бофорт образовались важнейшие окаменелости синапсид и других обитателей поздней перми, демонстрирующие следующий виток эволюции по направлению к млекопитающим. В некоторых местах кости и черепа в огромном количестве пылятся прямо на земле, так что палеонтологам приходится быть разборчивыми и собирать лишь те экземпляры, которые лучше сохранились.
Первооткрывателем этих невероятных окаменелостей стал шотландец Эндрю Геддс Бэйн, обнаруживший их в 1838 году близ Форт-Бофорта в отвалах грунта, сделанных во время прокладки дороги. Он отправил несколько образцов в Британский музей, где их описал Ричард Оуэн, один из основоположников палеонтологии. К концу XIX века в Великобританию поступали все новые окаменелости, и ими заинтересовался другой шотландец, Роберт Брум. Уже в 1897 году он осознал, что эти окаменелости принадлежат не рептилиям, а синапсидам — родичам млекопитающих.
В 1903 году Брум, получивший в Глазго специальность врача и анатома, переехал в Южную Африку, где стал работать врачом, а в свободное время собирал окаменелости. Вскоре он собрал и описал сотни экземпляров позднепермских синапсид, а также удивительных рептилий того же времени и гигантских амфибий. Он стал куратором палеонтологического отдела в Южноафриканском музее Кейптауна, но за эту работу плохо платили, и он едва сводил концы с концами. Его друг Рэймонд Дарт (см. главу 24) написал о постыдной ситуации премьер-министру Яну Смэтсу. Впоследствии, в 1934 году, Брума приняли на работу в Трансваальский музей в Претории. Тогда сфера его интересов изменилась: он стал заниматься преимущественно пещерами времён ледникового периода, расположенными на севере ЮАР, и вскоре прославился своими открытиями первых гоминид, в том числе большинства экземпляров австралопитека африканского и панатропа робустуса. В 1946 году Национальная академия наук наградила его медалью Даниэля Жиро Эллиота, а ещё позднее — Брум дожил до почтенного возраста 84 лет — он был отмечен за основополагающий вклад как в палеонтологию синапсид, так и в палеоантропологию.
Горгоновы морды, ужасные головы и двойные собачьи зубы
В красноцветных позднепермских отложениях были найдены разнообразные синапсиды; эволюцию этой группы удалось проследить на отрезке около 30 млн лет. Давно исчезли архаичные синапсиды со спинным гребнем, в частности диметродон, наиболее известный по раннепермским отложениям из Техаса (см. рис. 19.2,
А). Зато там нашлись более высокоразвитые и зверообразные синапсиды, скопом отнесённые в очередную «мусорную корзину» под названием терапсиды (рис. 19.3). Некоторые из них были первыми сухопутными травоядными животными, среди которых приземистые твари с беззубым клювом и большими клыками, напоминавшими собачьи, — дицинодонты (в переводе с греческого «двойные собачьи зубы»), достигавшие 3,5 м в длину и весившие до тонны. Другие травоядные назывались диноцефалами (ужасноголовыми): их увесистые черепа были усеяны всевозможными буграми, наростами и толстыми костяными «таранами». Некоторые диноцефалы имели до 4,5 м в длину и весили до 2 т.
 Рис. 19.3.
Рис. 19.3. Эволюционная радиация черепов синапсид от простейших пеликозавров, терапсид и цинодонтов до млекопитающих (из работы
Kenneth V. Kardong. Vertebrates: Comparative Anatomy, Function, Evolution. — Dubuque, Iowa: Brown, 1995; с разрешения McGraw-Hill)
На этих травоядных охотились всевозможные свирепые хищные терапсиды: биармозуховые, тероцефаловые, бауриоиды и другие. Самые эффектные из них — устрашающие горгонопсовые (в переводе с греческого «с лицом горгоны») с огромными черепами, снабжёнными впечатляюще мощными клыками и сильными жевательными челюстными мускулами. Горгонопсы отличались крепким телосложением. Крупнейшие из них больше медведя, череп достигал 45 см в длину, большой зуб — более 12 см, а длинное гибкое крокодилоподобное тело — до 3,5 м. В ходе эволюции, на протяжении позднего пермского периода эти терапсиды приобретали всё больше черт, свойственных млекопитающим. Небольшое отверстие в боковой части черепа, имевшееся у диметродона, превратилось в широкий заглазничный костный свод, в который вдавались челюстные мускулы; таким образом, животное не просто обладало мощным укусом, а могло понемногу жевать. Первичное рептильное нёбо стало покрываться вторичным нёбом, нараставшим поверх и закрывавшим носовые пазухи (вы можете пощупать вторичное нёбо, приподняв язык во рту). Благодаря вторичному нёбу, обладавшие им терапсиды могли набрать полный рот пищи и пережёвывать её, продолжая дышать, что необходимо для животного с быстрым метаболизмом. Напротив, типичная рептилия (например, змея или ящерица) должна задержать дыхание, пока не проглотит добычу; но и метаболизм у них медленный.
Вместо одиночного шаровидного сустава в задней части черепа, сразу под тем местом, где позвоночник соединяется с шеей, терапсиды имели двойной шаровидный сустав, благодаря которому увеличивалась сила и гибкость шейных мышц. У терапсид также значительно изменился скелет (см. рис. 19.1), поэтому внешне они больше походили на млекопитающих, чем древние синапсиды. Это касалось и осанки: они не ползали на брюхе, как крокодилы, а удерживали тело в полураспластанном или почти ровном положении.
Ухочелюсти
Самые интересные трансформации у этих животных затронули челюсти и область уха. У примитивных синапсид, например у диметродона, челюсть состояла из основной нижней кости (зубной), а также из ряда других «беззубых» костей, расположенных в задней части черепа: угловой, надугловой, сочленовной, венечной и зачастую многих других (рис. 19.4). Особенно важна в данном случае сочленовная кость, так как она образует челюстной сустав, прикрепляясь к квадратной кости черепа. Но из-за всех этих дополнительных костей и их швов в задней части челюсти аппарат усложнился и стал менее прочным, чем могла бы быть цельная кость. Поэтому, чем активнее терапсиды специализировались в сторону пережёвывания пищи и других сложных челюстных движений, тем сильнее зубная кость распространялась назад и вытесняла незубные кости в заднюю часть черепа. В итоге эти кости уменьшились, а затем были утрачены как нефункциональные.
 Рис. 19.4.
Рис. 19.4. Постепенная трансформация костей челюсти в ходе эволюции синапсид, по мере того как незубные элементы (закрашены тёмным) редуцировались, а зубная кость (не закрашена) росла назад и вытесняла их. В челюсти млекопитающих не осталось никаких незубных элементов, кроме сочленовной кости, которая соединяется с квадратной костью черепа, где переходит в косточки среднего уха (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 13.5)
Сохранилась лишь сочленовная кость, по-прежнему прикреплённая к квадратной кости черепа и служащая челюстным суставом. В конце концов расширившаяся зубная кость вплотную подошла к другой черепной кости, чешуйчатой, и появился новый челюстной сустав. У некоторых синапсид, например у диартрогната (в переводе с греческого «двойной челюстной сустав»),
бок о бок работали челюстные суставы между чешуйчатой и зубной костью, а также между квадратной и сочленовной костью. Поэтому такое животное буквально имело двойные суставы по обе стороны челюсти.
Что произошло затем, когда сустав между зубной и чешуйчатой костью окончательно возобладал? Исчез ли сустав между квадратной и сочленовной костью? Нет. В действительности эволюция блестяще воспользовалась открывшейся возможностью: этот сустав превратился в кости среднего уха! Квадратная кость стала наковальней, сочленовная — молоточком. Как известно, наковальня, молоточек и стремечко передают вибрации барабанной перепонки во внутреннее ухо. Звучит невероятно, но палеонтологическая летопись это подтверждает. Более того, большинство рептилий «слушают» лишь вибрацию грунта, которую могут уловить нижней челюстью, поскольку сустав между квадратной и сочленовной костью играет у них двоякую роль: является и челюстью, и ухом.
Если это по-прежнему кажется невероятным, добавлю, что такое превращение происходило и с вами, и со мной, и со всеми остальными млекопитающими нашего времени. На эмбриональной стадии развития хрящевые зачатки квадратной и сочленовной кости входят в состав зачатков челюстного сустава. По мере развития эмбриона они смещаются в область среднего уха, повторяя эволюционную историю синапсид.
Развитие тринаксодона
В конце пермского периода (около 250 млн лет назад) произошло величайшее массовое вымирание в истории Земли, когда исчезли около 70% сухопутных животных (включая насекомых) и 95% животных в океане. Причины Великого пермского вымирания были сложными, но само событие, видимо, спровоцировали мощные излияния лавы, покрывшей большую часть Восточной Сибири. Вместе с лавой высвободились огромные объёмы парниковых газов (особенно двуокиси углерода), попавшие в атмосферу и океаны. На Земле наступил «суперпарниковый» эффект, а океаны оказались перенасыщены двуокисью углерода, стали очень горячей и кислой средой; в результате там погибло практически всё живое. Доля кислорода в атмосфере сильно снизилась, в то время как атмосфера была перегружена углекислым газом, поэтому вымерли практически все сухопутные животные крупнее определённого размера. Уцелели лишь некоторые эволюционные линии сравнительно мелких синапсид, рептилий, амфибий и других жителей суши, которые перетерпели адское пекло поздней перми и сохранились до наступившего вслед за ней триасового периода.
После того как позднепермские терапсиды почти полностью полегли при массовом вымирании, их уцелевшие представители начали всё сначала. Пошла третья Великая эволюционная радиация новых терапсид, гораздо более похожих на млекопитающих. Их назвали цинодонтами (в переводе с греческого «собакозубые») (см. рис. 19.3). Среди них были животные размером с медведя, например циногнат («собачья челюсть»), достигавший 1–2 м в длину и имевший голову более 60 см; многие более мелкие виды, сравнимые по размеру с енотом и лаской. У большинства цинодонтов была ровная осанка, конечности полностью располагались под телом, что позволяло им быстро бегать (см. рис. 19.1). Челюстные кости, на которых отсутствовали зубы, стали у них крошечными и редуцировались фактически до костных щепок в задней части челюсти, где она крепится к черепу. У цинодонтов имелось сплошное вторичное нёбо до самой глотки, как у современных млекопитающих, многие другие признаки активной жизни и быстрого метаболизма. У многих были многобугорковые коренные зубы, а не простые конические колышки, как у примитивных синапсид. Это подсказывает, что такие животные были способны совершать сложные жевательные движения, а не заглатывать добычу целиком, как поступают рептилии.
Превращение примитивных амниот в млекопитающих проиллюстрировано таким обилием ископаемых переходных форм, относящихся к синапсидам, что невозможно выбрать одну окаменелость в качестве ключевого недостающего звена. Если это всё-таки необходимо, лучший кандидат на эту роль — тринаксодон (рис. 19.5; см. также рис. 19.1). Тринаксодон существовал в самом начале эволюционной радиации синапсид, после раннепермских тварей с гребнями на спине и обитателей средней и поздней перми, найденных в пустыне Кару (см. рис. 19.4). Он был одним из первых цинодонтов и первым животным, обладавшим значительным количеством продвинутых эволюционных признаков, знаменующих последний этап эволюционного превращения синапсид в млекопитающих. Тринаксодон типичен для раннего триаса (250–245 млн лет назад, формация Бофорт в Южной Африке), поэтому в нашем распоряжении есть много почти полных экземпляров, а их анатомия изучена лучше, чем у большинства других синапсид.


 Рис. 19.5.
Рис. 19.5. Тринаксодон жил в раннем триасе. Это был высокоразвитый цинодонт, обладавший многими признаками, характерными для млекопитающих. У него имелась шерсть, диафрагма и развитые зубы, пригодные для жевания: (
А) череп молодой особи с хорошо заметными трёхбугорковыми коренными зубами, по которым животное и получило своё название; (
Б) две особи свернулись и спрятались в норке; (
В) реконструкция внешнего облика (иллюстрации (
А–
Б) предоставлены Roger L. Smith, Южноафриканский изико-музей, Кейптаун; иллюстрация (
В) предоставлена Nobumichi Tamura)
Известны два вида тринаксодонов; представители обоих по размеру и комплекции напоминали ласку, имели длинную узкую мордочку и длинное изящное продолговатое тело с короткими лапками. Обычно они вырастали до 30–50 см в длину. Зубная кость является основным элементом челюсти тринаксодона, а кости, на которых нет зубов, напоминают крошечные щепки, хотя у животного сохранялся рептильный челюстной сустав из квадратной и сочленовной кости (см. рис. 19.4). У тринаксодона было полноценное вторичное нёбо, поэтому он мог одновременно дышать и есть. Глаза у него были большими (чтобы ориентироваться в темноте или в норах), голова — относительно крупная. Как и у потомков, его коренные зубы походили не на простые конические колышки, а имели развитые бугорки; такие зубы можно с полным правом назвать молярами и премолярами. Тринаксодон в переводе с греческого означает «тройнозубый». Действительно, у него в пасти были трёхбугорковые коренные зубы (см. рис. 19.5,
А). Височное окно для мышц, расположенное сбоку и сверху на черепе, необычайно велико и обеспечивало сложные жевательные движения челюсти. Однако, в отличие от большинства млекопитающих, тринаксодон всё ещё имел костную дугу, пролегавшую между височным окном и глазницей.
По обе стороны мордочки тринаксодона заметны крошечные ямки — это значит, у него имелись вибриссы. Если мордочка у тринаксодона была волосистой, можно почти не сомневаться, что и всё его тело покрывала шерсть. Обычно волосы не фоссилизируются, поэтому в данном случае мы, возможно, имеем первые признаки появления волос в эволюционной линии млекопитающих.
Хотя лапки у тринаксодона были короткими, они полностью располагались под телом; в результате животное принимало такую позу, будто наполовину припадало к земле (см. рис. 19.1). У тринаксодона имелись развитые кости плечевого и тазового пояса (особенно подвздошная кость, соединяющая бёдра с позвоночником; к ней крепятся ножные мускулы), почти как у более высокоразвитых цинодонтов и млекопитающих. Таким образом, тринаксодон был очень гибок, мог легко развернуться в ограниченном пространстве и свернуться калачиком (см. рис. 19,
Б). Ещё более показательно, что на грудных рёбрах у тринаксодона есть широкие выросты, из-за которых грудная клетка получалась довольно жёсткой и малоподвижной. Это препятствовало рёберному дыханию, характерному для большинства рептилий (и, по-видимому, для примитивных синапсид). Тринаксодон явно должен был обладать диафрагмой — мышечной стенкой между грудной и брюшной полостью (диафрагма помогает лёгким на вдохе и выдохе; эти мышцы есть у всех млекопитающих). Обобщая все признаки — сложные коренные зубы, вибриссы, диафрагму, — приходим к выводу, что тринаксодон был очень похож на млекопитающее, вероятно, покрыт мехом, имел быстрый метаболизм и был теплокровным.
Кроме того, множество полностью сочленённых экземпляров тринаксодона найдены в местах, очень напоминающих неглубокие норки (см. рис. 19.5,
В). Иногда в логове оказывались застигнутыми две и более особей, а ещё в тех же норках, что и тринаксодоны, попадаются амфибии-брумистеги. Сложно сказать, была ли амфибия добычей цинодонта, два животных вместе заползали в укрытие, пытаясь спастись от внезапного наводнения, либо всё дело в какой-то третьей причине.
Тринаксодон — отличное ископаемое переходное звено, сочетает рептильные признаки большинства примитивных синапсид и более звероподобные черты высокоразвитых цинодонтов. Он исключительно походил на млекопитающее своими небольшими размерами, шерстью, сложными зубами и умением жевать, а также быстрым метаболизмом. Однако кости челюсти и челюстной сустав у него оставались рептильными, это же касается плечевых костей и некоторых других примитивных признаков. Тринаксодон жил в норах, спасаясь от суровых условий раннетриасового мира, сложившихся после пермского вымирания: кислорода в атмосфере было мало, озоновый слой тонкий, атмосфера насыщена углекислым газом. Кроме того, в норах можно было уберечься от более крупных хищников того времени. А если обратить внимание на огромные глаза тринаксодона, напрашивается вывод, что он охотился преимущественно по ночам. Вероятно, добычей ему служили мелкие пресмыкающиеся, насекомые и прочие членистоногие, которые должны были в избытке существовать в мире, где почти не осталось хищников.
Тринаксодон вымер в среднем триасе. К тому времени миром завладели его более развитые родичи-цинодонты, господствовавшие на протяжении триаса. Тем временем начали появляться другие группы животных (в частности, крокодилы и первые динозавры). К концу триаса цинодонты вымерли и появились первые настоящие млекопитающие с челюстным суставом, состоявшим из чешуйчатой и зубной кости, и с настоящими сложными коренными зубами. Это были существа размером с землеройку, обитавшие в мире, принадлежавшем колоссальным динозаврам. В течение следующих 120 млн лет (две трети истории млекопитающих) эти обитатели мезозоя оставались мелкими (размером от землеройки до крысы), у них постепенно развивались сложные зубы и другие признаки. Млекопитающие прятались от динозавров в подлеске и выбирались из укрытий в основном ночью, когда ящеры спали. Затем, 65 млн лет назад, все динозавры, кроме птиц, вымерли, и млекопитающие унаследовали у них планету.
Лучше один раз увидеть!
Во многих крупных музеях можно увидеть диметродона и ряд других синапсид, найденных в раннепермских красноцветных отложениях Техаса. Речь, в частности, об Американском музее естественной истории (Нью-Йорк), Денверском музее природы и науки, Музее естественной истории под открытым небом (Чикаго), Музее сравнительной зоологии при Гарвардском университете (Кембридж, штат Массачусетс), Оклахомском музее естественной истории им. Сэма Ноубла при университете штата Оклахома (город Норман).
Большинство позднепермских и раннетриасовых синапсид экспонируются в музеях ЮАР и России, где их находили. Некоторые из этих окаменелостей есть в Американском музее естественной истории.
Дополнительная литература
Chinsamy-Turan, Anusuya, ed. Forerunners of Mammals: Radiation, Histology, Biology. — Bloomington: Indiana University Press, 2011.
Hopson, James A. Synapsid Evolution and the Radiation of Non-Eutherian Mammals //
Donald R. Prothero and Robert M. Schoch, eds. Major Features of Vertebrate Evolution. — Knoxville, Tenn.: Paleontological Society, 1994. — P. 190–219.
Hotton, Nicholas, III, Paul D. MacLean, Jan J. Roth, and E. Carol Roth, eds. The Ecology and Biology of Mammal-like Reptiles. — Washington, D. C.: Smithsonian Institution Press, 1986.
Kemp, Thomas S. Interrelationships of the Synapsida //
Michael J. Benton, ed. The Phylogeny and Classification of the Tetrapods. Vol. 2. Mammals. — Oxford: Clarendon Press, 1988. — P. 1–22.
Kemp, Thomas S. Mammal-Like Reptiles and the Origin of Mammals. — London: Academic Press, 1982.
Kemp, Thomas S. The Origin and Evolution of Mammals. — Oxford: Oxford University Press, 2005.
Kielan-Jaworowska, Zofia, Richard L. Cifelli, and Xhe-Xi Luo. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. — New York: Columbia University Press, 2004.
King, Gillian. The Dicynodonts: A Study in Palaeobiology. — London: Chapman and Hall, 1990.
McLoughlin, John C. Synapsida: A New Look into the Origin of Mammals. — New York: Viking, 1980.
Peters, David. From the Beginning: The Story of Human Evolution. — New York: Morrow, 1991.
20. Уходящие в воду
Происхождение китов
Амбулоцет
Этих догматиков, способных путём словесной эквилибристики представить чёрное белым, а белое — чёрным, никогда ни в чём не убедишь. Но амбулоцет — то самое животное, которое, по их заявлениям, даже теоретически не могло существовать... Не могу представить себе более подходящего сюжета для популяризации науки, а также более приятную и интеллектуально обоснованную дипломатическую победу над закоснелой креационистской оппозицией.
Стивен Джей Гулд. «Hooking Leviathan by Its Past»
Китовые истории
На протяжении тысяч лет люди изумлялись, глядя на самых удивительных обитателей моря — китов, дельфинов и их родичей. В древних средиземноморских культурах считалось, что если дельфины плывут за кораблём, это к удаче. Сюжет об Ионе и ките — популярный библейский эпизод. Как правило, люди причисляли китообразных к рыбам, в Античности не видели разницы между рыбой, китом и дельфином. Это особенно характерно для биологических трактатов древнегреческого философа Аристотеля, идеи которого прочно укоренились в церковной догматике почти на тысячу лет. Даже сегодня многие люди
всё ещё считают китов и дельфинов рыбами. Представители многих традиционных культур охотятся на китов, воспринимают их как один из даров моря, а не как млекопитающих с большим мозгом, сложными социальными отношениями и целым спектром эмоций, хотя потенциально эти животные могут соревноваться в интеллекте с людьми.
Первым человеком, осознавшим, что киты — не рыбы, был не кто иной, как изобретатель современной биологической систематики, шведский естествоиспытатель Карл фон Линне. Он больше известен под латинизированным именем Карл Линней, поскольку в его времена все учёные писали по-латыни. Опубликовав в 1750-е годы схему классификации животных, он верно отметил, что китообразные дышат лёгкими, а не жабрами; они теплокровны и имеют множество других анатомических признаков, отличающих их от рыб.
Хотя многие современники по-прежнему считали китов рыбами, к XIX веку линнеевские представления были восприняты в широких кругах естествоиспытателей. Вот как писал Оливер Голдсмит в книге «A History of the Earth and Animated Nature» («История Земли и одушевлённой природы») в 1825 году:
Как на суше существуют такие отряды животных, которые, по-видимому, были рождены, чтобы властвовать над остальными, обладая более значительными силами и разнообразными инстинктами, так и в океане возникли рыбы, скроенные по более высокородному образу, нежели остальные, и которые, имея сходство с обычными рыбами, по своим влечениям и телосложению сближаются с четвероногими. Это все китообразные; причём они настолько превосходят собратьев из глубин в своих влечениях и инстинктах, что все современные натуралисты чистосердечно исключают их из числа прочих тварей с плавниками и именуют не рыбами, а великими гигантами океана. По их мысли, говорить, что «рыбаки отправились в Гренландию за китами», было бы столь же неграмотно, как «охотник собрался в Блэкуэлл ставить силки на макрель».
«Великий морской змей»
В начале XIX века были открыты некоторые хорошо сохранившиеся окаменелости китов, но, к сожалению, эти находки попали в руки нечистоплотных дельцов, а не в лаборатории квалифицированных учёных. Самый известный из аферистов — «доктор» Альберт Кох. Этот мошенник был даже более беспринципен, чем Барнум, и всегда старался нажиться на диковинных россказнях о естественнонаучных находках. Наконец, ему повезло, и он обзавёлся гигантским скелетом, который назвал гидрархосом («великим морским змеем») (рис. 20.1). Скелет достигал 35 м в длину и помещался в трёх комнатах, а спереди к нему были приделаны большущие ласты. Череп существа заканчивался длинной мордой с огромными треугольными зубами. Скелет привлекал толпы охочих зевак. Однако Кох, будучи умелым организатором зрелищ, не годился в учёные. Он накупил у фермеров в штатах Алабама, Миссисипи и Арканзас ископаемые позвонки, принадлежавшие нескольким особям примитивных китов (так называемых археоцетов), которые встречаются в среднеэоценовых породах (возраст 50–37 млн лет). Этих костей было так много, что в некоторых районах Алабамы фермеры делали из них изгороди. Затем Кох соорудил составной скелет, на который пошли кости минимум трёх китов (чтобы скелет получился подлиннее и покрупнее). Это был излюбленный приём Коха. Ранее он сумел значительно увеличить попавший к нему скелет мастодонта, совместив кости от разных экземпляров и назвав получившееся существо Великим Миссурием.
 Рис. 20.1.
Рис. 20.1. «Гидрархос» Альберта Коха, с которым он объездил Америку и Европу в 1840-е годы. На самом деле этот «змей» собран из скелетов минимум трёх китов-археоцетов, чтобы придать ему вес (рисунок из фонда Wikimedia Commons)
Затем Кох отправился с «великим змеем» в тур по Европе, где путешествовал из города в город и повсюду привлекал огромные толпы желающих посмотреть на «библейского Левиафана». Убравшись из Лондона и Берлина, где учёные подсказывали журналистам, что это надувательство, Кох с «гидрархосом» побывал в Дрездене, Бреслау, Праге и Вене. Король Пруссии Фридрих Вильгельм IV был настолько впечатлён, что в 1847 году пожаловал Коху ежегодное содержание в размере 1000 имперских талеров. Хотя прусские учёные раскритиковали скелет и назвали подделкой, убедить стареющего монарха им не удалось. Гидеон Мантелл (первооткрыватель игуанодона, первого динозавра, получившего название) раскрыл обман и предупреждал людей, что те имеют дело с отъявленным мошенником. В Нью-Йорке анатом Джеффри Уаймен подтвердил, что «Великий морской змей» — никакая не рептилия, а кости взяты от нескольких животных. Коху оставалось ездить по глубокой провинции, где не слышали настоящих учёных. В конце концов он продал своего монстра в Музей полковника Вуда в Чикаго. Там скелет оставался до 1871 года, пока не сгорел во время Великого чикагского пожара.
Несмотря на историю с липовым коховским скелетом, другие ископаемые останки китов попадали в руки истинных натуралистов. В 1834 году анатом Ричард Харлан назвал одну из таких находок базилозавром («царственным ящером»), поскольку кости существа были очень велики. Харлан считал, что это останки одной из гигантских рептилий, которых сегодня называют динозаврами, открытых немногим ранее. Однако в 1839 году великий британский анатом Ричард Оуэн, придумавший слово «динозавр» и описавший некоторые из первых обнаруженных окаменелостей таких ящеров, увидел образцы костей базилозавра и пришёл к выводу, что они принадлежали не динозаврам или другим рептилиям, а огромным китам. Он попытался переименовать существо в зеуглодона (хомутозубого), чтобы заменить явно ошибочное «базилозавр», но было поздно. По правилам именования животных, валидным признаётся название, которое было дано раньше, независимо от степени его удачности. Поэтому кит остался базилозавром, хотя он и не рептилия, а млекопитающее.
По мере того как находились более полные экземпляры, киты-археоцеты привлекали всё больше внимания (рис. 20.2). Даже не будучи монстрами вроде искусственно увеличенного змея Коха, крупные археоцеты могли достигать 24 м в длину и весили около 5400 кг. Они мало чем напоминали современных китов: имели длинную заострённую морду с треугольными зубами, которыми удобно хватать рыбу, однако были значительно примитивнее. Прежде всего, у них не было дыхала на макушке, которое есть у всех современных китов, зато имелись ноздри на кончике носа, как у большинства других млекопитающих. Уши археоцетов тоже были очень примитивными, без специализированных косточек для подводной эхолокации, подобных тем, что есть у современных китов.
 Рис. 20.2.
Рис. 20.2. Собранный скелет базилозавра (публикуется с разрешения Национального музея естественной истории Смитсоновского института)
Передние конечности археоцетов превратились в ласты, но ни в одной из найденных в США фрагментарных окаменелостей задние конечности не сохранились. В 1990 году полностью сочленённые скелеты археоцетов (с задними конечностями) обнаружили в Египте. У кита длиной более 24 м задние лапы оказались не длиннее человеческой руки; они уже явно не функционировали, хотя к ним по-прежнему крепились мышцы в задней части тела. Поскольку киты не использовали их для ходьбы, эти рудименты напоминали о временах, когда киты были четвероногими. Если вы увидите в музее скелет современного кита, обратите внимание на его тазовый пояс чуть ниже позвоночника и последних рёбер грудной клетки. Если экземпляр полный и правильно собран, вы заметите нефункциональные останки бедренных и берцовых костей, спрятанные глубоко в теле животного и абсолютно бесполезные — они просто доказывают, что киты произошли от четвероногих сухопутных животных. Но каких?
Эволюция и киты
Когда в 1859 году опубликовали дарвиновское «Происхождение видов», тот факт, что киты являются млекопитающими, приобрёл ещё одну важную грань: очевидно, киты произошли от наземных животных, которые должны были вернуться в воду. В первом издании своей книги Дарвин размышлял, как могла произойти такая метаморфоза. В качестве примера он приводил историю о чёрных медведях, которые плавают с открытым ртом, хватая рыбу и прочую водоплавающую дичь. Дарвин писал: «Я не вижу трудности в образовании под действием естественного отбора породы медведей, более водных по строению и привычкам и с более крупной пастью, вплоть до появления существа столь уродливого, как кит». К сожалению, критики холодно восприняли эту идею, и в следующих изданиях Дарвин от неё отказался.
Вопрос о происхождении китов находился в подвешенном состоянии более века. Хотя во многих коллекциях имелись многочисленные окаменелости крупных археоцетов, почти отсутствовали хорошие окаменелости более примитивных китов, которые были бы полуводными либо полностью сухопутными животными и при этом обладали бы китовыми признаками. В 1966 году Ли ван Вален, палеонтолог из Чикагского университета, вновь поднял этот вопрос, остававшийся без внимания не одно десятилетие. Он обратил внимание на то, какими огромными притупленными зубами, похожими на треугольные лезвия, усажены челюсти археоцетов, и указал, что именно такие зубы были у группы больших хищных копытных млекопитающих, именуемых мезонихиями. Хотя у мезонихий имелись копыта, это были хищные или всеядные животные, напоминавшие помесь волка и медведя. У многих мезонихий огромные вытянутые морды, примерно как у археоцетов, а вскоре у них стали замечать и другие китовые признаки. Идея о том, что мезонихии могли быть предками китов, распространялась всё шире. Это мнение получило широкое распространение на тот момент, когда мы с Робертом Шохом написали книгу о копытных млекопитающих.
Серьёзные поиски более примитивных ископаемых китообразных начались только в 1970-е и 1980-е годы. В то время Пакистан был должен США много миллионов долларов за военное оборудование, которое приобрёл по оборонным контрактам. Пакистанцы стремились погасить долг, поэтому США без особого труда получили гранты на палеонтологические исследования в их стране. Кроме того, палеонтологи знали, что важные окаменелости древних китов (в основном археоцетов) впервые были найдены в северо-западной части Индии (сегодня это территория Пакистана) Гаем Пилгримом в 1920-х, затем Ашоком Сахни и другими в начале 1970-х. Поэтому ряд палеонтологов, в частности Филип Джинджерич из Мичиганского университета и Ханс Тевиссен из медицинского колледжа Северо-восточного университета штата Огайо, взялись за исследование пакистанских пород, чей возраст старше отложений с археоцетами и которые представляли собой осадочные слои, сформировавшиеся по берегам морей или на морском мелководье.
Благодаря очень кстати полученному щедрому финансированию пакистанских исследований палеонтологи смогли найти тот самый отрезок во времени и пространстве, на котором киты произошли от сухопутных животных. Это случилось в раннем эоцене (55–48 млн лет назад) в мелкой тропической акватории под названием Тетис. Она напоминала о днях, когда на Земле существовал суперконтинент Пангея и суперокеан Панталасса. Тетис простирался от Западного Средиземноморья до Индонезии; акватория распалась, когда Африка сдвинулась на север и замкнула Средиземное море, а Индия в среднем эоцене врезалась в подбрюшье Азии и рассекла оставшуюся часть Тетиса пополам. Однако прежде чем Тетис исчез, на его берегах появились не только первые киты, вернувшиеся в воду, но и древнейшие родичи ламантинов (см. главу 21) и многие другие своеобразные животные: мастодонты, мартышки и обезьяны, даманы.
Первым важным ископаемым переходным звеном в эволюции китов был пакицет, об обнаружении которого Джинджерич и его коллеги сообщили в 1983 году (рис. 20.3). Хотя скелет пакицета в целом похож на волчий (животное имело четыре длинные конечности для ходьбы), его череп напоминает черепа археоцетов. В частности, это касается больших зазубренных треугольных зубов. Мозг пакицета был маленьким и примитивным, ухо не имело никаких специальных черт, приспособленных для слушания под водой или эхолокации. Однако ушные кости плотные; другие признаки тоже свидетельствуют о его возможности слышать под водой. Пакицета нашли в речных отложениях возрастом около 50 млн лет, то есть он был преимущественно сухопутным животным, но проводил много времени в воде. Хотя длинные ноги животного с короткими кистями и ступнями приспособлены преимущественно для бега и прыжков, кости конечностей необычно толстые и могли служить балластом в воде, если предположить, что оно больше ходило по мелководью, чем плавало.
 Рис. 20.3.
Рис. 20.3. Происхождение китов от сухопутных млекопитающих с реконструкцией многочисленных переходных форм, найденных в эоценовых отложениях на территории Африки и Пакистана (рисунок Карла Бьюэлла из работы
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 14.16)
«Ходячий плавающий кит»
Крупнейший прорыв произошёл в 1994 году, когда Ханс Тевиссен объявил об открытии
Ambulocetus natans, название которого буквально означает «ходячий плавающий кит» (рис. 20.4). Речь о практически полном скелете, добытом в формации Верхняя Кулдана на территории Пакистана (прибрежные морские отложения возрастом около 47 млн лет). Это животное действительно наполовину похоже на кита, а наполовину — на сухопутное млекопитающее. Амбулоцет имел примерно 3 м в длину (сегодня таких размеров достигают крупные морские львы). У него была длинная зубастая морда, примерно как у других примитивных китов, с характерными треугольными зубами. Его уши по-прежнему были не слишком специализированными и явно не подходили для эхолокации, но с их помощью амбулоцет, вероятно, мог улавливать вибрацию в грунте или воде. Его длинные сильные лапы заканчивались очень длинными пальцами, между которыми, вероятно, имелись перепонки. Это был четвероногий кит, умевший как ходить, так и плавать — отсюда и название.


 Рис. 20.4.
Рис. 20.4. Амбулоцет — «ходящий плавающий кит»: (
А) наиболее полный скелет и его первооткрыватель Ханс Тевиссен; (
Б) слепок скелета в «идущей» позе; (
В) реконструкция внешнего вида плывущего амбулоцета ((
А) иллюстрация предоставлена H. Thewissen, NEOMED; (
Б) фотография автора; (
В) иллюстрация предоставлена Nobumichi Tamura)
Исследования позвоночника показали, что амбулоцет мог выгибать спину вверх и вниз подобно выдре, а не перебирать ластами в воде, как тюлень или пингвин. Такое изгибание очень похоже на движения некоторых китов, хотя торс у большинства из них негибкий и для движения они используют лишь хвост.
Амбулоцет явно не был быстрым пловцом. Тевиссен полагает, что его «крокодилья» комплекция скорее свидетельствует о повадках хищника-засадника, терпеливо выжидающего под водой, когда добыча подойдёт ближе, а затем бросающегося на неё. Тот факт, что эти особи были найдены в прибрежных морских породах формации Верхняя Кулдана, свидетельствует: амбулоцет жил по берегам озёр, рек и океанов. Химический анализ зубов дополнительно подтверждает, что амбулоцет обитал как в солёной, так и в пресной воде.
Через несколько лет после открытия амбулоцета обнаружили почти полный скелет другого кита, так называемого даланиста (см. рис. 20.3). Как и амбулоцет, даланист имел полностью функциональные передние и задние конечности; пальцы у него были ещё более длинными, и между ними натягивались перепонки. При этом морда ещё длиннее и «китовее», чем у амбулоцета, равно как сильный хвост.
В 1994 году, когда Тевиссен объявил об амбулоцете, Филип Джинджерич и его коллеги нашли другого, более продвинутого предка китов. Это ископаемое обнаружили в Южном Белуджистане на территории Пакистана (см. рис. 20.3). Животное, названное родоцетом, — самый известный представитель семейства примитивных китообразных протоцетид, по размеру сравнимых с дельфином (правда, один из них, гавиацет, достигал более пяти метров в длину). Череп родоцета гораздо крупнее, чем у амбулоцета, и больше похож на китовый; морда длинная, зубы типичные археоцетовые. Шейные позвонки свидетельствуют о том, что его туловище и голова слиты в единое обтекаемое тело, причём отсутствовала выраженная шея, которую родоцет мог бы поворачивать отдельно от торса. Кости конечностей у него были длинными, но короче, чем у амбулоцета и даланиста; пальцы тоже короче. То есть животное имело более короткие задние лапы с перепончатыми ступнями (ещё не полноценные китовые плавники). Однако тазовые кости и тазовые позвонки у него по-прежнему составляли единое целое — значит, животное могло ходить по земле. Пропорции скелета свидетельствуют, что родоцет плавал, перебирая лапами, а хвост использовал преимущественно в качестве весла.
После открытия родоцета обнаружили много других переходных форм, в частности такрацета и гавиоцета. Они обладали более специализированными кистями, постепенно превратившимися в китовые плавники (см. рис. 20.3), а задние лапы у них постепенно уменьшались. Их тела тоже всё больше напоминали дельфиньи, животные активно использовали для движения хвост (как современные киты). Это значит, что у них, вероятно, были горизонтально расположенные хвостовые лопасти.
Сегодня известно так много ископаемых переходных китообразных, что невозможно провести границу между сухопутными животными и настоящими китами. Происхождение китов, которое ещё в 1980-е годы оставалось тайной, сегодня является одним из самых подробно «документированных» эволюционных переходов в палеонтологической летописи.
Кратко о бегемотоподобных
Большинство палеонтологов считали волкоподобных копытных мезонихий вероятными предками китов — с тех самых пор, как эту гипотезу высказал в 1966 году Ли Ван Вален. Сходство их зубов и черепов поражает, причём ни у одной другой группы млекопитающих на планете нет таких своеобразных зубов. Когда Эрл Мэннинг, Мартин Фишер и я в 1988 году опубликовали анализ взаимосвязей между копытными, анатомические признаки, казалось, свидетельствовали в пользу родства между китами и мезонихиями. Кроме того, обе эти группы находились в близком родстве с парнокопытными животными (
Artiodactyla) — свиньями, бегемотами, верблюдами, жирафами, оленями, антилопами, быками и баранами.
Но в конце 1990-х молекулярные биологи начали секвенировать ДНК и некоторые белки, входящие в состав важных молекул у многих групп млекопитающих. Появлялись новые доказательства того, что китообразные не просто ближе к парнокопытным, чем любые современные животные, но и
произошли от них. Если говорить о современных парнокопытных, ближайшими родичами китов определённо являются бегемоты (см. рис. 20.3). Палеонтологи не спешили соглашаться с выводами молекулярной биологии, поскольку анатомические доказательства, полученные при изучении ископаемых мезонихий, казались гораздо убедительнее, а древнейшие киты и первые бегемоты совсем не походили друг на друга. Тем более что молекулярный анализ делали на биоматериале современных животных: у нас нет ДНК и белков ни одной из многочисленных окаменелостей, которые, очевидно, связывают китов и мезонихий.
Но палеонтологическая летопись вновь нас удивила и помогла разрешить эту проблему. В 2001 году две независимые группы исследователей под руководством Тевиссена и Джинджерича сообщили, что в Пакистане найдены окаменелости древних китов с хорошо сохранившимся голеностопом. Для голеностопа этих животных была характерна двухжелобковая конфигурация таранной кости (у млекопитающих является частью лодыжки), ранее известная
только у парнокопытных. Все парнокопытные, в отличие от других групп млекопитающих, имеют такую двухжелобковую таранную кость. Более того, многие парнокопытные могут быть причислены к этому отряду лишь по признаку наличия такой уникальной кости. Теперь, на основании изучения находок из Пакистана, ясно, что уникальная анатомия голеностопа характерна не только для парнокопытных, но и для китообразных.
Неприятие идеи, что киты — потомки парнокопытных, быстро сошло на нет, палеонтологи вернулись к своим кульманам и занялись анализом, учитывая новые анатомические находки и факты молекулярной биологии. Вскоре сошлись на том, что киты произошли от парнокопытных и должны классифицироваться как одна из групп в той эволюционной ветви, которая привела к появлению бегемотов. Вместо двух разных отрядов, «китообразные» и «парнокопытные», новые данные и достигнутое соглашение требуют переименовать общий отряд в китопарнокопытные и считать китов подотрядом в составе одной из эволюционных линий парнокопытных, куда относятся бегемоты и родственные им антракотерии. Однако это нарушает принципы таксономии. Когда одна группа входит в состав другой, название более крупной группы обычно не меняется. Следовательно, теперь, когда парнокопытные включают китообразных, переименовывать весь отряд в китопарнокопытные не требуется. Мы же не переименовали динозавров в птицезавров, чтобы объединить их с птицами. Молекулярные биологи называют бегемотоподобными группу, в которую входят киты и гиппопотамы, хотя большинство учёных предпочитают именовать её цетакодонтоморфами.
Теперь вместо привычной картинки, где китообразные представляют собой отряд, расположенный рядом с парнокопытными, они оказались внутри последних и отнесены к группе, близкородственной бегемотам и другим примитивным ископаемым парнокопытным. Мезонихии стали изгоями, теперь их принято считать ближайшими родичами китопарнокопытных. Такая классификация означает, что их характерные треугольные зубы должны были развиваться параллельно с аналогичными зубами археоцет, но легче согласиться с таким эпизодом конвергентной эволюции, чем списать на неё многочисленные примеры молекулярного сходства между китообразными и бегемотами.
Впрочем, как знать? Если бы мезонихии существовали сегодня и мы могли секвенировать их ДНК, возможно, ответ был бы другим. Они вымерли к концу эоцена, более 33 млн лет назад, поэтому правду мы никогда не узнаем.
Род гиппопотамов
Представить китообразных и гиппопотамов близкими родичами несложно. В конце концов, и те и другие — крупные водные животные. Опять же, нас выручает палеонтологическая летопись. Ископаемая история современного семейства бегемотовых прослеживается всего на 8 млн лет назад. Но бегемотов можно связать с вымершим семейством парнокопытных, антракотериями, история которых прослеживается до 50 млн лет назад. Антракотерии отличались друг от друга комплекцией и тонкостями адаптации, но большинство из них, видимо, вели водный или полуводный образ жизни.
Недавно в кашмирских породах обнаружили замечательную переходную окаменелость, связывающую две группы. Это животное, известное под названием индохиус (буквально «индийская свинья»), в 2007 году описал Ханс Тевиссен — на материале окаменелостей, собранных ранее индийским геологом А. Ранга Рао (рис. 20.5). Хотя это животное было лишь чуть крупнее кролика и прыгало на длинных лапках, а пропорциями тела напоминало небольшого оленя, характерные анатомические признаки показывают, что перед нами — переходное звено между китами и другими парнокопытными. В области уха у индохиуса находим многие черты, встречающиеся лишь у китов. Кости конечностей у него были очень плотными (как у гиппопотамов, китов и других водных животных), служили балластом, помогали животному бродить по воде или подныривать так, что оно не рисковало неожиданно всплыть. По данным химического анализа костей, индохиус был водным животным, но зубы у него такие, будто он питался наземными растениями. Индохиус — последнее недостающее звено, объединяющее самых примитивных китообразных наподобие пакицета, который тоже был преимущественно сухопутным животным, с антракоцетами и, соответственно, завершающим китово-бегемотовую эволюционную линию.

 Рис. 20.5.
Рис. 20.5. Индохиус, древнейший общий предок эволюционной линии китов и гиппопотамов: (
А) наиболее полный скелет; (
Б) реконструкция внешнего вида ((
А) иллюстрация предоставлена H. Thewissen, NEOMED; (
Б) иллюстрация предоставлена Nobumichi Tamura)
Таким образом, киты — совсем не рыбы и не потомки плавающих медведей. По данным молекулярного анализа, они произошли от общего предка, объединяющего их с антракотериями и бегемотами. При наличии таких окаменелостей, как пакицет, даланист, амбулоцет, родоцет и индохиус, по ископаемой летописи легко проследить, как киты произошли от сухопутных животных.
Лучше один раз увидеть!
Окаменелости амбулоцета, дорудона и других переходных представителей китообразных остаются в тех странах, где были найдены. Однако в некоторых музеях есть слепки этих переходных окаменелостей, а также полные скелеты базилозавра. В США следует назвать Музей естественной истории штата Алабама (город Таскалуса), Американский музей естественной истории (Нью-Йорк), Музей естественной истории под открытым небом (Чикаго), Палеонтологический музей при Мичиганском университете (Анн-Арбор), Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия) и Музей естественной истории округа Лос-Анджелес (Лос-Анджелес). В Европе такие образцы экспонируются в Центре биоразнообразия Naturalis в Лейдене (Нидерланды), а также в Музее естественной истории Зенкенберг (Франкфурт-на-Майне, Германия). Несколько сложнее добраться до Новозеландского музея Те Папа Тонгарева в Веллингтоне и до Национального музея природы и науки (Токио, Япония).
Дополнительная литература
Berta, Annalisa, and James L. Sumich. Return to the Sea: The Life and Evolutionary Times of Marine Mammals. — Berkeley: University of California Press, 2012.
Berta, Annalisa, James L. Sumich, and Kit M. Kovacs. Marine Mammals: Evolutionary Biology. 2nd ed. — San Diego: Academic Press, 2005.
Janis, Christine M., Gregg F. Gunnell, and Mark D. Uhen, eds. Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals. — Cambridge: Cambridge University Press, 2008.
Prothero, Donald R., and Robert M. Schoch. Horns, Tusks, and Flippers: The Evolution of Hoofed Mammals and Their Relatives. — Baltimore: Johns Hopkins University Press, 2002.
Rose, Kenneth D. The Beginning of the Age of Mammals. — Baltimore: Johns Hopkins University Press, 2006.
Rose, Kenneth D., and J. David Archibald, eds. The Rise of Placental Mammals: The Origin and Relationships of the Major Extant Clades. — Baltimore: Johns Hopkins University Press, 2005.
Thewissen, J. G. M., ed. The Emergence of Whales: Evolutionary Patterns in the Origin of the Cetacea. — Berlin: Springer, 2005.
Thewissen, J. G. M., ed. The Walking Whales: From Land to Water in Eight Million Years. — Berkeley: University of California Press, 2014.
Zimmer, Carl. At the Water’s Edge: Fish with Fingers, Whales with Legs, and How Life Came Ashore but Then Went Back to Sea. — New York: Atria Books, 1999.
21. Ходячие ламантины
Происхождение сиреновых
Пезосирен
До пояса она напоминала человека, но ниже походила на рыбу с широким хвостом в форме полумесяца. Лицо её было округлым и мясистым, нос — толстым и плоским; чёрные волосы вперемешку с серым мехом спускались по плечам и прикрывали живот. Высунувшись из воды, она откинула волосы руками с лица, а когда вновь нырнула, фыркнула, как пудель. Мой товарищ забросил удочку, посмотреть, клюнет ли она. Но она ушла на глубину и больше не появилась.
Герберт Вендт. «Auf Noahs Spuren»
Русалки!
Мифы о русалках существуют многие тысячелетия, встречаются в сказках о море во многих культурах. Самая древняя из известных историй такого рода появилась в Ассирии около 2300 года до н. э. Она рассказывает о богине Атаргатис, которая превратилась в русалку, раскаиваясь за случайное убийство своего возлюбленного-пастуха. В «Одиссее», приписываемой Гомеру и датируемой примерно VIII веком до н. э., упоминаются сирены — мифические женщины-рыбы, зачаровывающие моряков своим пением так, что их корабли разбиваются о рифы. Таинственные «морские девы» упоминаются в нескольких сказках Шехеразады из сборника «Тысяча и одна ночь». Рассказы о встречах с русалками и русалами в последние 2000 лет были распространены почти во всех европейских странах. Герберт Вендт приводит одно из таких сообщений, полученное от двух французских моряков, якобы повстречавших русалку у острова Мартиника в 1671 году. Эти легенды канонизировались в популярных сказках, таких как «Русалочка» (1836) Ханса Кристиана Андерсена, по которой в 1989 году был снят знаменитый диснеевский мультфильм. В фильме «Всплеск» (1984), где Дэрил Ханна сыграла настоящую русалку, вновь пересказывается этот миф. Совсем недавно, в 2012 и 2013 годах, на кабельном канале Animal Planet вышли два «документальных» фильма о том, что русалки якобы реально существуют и даже были найдены. Множество людей поверили в эти липовые «факты». Псевдодокументалистика воспринимается настолько серьёзно, что учёным из Национального управления океанических и атмосферных исследований дважды пришлось тратить драгоценное время и публиковать на сайте агентства специальные пресс-релизы о том, что такие фильмы — чистая фикция и русалок не существует.
Некоторые из легенд — лишь плод бурной человеческой фантазии, их можно сравнить с другими сюжетами о полулюдях-полуживотных, например о кентавре (человек-лошадь) или о минотавре (человек-бык). Но многие учёные считают, что байки моряков основаны на встречах с реальными обитателями моря, которых молва превратила в сказочных русалок. В 1493 году, совершая второе путешествие в районе Гаити, Колумб сообщил о встрече с тремя «женоподобными существами», которые «высоко высовывались из воды, но были не столь красивы, как о них рассказывают». Знаменитый английский пират Чёрная Борода (Эдвард Тич) заявлял, что видел русалок в Карибском море и впоследствии старался держаться подальше от вод, где их замечали. Моряки и пираты верили, что русалки могут зачаровать, польстившись на золото, а затем утащить на морское дно.
Как и рассказы о морских змеях, отдельные «свидетельства очевидцев» о встречах с русалками поступают со всего мира, от Канады до Израиля и Зимбабве. Моряки утверждали, что в Индийском океане русалки плавают парами, причём если одну загарпунить, вторая пытается её спасти. Говорят, они плачут «настоящими слезами», а мать-русалка может баюкать русалёнка на руках.
Наука о русалках
Есть ли реальные факты в основе этих легенд? Некоторые зоологи усматривают в них связь с ламантинами, встречающимися, как правило, в мелких тропических водах Западного полушария, и дюгонями, обитающими преимущественно в Индийском океане, а также с их родичами из отряда морских млекопитающих, известных под названием морских коров или сиреновых (в честь мифических сирен). Дюгонь и ламантин умеют плавать в вертикальном положении, держа голову над водой, когда хотят рассмотреть корабль или другой предмет на поверхности воды (рис. 21.1). У всех сиреновых на груди есть хорошо заметные молочные железы, которые могут напоминать женскую грудь, причём самки кормят сосунков примерно в такой же позе, что и женщины. Но как можно принять столь уродливое создание за прекрасную деву? Если ламантин всплывёт из воды и его морда будет опутана длинными водорослями, издалека они вполне могут напоминать волосы (особенно в бликах, играющих на воде в открытом море). В таком случае несложно ошибочно вообразить, что это водяная женщина (тем более моряку, много времени находящемуся вдали от земли и женщин). Нескольких таких «свидетельств», оставленных Колумбом и другими первооткрывателями, достаточно для подтверждения мифа, тысячелетиями существовавшего почти во всех культурах.
 Рис. 21.1.
Рис. 21.1. Когда сиреновые (например, этот ламантин) всплывают в вертикальном положении, легко понять, как моряки, видевшие животных издали, могли принять их за русалок (рисунок из фонда Wikimedia Commons)
Когда удалось отловить ламантинов и дюгоней и показать их первым естествоиспытателям, возник ужасный конфуз. Осмотр показал, что эти существа ничуть не похожи на волшебных русалок. Первые натуралисты, изучавшие их анатомию, классифицировали морских коров как китообразных, поскольку это полностью водные животные, не имеющие ног, но их передние конечности представляют собой ласты, а на хвосте — две лопасти. Карл Линней заметил множество анатомических особенностей, сближающих их со слонами, а также первым причислил их к отряду хоботных, к которому также относятся слоны, мамонты и мастодонты. В 1816 году зоолог Анри де Бленвиль поддержал линнеевскую интерпретацию, хотя большинство естествоиспытателей по-прежнему относили ламантинов и дюгоней к китообразным.
Но постепенно выявлялись всё новые близкие анатомические черты, сходство между хоботными и сиреновыми становилось всё явственнее. Оба отряда имели ряд уникальных специализированных признаков. В конце концов зоологи отказались от китовой интерпретации. Дискуссия о родстве сиреновых и хоботных достигла апогея в 1975 году, когда Малколм Маккенна предложил объединить их в общий отряд, который назвал тетитериями. Группа получила такое название, поскольку окаменелости из обеих эволюционных линий демонстрируют: и те и другие появились где-то на берегах акватории Тетис, простиравшейся от Средиземноморья через Ближний Восток, мимо Индии до Австралии. Через несколько лет Маккенна, Дэрил Домнинг и Клейтон Рэй описали окаменелость под названием бегемотопс, найденную в олигоценовых породах на северном берегу полуострова Олимпия в штате Вашингтон. Эта находка подтверждала тетитериевое происхождение сиреновых. С тех пор идея о тетитериевых подкреплена многочисленными данными молекулярного анализа, демонстрирующими близкое родство между сиреновыми и хоботными и подтверждающими их многочисленные анатомические сходства, подмеченные ещё Линнеем.
Морские коровы уходят в море
Существуют убедительнейшие анатомические и молекулярные доказательства в пользу того, что сиреновые произошли от общего с хоботными предка около 50 млн лет назад. Что показывает палеонтологическая летопись? Первым изученным ископаемым сиреновых был очень примитивный представитель этого отряда. В 1855 году легендарный британский анатом Ричард Оуэн описал странный череп, присланный в Лондон с Ямайки. Его нашли в местечке Фрименс Холл, в формации Чейпелтон (возраст — 50–47 млн лет). Оуэн, который придумал термин «динозавры», описывал окаменелости, доставленные из Южной Америки на корабле Дарвина «Бигль», и в конце концов стал основным научным оппонентом Дарвина. Хотя череп примитивен, некоторые его куски отломаны, а зубы сточены до корней (рис. 21.2), Оуэн верно определил, что в нём есть слегка опущенные книзу кости рта, расположенное высоко на черепе носовое отверстие, а также заметил многие другие признаки, свойственные сиреновым. От остального скелета сохранились лишь фрагменты, но они позволяли предположить, что окаменелость принадлежит четвероногому животному размером примерно с овцу. Череп и фрагменты костей были найдены вместе с обломками очень крепких и плотных рёбер — типичный признак сиреновых. Рёбра служат балластом и нужны животному, чтобы не всплывать. Очень плотная кость, пусть даже одного обломка ребра, свидетельствует, что речь идёт о морской корове. Оуэн назвал ископаемого обладателя этого черепа
Prorastomus sirenoides (родовое название означает «широкие передние челюсти», видовое — сиреновый). Оуэн уверенно определил, что перед ним окаменелость очень примитивного животного, родственного современным морским коровам. Хотя учёный был одним из последних серьёзных зоологов, отрицавших естественный отбор, он не смог игнорировать сходство окаменелости с современными сиреновыми.

 Рис. 21.2.
Рис. 21.2. Прорастомус: (
А) череп, описанный сэром Ричардом Оуэном; (
Б) реконструкция внешнего вида. ((
А) иллюстрация предоставлена Daryl Domning; (
Б) иллюстрация предоставлена Nobumichi Tamura)
Шли годы, новые окаменелости обнаруживали на берегах Атлантического и Тихого океанов, а также в других местах, где океаны некогда затапливали сушу. В 1904 году австрийский палеонтолог Отенио Абель описал череп более высокоразвитого представителя сиреновых,
Protosiren fraasi, найденного в нижней пачке Билдинг-Стоун формации Джебель-Мокаттам в Египте, датируемой средним эоценом (47–40 млн лет назад) (рис. 21.3). Здешними известняками украшали плиты великих египетских пирамид. Череп больше напоминал современных сиреновых: морда была сильнее загнута книзу, специализированное носовое отверстие отнесено ближе к затылку, имелись и другие продвинутые черты. Впоследствии другие экземпляры этого животного попадались по всему миру: в Северной Каролине, во Франции и Венгрии, в Пакистане и Индии. Таким образом, протосирен обитал в тёплых тропических и субтропических водах почти повсеместно. Когда были найдены останки скелета, выяснилось, что задние конечности у протосирена миниатюрные. Кроме того, тазовые кости не слишком прочно прикреплялись к нижней части позвоночника, поэтому животное было практически полностью водным и едва ли могло ходить по суше. У большинства ископаемых сиреновых моложе протосирена задние конечности редуцированы ещё сильнее — эти животные вели полностью водный образ жизни и не могли ходить. У современных ламантинов и дюгоней есть крошечные рудименты тазовых и бедренных костей, расположенные глубоко в мышцах нижней части спины и совершенно нефункциональные; они лишь доказывают, что эти животные произошли от четвероногих сухопутных предков.
 Рис. 21.3.
Рис. 21.3. Череп протосирена — более высокоразвитого представителя сиреневых, чем прорастомус (иллюстрация предоставлена Daryl Domning)
Итак, у древнейшего ископаемого предка сиреновых (прорастомуса) начинают развиваться характерные черты черепа, а также тяжёлые плотные рёбра, как у морских коров, но его конечности плохо сохранились. Следующая по возрасту окаменелость (протосирен) имеет укороченные задние лапы, слабо связанные с позвоночником, так что это животное обитало в основном в воде. Требовалось найти ископаемое животное, у которого имелись бы явно сиреновые рёбра и череп, а ноги пригодны для ходьбы по суше. Это было бы решающим доказательством происхождения морских коров от сухопутных предков.
В дебрях Ямайки
Райский тропический остров Ямайка совсем не похож на бесплодные пустоши, которые обычно показывают в документальных фильмах об искателях окаменелостей, но на Ямайке действительно были сделаны некоторые важные палеонтологические находки. Примерно в 15 км к югу от курортного городка Монтего-Бей, в районе под названием Семь Рек, что в приходе Сент-Джеймс, есть примечательные залежи костей. Роджер Портелл и другие палеонтологи годами собирали материалы в формации Чейпелтон — том самом нижнеэоценовом подразделении, где нашёлся череп оуэновского прорастомуса. Портелл искал окаменелости моллюсков, в частности гигантской морской улитки
Campanile и других вымерших одностворчатых и двустворчатых. Все кости в этих отложениях были фрагментарными, поскольку когда-то их смыло в древнюю лагуну или дельту, где они смешались с останками морских моллюсков. Здесь находили кости игуан, примитивных носорогов и, возможно, лемуроподобных приматов.
К середине 1990-х годов коллекции окаменелостей из формации Чейпелтон выросли, и на них обратил внимание Дэрил Домнинг из Говардского университета, крупнейший эксперт по ископаемым морским коровам. В этих местах могли быть найдены и другие экземпляры сиреновых, похожих на прорастомуса, обнаруженного 150 годами ранее. Домнинг плодотворно провёл на Ямайке несколько сезонов, добыв сотни костей. Но вместо новых окаменелостей прорастомуса он обнаружил совершенно новый род и вид примитивных морских коров — более того, ему удалось найти практически полный скелет!
В 2001 году Домнинг опубликовал собственноручно подготовленное описание образца в известнейшем научном журнале Nature. Он назвал существо
Pezosiren portelli («ходячая сирена Портелла») (рис. 21.4). Пезосирен сравним по размеру с крупной свиньёй (около 2,1 м в длину), его череп почти как у прорастомуса. Он имел некоторые более примитивные черты, чем прорастомус (например, по верхней части черепа шёл гребень), но в целом был более высокоразвитым животным (например, это касается области уха и формы кончика нижней челюсти, обращённого книзу). Кроме того, у пезосирена имелись типичные толстые и плотные рёберные кости, характерные для всех сиреновых. Пезосирен обладал крепким длинным бочкообразным телом и коротким хвостом. Очень важно, что в этой окаменелости почти полностью сохранились тазовые кости, передние и задние лапы. Пусть лапы и короткие, они заканчиваются обычными кистями и ступнями, ничуть не специализированными для плавания. Судя по деталям конечностей и позвоночника, пезосирен плавал вдоль дна на мелководье, перебирая лапами, примерно как гиппопотам, а не двигал хвостом вверх-вниз (так плавают выдры, сиреновые, киты, морские котики и морские львы).

 Рис. 21.4.
Рис. 21.4. Пезосирен: (
А) Дэрил Домнинг с реконструированным скелетом
P. portelli; (
Б) реконструкция внешнего облика ((
А) иллюстрация предоставлена Daryl Domning; (
Б) иллюстрация предоставлена Nobumichi Tamura)
Так было найдено недостающее звено между водными морскими коровами и их сухопутными предками, идеальная промежуточная форма. У пезосирена был такой череп и такие увесистые рёбра, как у сиреновых, при этом он имел полностью развитые лапы и ступни четвероногого существа. Подобно многочисленным ходячим китам и другим окаменелостям, открытым за последние годы, пезосирен демонстрирует, как ещё одна группа сухопутных животных вернулась в море.
Из Африки
Оставалось сложить последний фрагмент мозаики. Ближайшие родичи морских коров — слоны и прочие тетитерии, возникли в Северной Африке, Пакистане и других регионах по берегам моря Тетис. Но древнейшие ископаемые сиреновые (прорастомус и пезосирен) найдены на Ямайке. В 2013 году группа учёных, в которую входили Жюльен Бенуа (руководитель) и ещё девять человек, опубликовала информацию о новых раннеэоценовых экземплярах возрастом около 50 млн лет из Туниса. Среди находок было несколько черепных костей с захватом области уха, явно сиреновых по форме, а также другие фрагменты скелета. Образцы обнаружили в местечке Шамби, поэтому описываемое животное условно назвали шамбийской морской коровой (кости слишком фрагментарны, чтобы давать им официальное таксономическое название). При всей неполноте образцов область уха шамбийской морской коровы сохранилась хорошо, и из этого можно заключить, что древнейшие сиреновые — равно как родственные им древнейшие хоботные, даманы и прочие тетитерии — возникли в районе Тетиса (в основном в Африке). Морские коровы были водными животными и вскоре расселились от Карибского моря до Индии (рис. 21.5). Хоботные, даманы и прочие тетитерии оставались в Африке, откуда выбрались всего 16 млн лет назад через Аравийский полуостров, а затем распространились по всему миру. Вскоре мамонты и мастодонты уже обитали на всех континентах Северного полушария, слоны пришли в Азию, а даманы расселились по многим регионам Евразии. Мир навсегда изменился.
 Рис. 21.5.
Рис. 21.5. Эволюционная история сиреновых (рисунок Mary P. Williams на основе материалов Дэрила Домнинга)
Стеллерова корова
В начале XVIII века император Пётр Великий стремился расширить границы Российской империи и усилить её влияние в мире. Он пытался модернизировать и цивилизовать российскую политику и социальный уклад, в частности, хотел привить европейские обычаи и способствовал развитию науки и образования, как это делалось во всех передовых европейских государствах: Франции, Англии, Нидерландах, некоторых княжествах Германии. Он снаряжал морские экспедиции на дальние окраины своей империи — к берегам Сибири и особенно тихоокеанскому побережью, где издавна оставались неизученными целые регионы, такие как Камчатка.
На флоте Петра Великого с 1704 года служил капитан Витус Беринг, датчанин по национальности. К 1725 году он занимался исследованием регионов к северу от Камчатского полуострова, на тот момент почти неизвестных. Беринг полагал, что между Азией и Северной Америкой лежит море, но не успел зайти достаточно далеко на северо-восток, прежде чем экспедиции пришлось вернуться на Камчатку. Много лет он выбивал поддержку и снаряжение, подыскивал людей, чтобы предпринять более масштабную экспедицию и выяснить, что находится к северо-востоку от Камчатки. Наконец, в 1741 году Беринг повёл несколько кораблей с многочисленными экипажами в те края, где посетил многие из Алеутских островов, добрался до острова Кадьяк и материковой Аляски — так произошла первая высадка европейцев на эти берега. Погода была плохой и ветреной, корабли несколько раз теряли друг друга, а один потерпел крушение. Команда этого корабля болела цингой, люди умирали: они питались только морским зверем и рыбой, но не имели фруктов и не получали витамин C. К августу 1742 года уцелевшие члены экипажа одного из кораблей, сумевшие починить его после крушения, добрались до берегов России. Сам Беринг умер в пути и был похоронен на острове близ Камчатки, который сегодня носит его имя. В честь него также названы Берингово море, Берингов пролив и ледник Беринга.
Во второй экспедиции участвовал немецкий естествоиспытатель Георг Стеллер. Его наняли в качестве главного натуралиста, и, к счастью для потомков, он описал животный мир Северной Пацифики таким, каков он был к моменту прибытия туда первых европейцев. Стеллер убедил Беринга отпускать его в пешие вылазки на берег и стал первым европейцем, ступившим на землю Аляски. Стеллер открыл немало видов млекопитающих и птиц, многие животные названы в его честь. В частности, стеллерова черноголовая голубая сойка (полярная сойка с характерной чёрной головой и хохолком, обитающая в горах на западе Северной Америки), а также множество уязвимых видов: гигантский стеллеров морской лев, стеллерова гага и белоплечий орлан. Ещё два вида уже истреблены: речь об очковом баклане и стеллеровой морской корове.
Когда охотившиеся моряки не могли прокормиться одними каланами, они взялись за безобидную стеллерову корову. Это было огромное животное, крупнейшее морское млекопитающее того времени, если не считать китов (рис. 21.6).

 Рис. 21.6.
Рис. 21.6. Стеллерова морская корова: (
А) скелет, выставленный в Музее сравнительной зоологии Гарвардского университета; (
Б) сравнение размеров стеллеровой коровы и человека ((
А) фотография автора; (
Б) рисунок Mary P. Williams)
Корова имела 8–9 м в длину и весила 7–9 т. Стеллеровы коровы были совершенно кроткими и не боялись человека, хотя туземные охотники перебили большую часть этих животных — их осталось всего несколько тысяч в нескольких небольших популяциях. Вот что Стеллер писал в своём рапорте:
По всему побережью острова, особенно там, где ручьи впадают в море и во множестве имеются всякие водоросли, морская корова... встречается в любое время года, они пасутся здесь большими стадами. Самые крупные коровы имеют четыре–пять морских саженей в длину (7–9 м), а также до трёх с половиной морских саженей в толщину (около 2,25 м) в области пупа — там они толще всего. Вплоть до пупа корова похожа на сухопутное животное, а далее и до хвоста — на рыбу. Голый череп её ничуть не отличим от лошадиного, но мясистая голова более напоминает буйволовую, особенно губы. Глаза животного лишены век и не больше, чем у овцы... Брюхо пухлое и очень объёмистое, а также неизменно столь набитое, что при малейшем ранении в живот содержимое внутренностей коровы с шипением вырывается наружу. Пропорционально оно напоминает живот лягушки... Подобно обычному скоту, эти животные обитают в море стадами, самцы и самки обычно держатся вместе, перемещаются вдоль всего побережья вместе с молодняком. Этих животных не интересует ничего, кроме еды. Спина и половина брюха постоянно видны над водой, коровы всё время пасутся, точно как сухопутные животные, медленно двигаясь вперёд. Они соскабливают лапами водоросли с камней и непрестанно жуют... во время отлива они удаляются с мелководья в море, но с приливом вновь возвращаются к отмели, порой они подплывали так близко, что их можно было достать шестом... Они ничуть не боятся людей. Когда корова хочет отдохнуть, она ложится кверху брюхом в тихом местечке рядом с бухтой и плавает как поплавок. Я не заметил у них признаков особой смышлёности... но они исключительно любят друг друга, настолько, что если одну корову загарпунить, остальные стараются её спасти, не дать вытянуть на берег — для этого они смыкаются вокруг неё кольцом. Другие пытались перевернуть ялик. Некоторые налегали на канат либо пытались извлечь гарпун из тела товарки, что им действительно несколько раз удавалось. Мы также видели, как самец однажды двое суток подряд подплывал к берегу, где лежало тело мёртвой самки, проверял, что с ней. Тем не менее коровы всё время держались в одних и тех же местах, независимо от того, сколько их было ранено или убито. Жир этих животных не маслянистый и не расплывчатый, а довольно плотный и железистый, белоснежный; а если он несколько дней полежит на солнце, приобретает такой приятный жёлтый цвет, как самое лучшее голландское масло. Вываренный жир гораздо слаще и вкуснее наилучшего говяжьего сала, по цвету и текучести напоминает свежее оливковое масло, сладок, как миндальное масло, исключительно приятно пахнет и очень питателен. Мы пили его целыми чашками, не чувствуя ни малейшей тошноты... Мясо старых животных неотличимо от говядины, но по сравнению с мясом любого сухопутного и морского зверя обладает примечательным свойством: даже в жаркие летние месяцы оно отлично хранится на воздухе, не протухает две недели и даже больше, хотя мясные мухи сразу же оскверняют его и мясо полностью покрывается червями.
Как только известия об открытиях Стеллера и Беринга достигли России, русские охотники и торговцы пушниной пустились на Камчатку и Алеутские острова, к берегам Берингова моря. Землепроходцы стреляли и ели практически всех животных, каких могли поймать. Охотились в основном на каланов ради их ценного меха, но убивали и тюленей, морских львов, моржей, китов, других животных. Небольшую популяцию стеллеровых коров, насчитывавшую всего несколько тысяч особей, истребили быстро: их убивали на мясо или просто из развлечения. К 1768 году эти крупнейшие сиреновые исчезли — всего через 27 лет после того, как их открыл Стеллер.
Лучше один раз увидеть!
Оригинальные окаменелости пезосирена нигде не экспонируются, но их слепки можно увидеть в Геологическом музее Вест-Индского университета (Мона, Ямайка), Центре исследования и охраны природы Спэниш-Бей (Белиз), Национальном музее естественной истории (Париж) и в Национальном музее природы и науки (Токио).
Скелеты стеллеровой морской коровы есть в коллекции 27 организаций по всему миру, в некоторых музеях они выставлены на всеобщее обозрение. Это Музей сравнительной зоологии Гарвардского университета (Кембридж, Массачусетс), Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия), Музей естественной истории (Лондон), Национальный музей Шотландии (Эдинбург), Национальный музей естественной истории (Париж), Музей естественной истории (Лион), Государственный музей естественной истории (Брауншвейг, Германия), Музей естественной истории (Вена), Шведский музей естественной истории (Стокгольм), Зоологический музей (Лунд, Швеция), Финский музей естественной истории (Хельсинки), Государственный музей природоведческой истории при Академии наук Украины (Киев), Зоологический музей при Зоологическом институте РАН (Санкт-Петербург) и Зоологический музей Московского государственного университета.
Дополнительная литература
Berta, Annalisa, and James L. Sumich. Return to the Sea: The Life and Evolutionary Times of Marine Mammals. — Berkeley: University of California Press, 2012.
Berta, Annalisa, James L. Sumich, and Kit Ь. Kovacs. Marine Mammals: Evolutionary Biology. 2nd ed. — San Diego: Academic Press, 2005.
Janis, Christine M., Gregg F. Gunnell, and Mark D. Uhen, eds. Evolution of Tertiary Mammals of North America. Vol. 2, Small Mammals, Xenarthrans, and Marine Mammals. — Cambridge: Cambridge University Press, 2008.
Prothero, Donald R., and Robert M. Schoch. Horns, Tusks, and Flippers: The Evolution of Hoofed Mammals and Their Relatives. — Baltimore: Johns Hopkins University Press, 2002.
Rose, Kenneth D., and J. David Archibald, eds. The Rise of Placental Mammals: The Origin and Relationships of the Major Extant Clades. — Baltimore: Johns Hopkins University Press, 2005.
22. Лошадь зари
Происхождение лошадей
Эогиппус
Геологические свидетельства о происхождении лошади — один из классических примеров эволюции.
Уильям Диллер Мэтью. «The Evolution of the Horse»
Цок-цок
Когда Колумб в 1492 году прибыл на берега Карибского моря, в обеих Америках не было ни одной лошади. Первых домашних лошадей он привёз в Западное полушарие во время своей второй экспедиции в 1493-м. В 1521 году Эрнан Кортес завоевал империю ацтеков. Конкистадорам помогло не только их огнестрельное оружие и распространявшиеся повсюду заразные болезни, но и лошади. Впервые увидев испанских всадников, ацтеки пришли в ужас и сочли, что человек и лошадь — одно существо наподобие кентавра.
Вскоре лошади распространились в Западном полушарии. Они стали главным транспортным средством и важнейшим вьючным животным: именно для такой работы их тысячелетиями использовали в Европе и Азии. Лошади преобразили культуру индейцев Великих Равнин, которые вскоре стали искусными наездниками, научились охотиться и сражаться верхом. На лошадях индейцы могли вести кочевую жизнь, следуя за стадами бизонов. Лошади стали основой культуры Старого Запада, особенно когда эксплуатация огромных скотоводческих ранчо стала невозможна без участия ковбоев. Однако из-за появления двигателя внутреннего сгорания и автомобиля к 1920-м годам лошадь практически «устарела», что стало очевидно в годы Первой мировой войны, когда кавалерийские подразделения оказались особенно уязвимы для новых видов оружия. Сегодня лошади — статусный символ богатства, хотя в мире остались регионы, где фермерство и культура содержания лошадей по-прежнему очень важны.
Вплоть до 1807 года считалось, что древние лошади обитали только на территории Евразии, но в этом году Уильям Кларк, совершивший знаменитую экспедицию вместе с Льюисом, нашёл кости североамериканских лошадей в местечке Биг-Боун-Лик в штате Кентукки, где ранее обнаруживали окаменелости вымерших мастодонтов, мамонтов, гигантских ленивцев и прочих животных ледникового периода. Он отправил окаменелости своему покровителю, президенту Томасу Джефферсону, увлекавшемуся палеонтологией, но Джефферсон никогда ничего не писал о важности этой находки.
10 октября 1833 года во время экспедиции на «Бигле» молодой Чарльз Дарвин посетил Аргентину. Он «просто изумился», обнаружив зубы и кости ископаемых лошадей в том же пласте, где находились вымершие гигантские глиптодонты, напоминавшие броненосцев (по размеру их панцирь был сравним с «Фольксвагеном-жуком»). Эти окаменелости показали, что в Америке не просто существовали аборигенные лошади — они жили бок о бок с вымершими животными последнего ледникового периода. Дарвин передал все эти ископаемые знаменитому британскому палеонтологу Ричарду Оуэну, который назвал этих лошадей
Equus curvidens и отметил: «Доказательства существования в Южной Америке животных этого рода, который был там истреблён, а впоследствии повторно интродуцирован на континенте, являются одним из интереснейших палеонтологических открытий мистера Дарвина». Затем, в 1848 году Джозеф Лейди, основатель американской палеонтологии, опубликовал работу о многочисленных изученных им экземплярах лошадей ледникового периода, в которой заключил, что разнообразные лошади обитали в Северной Америке задолго до прибытия Колумба.
Тем временем европейские палеонтологи тоже находили ископаемых лошадей. Попадались не только многочисленные плейстоценовые представители современного рода
Equus, жившие в ледниковом периоде, но и более примитивные лошади из более древних отложений — например, гиппарион, относившийся к позднему и среднему миоцену; анхитерий, живший в раннем миоцене. Также был открыт палеотерий, но оказалось, что он не относится ни к настоящим лошадям, ни вообще к семейству лошадиных. В 1872 году «бульдог Дарвина» Томас Генри Гексли указал, что развитие лошадей в Европе прослеживается по четырём родам, последовательно сменявшим друг друга. Годом позже русский палеонтолог Владимир Онуфриевич Ковалевский развил эту идею.
Открытия новых ископаемых лошадей следовали одно за другим, по мере того как американские палеонтологи — Лейди, Эдвард Дринкер Коп из Академии естественных наук в Филадельфии и Отниел Чарльз Марш из Йельского университета принялись за описание больших коллекций, прибывавших с американского Запада. В 1871 году Марш назвал орогиппусом ископаемую лошадь из Скалистых гор, а в 1872 году Коп назвал древних эоценовых лошадей эогиппусами. Когда Гексли в 1876 году прибыл в США, чтобы выступить с серией лекций по случаю столетия американского государства, он планировал пропагандировать идеи Дарвина и рассказать об эволюции лошадей в Европе. В ходе поездки учёный посетил Марша в Йеле и провёл за изучением его коллекции целых два дня. Его сын Леонард Гексли, написавший биографию отца, рассказывает: «Всякий раз, требовался ли ему образец, чтобы проиллюстрировать тот или иной тезис либо чтобы продемонстрировать переход от более древних и неспециализированных форм к более поздним и совершенным, профессор Марш просто обращался к ассистенту, просил его найти коробку с таким-то номером, пока Гексли, наконец, не сказал: “Думаю, вы волшебник. Чего бы я ни попросил, вы сразу мне это предоставляете”». Затем Гексли пересмотрел свои заметки и доработал лекцию таким образом, чтобы в ней была описана эволюция американских лошадей (рис. 22.1).
 Рис. 22.1.
Рис. 22.1. «Генеалогия лошади». Иллюстрации Отниела Чарльза Марша, описывающие трансформацию зубов и конечностей у лошадей на основании окаменелостей из его коллекции, собранных в Северной Америке (из работы
Marsh O. C. Polydactyl Horses, Recent and Extinct //
American Journal of Science and Arts. — 1879. — Vol. 17)
Вскоре стало понятно, что лошади развивались преимущественно в Северной Америке, а европейские лошади, такие как анхитерий и гиппарион, — переселенцы, которые произошли от основной американской линии. В 1926 году палеонтологи, в частности Уильям Диллер Мэтью, составили упрощённую схему, демонстрировавшую эволюцию лошадей с течением времени (рис. 22.2). У крошечных эоценовых лошадок было три-четыре пальца, зубы имели невысокую коронку и были приспособлены для поедания листьев и фруктов. Затем в олигоцене появились мезогиппус и миогиппус, у которых было по три пальца, а ноги и пальцы стали гораздо длиннее. Ещё позже существовали миоценовые лошади, в частности меригиппус, с ещё более длинными ногами и пальцами, редуцированными боковыми пальцами и зубами с более высокой коронкой для поедания жёсткой травы. Наконец, в плиоцене и плейстоцене эта серия завершается лошадьми (Equus), у которых длинные ноги и всего один палец, боковые пальцы редуцированы до крошечных нефункциональных костных щепок. Зубы лошади имеют очень высокую коронку.
 Рис. 22.2.
Рис. 22.2. Упрощённая схема Уильяма Мэтью, демонстрирующая эволюцию лошадей, изменения их зубов и скелета, а также простые линейные трансформации с течением воемени (из работы
William Diller Matthew. The Evolution of the Horse: A Record and Its Interpretation //
Quarterly Review of Biology. — 1926. — Vol. 1)
За следующие 90 лет после публикации схемы наши знания об эволюции лошадей значительно обогатились. На смену упрощённой линейной последовательности пришла сложная разветвлённая структура, где разные эволюционные линии лошадей существуют бок о бок (рис. 22.3). Например, только в каменоломне Рэйлуэй-Куорри, в миоценовой формации Валентайн на севере штата Небраска нашлось 12 экземпляров ископаемых лошадей, причём все эти животные жили в этом месте одновременно. Мы с Нилом Шубином изучали миогиппуса и мезогиппуса, и наше исследование показало, что в какой-то момент были современниками три вида мезогиппусов и два вида миогиппусов — все они обнаруживаются на одном и том же уровне, в одних и тех же отложениях Биг-Бэдлэндс в Южной Дакоте, а также в аналогичных породах Вайоминга и Небраски.
 Рис. 22.3.
Рис. 22.3. Более современная схема эволюции лошадей, напоминающая ветвистое дерево (рисунок К. Д. Протеро по работе
Donald R. Prothero. Mammalian Evolution //
Donald R. Prothero and Robert M. Schoch, eds. Major Features of Vertebrate Evolution. — Knoxville, Tenn.: Paleontological Society, 1994)
«Лошадь зари»
Что насчёт древнейших лошадей? Как они выглядели и жили? Ископаемые лошади повсеместно встречаются в нижнеэоценовых отложениях (сформировались 55–48 млн лет назад) на западе Северной Америки, особенно в формации Уилвуд бассейна реки Бигхорн в Вайоминге, в формации Уосатч в бассейнах рек Уинд и Паудер в Вайоминге, а также в формации Сан-Хосе в штате Нью-Мексико. Там были добыты буквально тысячи зубов и челюстей, а кроме того, несколько хороших, частично сохранившихся скелетов. Большинство первых лошадей были размером с лису или бигля (250–450 мм в высоту). Много лет в разных книгах этих лошадок ошибочно сравнивают с более мелким фокстерьером, копируя данные из публикации Генри Фэрфилда Осборна — богатого джентльмена, увлекавшегося охотой на лис.
По сравнению с потомками, древнейшие лошади отличались короткой головой и мордой, небольшим мозгом, а их зубы имели очень невысокие коронки и короткие корни. Щёчные зубы состояли из множества поперечных выступов и низких бугорков: такая адаптация помогала питаться нежным кормом, например листьями и фруктами (см. рис. 22.1 и 22.2). Конечности и пальцы у этих лошадей были сравнительно короткими, хотя животные бегали на кончиках пальцев, а также хорошо прыгали (рис. 22.4). На передних лапах у них было по четыре пальца (мизинец совсем крошечный, а
большой палец полностью утрачен, поэтому они ходили на трёх пальцах), а на задних — по три коротких пальца (без большого и мизинца). Они имели длинный костистый хвост, примерно как у кошки, а не такой редуцированный, с длинными волосами, какой впоследствии развился у лошадей. Короче говоря, если бы вы увидели такую лошадку, никогда не приняли бы её не то что за лошадь, даже за мельчайшего карликового пони. Скорее она напомнила бы вам носуху или другое животное, ничуть не похожее на лошадь; впрочем, их невозможно сравнивать с какими-либо современными млекопитающими.

 Рис. 22.4.
Рис. 22.4. Североамериканские лошади раннего эоцена: (
А) собранный практически полный скелет; (
Б) реконструкция внешнего облика ((
А) фотография предоставлена Смитсоновским институтом; (
Б) иллюстрация предоставлена Nobumichi Tamura)
Палеонтологические данные свидетельствуют, что эти крошечные лошади были превосходно приспособлены к жизни в густых джунглях и в перегретом климате, царившем в раннем эоцене. Тогда атмосфера была так насыщена углекислым газом, что даже на полюсах сложились вполне тепличные условия: там обитали крокодилы, аллигаторы (пусть по полгода стояла полярная ночь), а также лошади и тапиры. Такие места, как Монтана и Вайоминг (из пород на территории этих штатов и были добыты описываемые окаменелости), выглядели совсем иначе, нежели сегодня. Ныне там суровые степи, где выпадает много снега, а морозы стоят долгими месяцами. В эоцене это был край тропических лесов.
В джунглях обитали не только лошади, но и многочисленные тапиры, напоминавшие лошадей, и многие другие примитивные копытные млекопитающие. Кроны деревьев кишели приматами, похожими на лемуров, а также другими, ныне вымершими древесными зверями. Существовали и древние хищные млекопитающие, но они были не крупнее волка. В отсутствии конкуренции суперхищником эоценовых джунглей была нелетающая птица, достигавшая 2,5 м в высоту, имевшая огромный клюв и крошечные крылья. Вид, обитавший в Скалистых горах, называется диатрима, европейский вид — гасторнис. Добычей этих птиц были палеотериевые — например, палеотерий — близкие родичи лошадей. Палеотерии не были настоящими лошадьми, которые бы относились к основной североамериканской эволюционной линии, но они занимали лошадиную «нишу» в раннеэоценовой Европе.
Как же их называть?
Основная дилемма, связанная с этими существами, сводилась к тому, как их называть. Первое название эоценовым лошадям Северной Америки (
Orohippus angustidens) дал Марш в 1875 году, основываясь на сильно повреждённых образцах зубов и челюстей, добытых в Нью-Мексико и относящихся к раннему эоцену. Затем, в 1876 году Марш назвал некоторых раннеэоценовых ископаемых лошадей эогиппусами (в переводе с греческого «лошадь зари»), взяв за основу экземпляр
E. Validus — хороший, частично сохранившийся скелет. Многие другие хорошо сохранившиеся экземпляры тоже были причислены к роду
Eohippus. Вскоре стало очевидно, что раннеэоценовый эогиппус совсем не походил на среднеэоценового орогиппуса, поэтому название закрепилось за лошадьми раннего эоцена. Оно стало широко использоваться в начале XX века; вы найдёте его практически на всех сравнительно старых схемах, описывающих эволюцию лошадей (в том числе в популярных учебниках).
В 1932 году сэр Клайв Форстер Купер из Британского музея естественной истории заметил, что окаменелости американских лошадей исключительно похожи на ту, которую описал Ричард Оуэн в 1841 году. Эта находка, добытая в формации Лондонская глина, датируемой ранним эоценом, была названа гиракотерием («зверь, похожий на дамана»). Поскольку это название появилось на 35 лет раньше, чем эогиппус, по правилу приоритета Международного кодекса зоологической номенклатуры оно стало валидным наименованием для древней лошади — если эогиппусы и гиракотерии суть одни и те же животные. Эту точку зрения в 1951 году поддержал блестящий палеонтолог Джордж Гейлорд Симпсон, и она получила широкое признание. На протяжении почти всей оставшейся части XX века древние эоценовые североамериканские и европейские лошади причислялись к гиракотериям. Это название также по-прежнему встречается во многих книгах и других источниках, не поспевающих за вечно меняющейся наукой.
Однако наука не стоит на месте, обнаруживаются всё новые, хорошо сохранившиеся экземпляры, меняется и философия именования окаменелостей. В начале XX века палеонтологи увлекались таксономической сегрегацией, придумывая отдельное видовое и родовое название практически для каждой новой окаменелости, независимо от того, сколь незначительны различия между находками. В 1930-е и 1940-е годы палеонтологи начали присматриваться к нормальному диапазону изменчивости, который прослеживается среди диких популяций в естественных условиях, и вскоре осознали, что многие признаки, по которым выделялись новые виды, были нормальными проявлениями именно такой изменчивости в рамках одного вида. Подобное «популяционное мышление» преобладает с 1940-х и далее; большинство палеонтологов предпочитают причислять многочисленные и слегка различающиеся окаменелости к одному виду, если нет убедительных сведений об их анатомии, распределении в пространстве и времени либо статистической информации о нормальной внутривидовой изменчивости, которые действительно позволили бы считать этих животных представителями разных видов по нормам современной биологии.
За последние годы появилось много новых экземпляров (особенно хорошо сохранившихся экземпляров с ранее не известными анатомическими деталями), заставивших учёных вернуться к изучению окаменелостей, ранее попадавших в таксономические «мусорные корзины». Согласно новым представлениям о классификации (так называемой кладистике), «мусорные корзины» бессмысленны с эволюционной точки зрения и не являются естественными группами организмов. Соответственно, они не заслуживают официального таксономического наименования. Так, многие называют рыбами всех водных позвоночных, не являющихся четвероногими (тетраподами). Однако двоякодышащие рыбы находятся в гораздо более тесном родстве с тетраподами, чем с костистыми рыбами. Следовательно, они значительно ближе к человеку, чем к бесчелюстным рыбам. В современной таксономии больше не используется общее название «рыбы» (
Pisces), поскольку оно отражает факт современной экологии, но не описывает естественную группу, имеющую свою характерную эволюционную историю.
Действительно, после более внимательного изучения первых окаменелостей и открытия новых, которые сохранились лучше, пришлось отказаться от идеи причислять всех раннеэоценовых североамериканских и европейских лошадей к гиракотериям. Сначала, в 1989 году Джереми Хукер из Лондонского музея естественной истории проанализировал все окаменелости гиракотериев, добытые в формации Лондонская глина, и пришёл к выводу, что это не лошади, а европейские палеотерии. Таким образом, уже нельзя использовать слово «гиракотерии» в качестве удобного общего наименования для всех раннеэоценовых североамериканских лошадей. Некоторые учёные не согласны с этим выводом, но не потому, что располагают иными фактами или обоснованными анализами. Просто они так долго считали всех североамериканских лошадей гиракотериями, что не могут расстаться с привычкой. Затем, в 2002 году Дэвид Фрёлих из Техасского университета внимательно проанализировал экземпляры всех американских раннеэоценовых лошадей. Он обнаружил, что ко всем этим животным неприменимо общее родовое название, так как они оказались сброшены в огромнейшую таксономическую «мусорную корзину» на основании примитивных характеристик. Название эогиппус можно оставить только для экземпляра
angustidens (Коп) и
validus (Марш), а большинство представителей раннеэоценовых лошадей, долго именовавшиеся гиракотериями или эогиппусами, не могут быть отнесены ни к одному из этих родов, должны быть причислены к новым родам или к старым, которые можно возродить. Например, род
Protorohippus, названный так Якобом Вортманом в 1894 году, включает сравнительно высокоразвитые виды лошадей, например
montanum и
venticolum. Фрёлих выделил некоторые новые рода лошадей, в частности
Sifrhippus — для карликовой лошадки
sandrae, жившей в самом начале эоцена;
Minippus — для видов
index и
jicarillai, а также
Arenahippus — для видов
grangeri,
aemulor и
pernix. Некоторые виды, например
tapirinum, открытый Копом, относились не к лошадям, а к другим непарнокопытным, в частности к тапирам, и сегодня называются
Systemodon tapirinum.
Соответственно, не существует единого рода эоценовых лошадей, запомнив который было бы легко заучивать их названия и подписывать схемы. Мы не можем просто называть их всех эогиппусами и удовлетвориться таким вариантом, поскольку это фактическая ошибка и чрезмерное упрощение. Природа гораздо затейливее и разнообразнее, чем наше мышление и схемы; необходимо корректировать наши взгляды в соответствии с новейшими исследованиями. Так, нельзя употреблять давно признанное некорректным наименование «бронтозавр» или называть Плутон планетой. Итак, любая схема эволюции лошадей в любом учебнике ошибочна, если все раннеэоценовые лошади проходят в ней под общим родовым названием, будь то эогиппус или гиракотерий. На современной схеме, если она претендует на научную актуальность, должны быть перечислены как минимум
Protohippus,
Sifrhippus,
Minippus и
Arenahippus.
Откуда же взялась лошадь?
Когда я учился в университете, мы проходили курс по палеонтологии позвоночных, и там был заключительный проект: профессор Майкл Вудбёрн давал каждому студенту большую смешанную подборку костей раннеэоценовых животных из бассейна реки Бигхорн, найденных вблизи города Имблем в штате Вайоминг. Мы должны были рассортировать их, идентифицировать, вооружившись научной литературой, и составить список видов, которые нам выпали. Это была сложная задача, поскольку в те годы практически отсутствовала актуальная классификация раннеэоценовых животных. Ситуация изменилась после того, как Пол Джинджерич, Кеннет Роуз, Дэвид Краузе и Томас Боун опубликовали уйму статей об ископаемых животных из бассейна Бигхорн. Эти работы выходили с конца 1970-х.
Насколько всё упростилось бы, будь у нас эти статьи в 1975-м!
Вот что мне больше всего запомнилось из этого проекта. Мой лоток был полон челюстей древних непарнокопытных, прежде всего, лошадей (как бы они сегодня ни назывались), а также гомогалаксов — древних родичей тапиров. Отсеять первые от вторых было дьявольски трудно, хотя мой младший сын уже в возрасте двух лет отличал тапира от лошади! Сегодня кажется, что зубы и анатомию лошадей и тапиров не перепутаешь, но 55 млн лет назад у них были практически идентичны не только зубы, но и, по большей части, весь череп и скелет (рис. 22.5). Имелись лишь считанные малозаметные отличия (например, насколько непрерывны поперечные эмалевые выступы у лошади по сравнению с тапиром). Чтобы их замечать, требовался опыт и острый глаз.
 Рис. 22.5.
Рис. 22.5. Эволюционная радиация примитивных непарнокопытных (по образцу
Donald R. Prothero. Evolution: What the Fossils Say and Why It Matters. — New York: Columbia University Press, 2007. — Fig. 14.5)
Если взять других раннеэоценовых непарнокопытных, эта тенденция ещё выразительнее. Древнейший предок носорога, так называемый гирахиус, едва отличим от древних тапиров и лошадей, хотя современные носороги, тапиры и лошади совсем не похожи. Первые представители бронтотериев — вымерших млекопитающих, напоминавших носорогов, тоже были очень похожи на древних носорогов, тапиров и лошадей. Иными словами, исключительно разнообразные современные непарнокопытные уже в раннем эоцене восходят к общему предку, ничуть не походившему на своих нынешних потомков. Затем, в ходе эволюционной дивергенции, генетические линии его потомков постепенно разошлись, и теперь этих животных легко отличить друг от друга. Действительно, к среднему эоцену чётко оформились эволюционные линии лошадей, тапиров, носорогов и бронтотериев, и различить их мог бы даже ребёнок, хотя ни одно из этих животных не походило на своих современных потомков.
Откуда же взялись лошади и их непарнокопытный род? Палеонтологи долго указывали в качестве предков лошадей группу древних копытных млекопитающих, фенакодонтовых, широко распространённых в палеоцене и раннем эоцене. Зубы у них были почти как у древних непарнокопытных, а черепа и скелеты обладали всеми чертами, позволявшими идентифицировать их как общего предка непарнокопытных. Но в 1989 году Малколм Маккенна и трое его китайских соавторов описали новую позднепалеоценовую окаменелость, найденную в Монголии (возраст — около 57 млн лет). Они назвали её радинския, в честь Леонарда Радински, одного из отцов-основателей исследований непарнокопытных, умершего в 1986 году (см. рис. 22.5). Радинския выглядит точь-в-точь как маленькая лошадь, с той оговоркой, что она примитивнее самых древних лошадей. Маккенна с коллегами были озадачены тем, сколь примитивна эта окаменелость, и не уверены, классифицировать её как непарнокопытное либо отнести к другой группе млекопитающих, близких к непарнокопытным. Впоследствии большинство учёных согласились, что радинския доказывает следующий факт: стремительная эволюция непарнокопытных в раннем эоцене на территории Европы и Северной Америки объясняется не тем, что они якобы произошли от местных фенакодонтид. Просто непарнокопытные проникли в Европу и Северную Америку из Азии около 55 млн лет назад, а затем стали быстро видоизменяться, вытесняя большинство древних автохтонных копытных млекопитающих (в том числе своих близких родичей фенакодонтид), вплоть до их полного вымирания к концу среднего эоцена.
Превосходная палеонтологическая летопись истории лошадей демонстрирует не только изначальное сходство и дивергентную эволюцию непарнокопытных, которая началась в Азии, но и эволюцию лошадей в Северной Америке, а также их исчезновение в Западном полушарии в эпоху плейстоцена. Им было суждено вернуться домой только в конце XV века.
Лучше один раз увидеть!
Во многих американских музеях есть экспонаты, демонстрирующие эволюцию лошадей. Обычно можно увидеть раннеэоценовую лошадь, олигоценового мезогиппуса из пустошей Уайт-Ривер (штат Южная Дакота), несколько миоценовых лошадей и плейстоценовую лошадь
Equus. Речь, в частности, об Американском музее естественной истории (Нью-Йорк), Музее естественной истории под открытым небом (Чикаго), Флоридском музее естественной истории при Флоридском университете (Гейнсвилл), Национальном музее естественной истории Смитсоновского института (Вашингтон, округ Колумбия) и о Музее естественной истории округа Лос-Анджелес (Лос-Анджелес).
Дополнительная литература
Franzen, Jens Lorenz. The Rise of Horses: 55 Million Years of Evolution. / Translated by Kirsten M. Brown. — Baltimore: Johns Hopkins University Press, 2010.
MacFadden, Bruce J. Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae. — Cambridge: Cambridge University Press, 1994.
Prothero, Donald R., and Robert M. Schoch, eds. The Evolution of Perissodactyls. — New York: Oxford University Press, 1989.
Prothero, Donald R., and Robert M. Schoch, eds. Horns, Tusks, and Flippers: The Evolution of Hoofed Mammals and Their Relatives. — Baltimore: Johns Hopkins University Press, 2002.
23. Гигантские носороги
Крупнейшее сухопутное млекопитающее
Парацератерий
Все мы прекрасно знали, что белуджитерий был существом гигантским. Но величина костей повергла нас в изумление.
Мы принесли лишь переднюю часть черепа с несколькими зубами. Для Грепджера этого, однако, было достаточно:
«Для меня всё ясно, — сказал он, — белуджитерий — это гигантский безрогий носорог. Другого такого зверя наука не знает!»
Рой Эндрюз. «Диковинные звери»
Зыбучие пески!
В 1922 году знаменитый палеонтолог Генри Фэрфилд Осборн — директор Американского музея естественной истории, ведущий учёный и общественный деятель своего времени, отправил экспедицию в Монголию, где предполагалось найти окаменелости древнейших предков человека. Осборн ошибочно полагал, что человек сформировался в Азии, и использовал этот повод, чтобы выбить солидное финансирование у богатых спонсоров и попечителей музея. Экспедицию организовали с размахом: караван из 75 верблюдов (на каждого навьючили по 400 кг припасов и бензина), три кузовных автомобиля Dodge, два грузовика Fulton, большая компания учёных и ассистентов. Во главе экспедиции стоял легендарный Рой Чепмен Эндрюз — бесстрашный исследователь и искатель приключений. Многие считают, что именно он стал прототипом киногероя Индианы Джонса. Осборн сказал Эндрюзу: «Окаменелости там. Я уверен. Поезжайте и найдите их».
Экспедиция выдвинулась из Пекина, миновала Великую Китайскую стену и вскоре прославилась находками удивительных окаменелостей, принадлежавших динозаврам мелового периода; в том числе Эндрюс с коллегами нашли первую известную кладку динозавровых яиц. Но, несмотря на успешные поиски окаменелостей, учёные не получили доказательств существования древнейших людей в Азии. Просто Осборн ошибался, а Дарвин был прав: древние люди развивались в Африке. По иронии судьбы, первая настоящая окаменелость древнего человека (
Australopithecus africanus (Ребёнок из Таунга; см. главу 25)) была найдена в Южной Африке в 1924 году, в то самое время, когда Осборн и Эндрюз добивались от спонсоров дополнительных средств на поиски предков человека в Азии. Осборн, как и большинство учёных того времени, раскритиковал африканскую окаменелость, сочтя её черепом молодой обезьяны, возраст которого неизвестен.
Кроме эффектных окаменелостей динозавров, палеонтолог Уолтер Грейнджер и помогавшие ему китайцы нашли много важных и впечатляющих окаменелостей млекопитающих. Вот что Эндрюз писал о третьей экспедиции в 1925 году в своей образной книге с очень неполиткорректным, империалистическим названием «Новое покорение Центральной Азии»:
«Честь наиболее важного открытия, сделанного в Лохе, принадлежит одному из наших сборщиков-китайцев, Лю Хси-Ку. От его цепкого взгляда не ускользнуло поблёскивание белой кости в красной осадочной породе на крутом склоне холма. Он немного там покопался и сообщил о находке Грейнджеру, который довёл раскопки до конца. С каким изумлением он обнаружил ступню и нижнюю часть ноги стоящего белуджитерия, словно животное просто позабыло её здесь, сделав следующий шаг (см. рис. 23.1). Окаменелости так редко встречаются в подобном положении, что Грейнджер просто сел и задумался, что к чему. Здесь мог быть лишь один ответ: зыбучие пески! Лю нашёл правую заднюю ногу; следовательно, правая передняя нога должна была располагаться ниже по склону. Грейнджер определил, куда двигалась нога, отмерил около девяти футов
[7] и стал копать. Что бы вы думали — она там была, огромная кость, напоминавшая ствол окаменевшего дерева, и она тоже стояла прямо. Не составило труда найти и две левые конечности, ведь было очевидно, что здесь произошло. Когда мы раскопали все четыре ноги, каждую в отдельной яме, зрелище открылось экстраординарное (см. рис. 23.1). Мы с Грейнджером забрались на вершину холма и стали грезить о днях, когда случилась эта трагедия. Умея читать язык окаменелостей, мы легко восстановили всю историю по четырём обломкам костей. Вероятно, чудовище подошло к пруду, чтобы напиться, а вокруг были коварные зыбучие пески. Вдруг зверь стал тонуть. Положение ног показывает, что он немного подался назад, опёршись на ляжки и отчаянно пытаясь выбраться из затягивающего песка. Вероятно, он утонул быстро, но боролся до последнего, и умер лишь после того, как нос и глотка забились густой грязью. Если бы он увяз не полностью и умер от истощения, тело завалилось бы набок. Если бы мы могли откопать целый скелет, прямо стоящий в такой могиле, это был бы экземпляр на загляденье всему миру.
Я спросил Грейнджера: «Уолтер, что вы имеете в виду, когда говорите, что рассчитываете найти только ноги? Почему не всё остальное?» — «Не взыщите, — ответил он, — но, если бы мы не опоздали и прибыли сюда тридцать пять тысяч лет назад, когда холм ещё не снесло эрозией, я предъявил бы вам весь скелет целиком!» В самом деле, мы опоздали на какие-то тридцать пять тысяч лет. Когда выветрилась осадочная порода, похоронившая гиганта, кости постепенно рассыпались и теперь валяются на дне долины тысячью бесполезных фрагментов. Должно быть, в Монголии во времена олигоцена обитало множество белуджитериев, поскольку фрагменты их костей мы находим во всех окаменелых отложениях той эпохи.
 Рис. 23.1.
Рис. 23.1. Кости ног парацератерия были найдены в вертикальном положении, будто лежали с тех пор, как животное увязло в зыбучих песках (негатив № 285735, публикуется с разрешения библиотеки Американского музея естественной истории)
Рассказ Эндрюза получился красочным, но подробности, вероятно, несколько отличались. Зыбучие пески лишь в кино стремительно засасывают жертву по самую макушку. В реальности зыбучий песок — это просто песок, насыщенный водой. Под давлением он разжижается и обтекает ноги жертвы, которые постепенно тонут. Но всё же это в основном вода, человек или животное не могут погрузиться в него глубже, чем в бассейн, где остаются на плаву. Чтобы выбраться из зыбучего песка, на нём нужно распластаться (как на воде), а затем кто-то должен протянуть вам палку или верёвку и вытащить.
Вероятно, утонувшее в зыбучем песке животное завязло в нём не более чем по брюхо. Потом зыбучий песок затвердел, и зверь умер от жажды. Оставшаяся часть тела, вероятно, не погрузилась в песок и стала лёгкой добычей для падальщиков, растерзавших мёртвого или умирающего зверя, который не мог двинуться с места.
Монгольские чудовища
Белуджитерии, о которых разговаривали Эндрюз и Грейнджер, были гигантскими безрогими носорогами, сегодня известными под именем парацератерий. Отдельные зубы этих животных находят с 1907 года, но первые хорошо сохранившиеся экземпляры в 1910 году открыл британский палеонтолог Клайв Форстер Купер — в Белуджистане, регионе на территории современного Пакистана. Экземпляры представляли собой считанные повреждённые черепа и челюсти, а также отдельные кости; по ним было сложно понять, насколько колоссально это животное. В 1913 году Купер дал название
Baluchitherium osborni сравнительно полному черепу из своих коллекций, и это наименование оставалось популярным на протяжении десятилетий. Через четыре года русский палеонтолог Алексей Алексеевич Борисяк назвал другой скелет — самый полный из найденных на тот момент — индрикотерием (рис. 23.2).
 Рис. 23.2.
Рис. 23.2. Единственный относительно полный скелет парацератерия, выставленный в Палеонтологическом музее им. Ю. А. Орлова в Москве (фотография предоставлена M. Fortelius)
Все три названия широко использовались десятилетиями, хотя учёные отвергали неофициального белуджитерия, которое жаловал и продвигал Осборн (один из видов этого рода назвали в его честь), так как это явно был отдельный экземпляр парацератерия. Поскольку название «парацератерий» дано раньше, белуджитерий является младшим синонимом. В 1989 году Спенсер Лукас и Джей Собус доказали, что существовала всего одна популяция этих животных, отличавшихся сильной изменчивостью; следовательно, старейшее название
Paraceratherium имеет приоритет над более поздними
Baluchitherium и
Indricotherium. Большинство учёных, работающих с данными окаменелостями, соглашаются, что
Paraceratherium, скорее всего, был представлен одним родом, включавшим максимум три-четыре вида, поскольку огромным животным, чтобы нормально питаться, нужны обширные пастбища. По экологическим причинам представляется маловероятным, что множество видов таких животных могли одновременно населять огромные территории в одном и том же регионе.
Как бы ни назывались эти невероятные звери, они паслись на широких просторах Азии в олигоцене и раннем миоцене примерно 33–18 млн лет назад. Их окаменелости были найдены не только в Монголии и Пакистане, но и в некоторых регионах Китая, Казахстана, сравнительно недавно — в Турции и Болгарии.
Парацератерий — крупнейшее из когда-либо найденных млекопитающих (рис. 23.3). Он достигал 4,8 м в холке, 8 м в длину, был тяжелее любого слона или мастодонта. Сначала его вес оценивали в 34 т, но новейшие методы позволяют снизить величину до 20 т, что немногим больше веса крупнейших из когда-либо живших родичей слонов, дейнотерид.
 Рис. 23.3.
Рис. 23.3. Реконструкция парацератерия, выполненная из стекловолокна, изначально выставленная в музее при Университете штата Небраска, а ныне находящаяся в Центре Риверсайд в городе Скоттсблафф (штат Небраска). Для сравнения показан современный слон (на заднем плане), а у ног парацератерия (справа на переднем плане) находится реконструкция гиракодона — животного, от которого произошли парацератерии (публикуется с разрешения музея при Университете штата Небраска)
У парацератерия был огромный череп длиной более 2 м с коротким хоботом (судя по глубокому носовому отверстию в черепе). Губа с хватательным придатком, приспособленная для ощипывания веток, а также сравнительно примитивные зубы с низкой коронкой, пригодные только для жевания листьев, но не для питания жёсткой травой (см. рис. 23.4). Нет никаких признаков, что на черепе у него имелся рог, то есть парацератерий был безрогим (как и большинство вымерших носорогов). В большей части реконструкций его изображают просто как увеличенного носорога, с маленькими носорожьими ушами. Однако я указывал, что при такой массе у него должны были возникать проблемы с теплоотдачей. Поэтому, чтобы не перегреваться, потребовались бы большие «радиаторы» (например, уши, как у слона).
 Рис. 23.4.
Рис. 23.4. Громадный череп парацератерия, найденный в Монголии участниками экспедиции, снаряжённой Американским музеем естественной истории в 1922 году (негатив № 310387, публикуется с разрешения библиотеки Американского музея естественной истории)
Хотя парацератерий и был крупнее любого слона, у него были относительно длинные запястные и голеностопные кости, так как он произошёл от более мелких родичей носорогов (от гиракодонтов), имевших длинные конечности и лапы, приспособленные для бега (рис. 23.5; см. также рис. 23.3). Несмотря на огромный размер и массу животного, конечности парацератерия так и не приобрели пропорций, типичных для динозавров-зауропод и слонов. Пальцы ног у них очень короткие и сильно уплощены из-за огромного веса, который им приходится нести. Короче говоря, парацератерий был носорогом, попытавшимся занять экологическую нишу жирафов и питаться листвой древесных крон. При этом у него выросли не только ноги и шея, как у жирафа, но и всё остальное.
 Рис. 23.5.
Рис. 23.5. Реконструкция гиракодона — родича носорогов размером примерно с немецкого дога. Именно от гиракодона произошёл парацератерий (рисунок R. Bruce Horsfall)
Откуда пошли носороги
Парацератерий возник в рамках грандиозной эволюции носорогов, развернувшейся в кайнозое. Древнейшие носороги найдены в нижнеэоценовых отложениях (им около 52 млн лет), и они очень похожи на самых древних тапиров и лошадей (см. рис. 22.5). К концу среднего эоцена (40 млн лет назад) носороги разделились на три семейства. Представители семейства аминодонтовых, ныне вымершие, явно приспособились к полуводной жизни, как гиппопотамы. У них были крупные челюсти и череп, как у бегемотов, а также грузное тело и короткие лапы.
Второе вымершее семейство — гиракодонтовые, или «бегающие носороги». Представители этого семейства были распространены на территории Северной Америки и Азии в позднем эоцене.
Наиболее изученный вид
Hyracodon nebraskensis (см. рис. 23.5) найден в пустошах Уайт-Ривер. Это было животное размером примерно с немецкого дога, длинными костями конечностей, кистей и ступней. По костям понятно, что гиракодон бегал быстрее любого носорога, жившего до или после него. Гиракодонтовые в дальнейшем процветали в Азии, где стали крупнее и совершеннее. Чуть более крупным представителем гиракодонтовых был китайский род
Juxia — животные размером с лошадь, длинной шеей и длинными же ногами. Далее от этих носорогов произошёл уртинотерий, по размеру сравнимый со слоном, и, наконец, парацератерий.
К третьему семейству носороговых принадлежат современные африканские и азиатские носороги. В прошлом это семейство включало десятки родов и видов, стремительно развивавшихся в Евразии и Северной Америке (в конце концов достигших Африки).
Североамериканские носороги вымерли около 5 млн лет назад, в конце миоцена, а носороги продолжали существовать в Евразии вплоть до конца последнего ледникового периода (20 тыс. лет назад), когда были почти полностью истреблены. Сегодня уцелело всего пять видов носорогов, относящихся к четырём родам: два африканских, индийский, а также находящиеся на грани исчезновения яванский и суматранский носороги.
Все пять видов подвергаются большой опасности из-за хищнической охоты за их рогом, используемым в традиционной китайской медицине. (С медицинской точки зрения носорожий рог не представляет ценности, так как состоит из слипшейся спресованной шерсти; его химический состав примерно такой же, как у человеческих волос и ногтей.) Масштабы охоты таковы, что дикие носороги были бы истреблены за несколько лет, если бы не огромные усилия по их защите: один грамм перетёртого в порошок носорожьего рога стоит дороже грамма золота или кокаина.
Биология гигантских носорогов
Поскольку парацератерий был немногим крупнее слонов, мы можем в значительной мере восстановить его образ жизни и биологию по аналогии со слоном. Вероятно, небольшие стада этих животных бродили по обширным территориям, удовлетворяя недюжинный аппетит. По-видимому, они догола ощипывали кроны деревьев. На основе изучения слонов и с учётом биомеханических ограничений, эти животные должны были двигаться медленно, развивая скорость не более 30 км/ч, обычно — около 10–19 км/ч. Однако с такими длинными ногами они могли обходить большие территории даже неторопливым шагом. Высота и большие размеры этих животных указывают на то, что их сердце билось медленно (сердце слона делает всего 30 ударов в минуту). Крупные размеры также позволяют предположить, что эти животные должны были жить долго, примерно как нынешние слоны (вероятно, средний возраст составлял 50–70 лет).
Популяция парацератериев должна была оставаться немногочисленной, причём самки рожали одного детёныша где-то раз в год. Вероятно, он становился взрослым примерно к десяти годам. Видимо, большую часть жарких дневных часов парацератерии отдыхали в тени и купались в прудах, чтобы не перегреться, а в более прохладное время суток — вечером, ночью и ранним утром — почти непрерывно ели. Как и у современных лошадей, носорогов и слонов, пища у них переваривалась сравнительно неэффективно, с ферментацией в толстом кишечнике. У них не было такого мощного четырёхкамерного желудка, как у жвачных: быков, баранов, коз, антилоп, жирафов, оленей и т. п. Следовательно, как лошади и слоны, парацератерии должны были ежедневно потреблять огромное количество пищи, но переваривали незначительную её часть по сравнению со жвачными млекопитающими.
Такое огромное тело могло доставлять неудобства (особенно связанные с теплоотдачей), но давало и преимущества. Как и современные слоны, парацератерии могли не опасаться хищников, поскольку на такую крупную дичь едва ли можно было успешно охотиться. Большинство хищников, найденных в олигоценовых отложениях Азии, были мельче волка, поэтому никто из них не мог потягаться со взрослым парацератерием. Но детёныши были уязвимы. Вероятно, парацератерии, как и слоны, жили небольшими стадами, где основную роль играли самки: старшая, её сёстры, дочери и племянницы. Они все вместе оберегали детёнышей и молодняк, пока те не вырастали достаточно крупными, чтобы не бояться хищников.
Страна гигантов
В олигоцене на нынешней территории Монголии и Китая сложилась очень специфическая экосистема — сразу по нескольким позициям. В большинстве мест, где находят окаменелости парацератериев, в изобилии встречаются окаменелости грызунов и зайцеобразных. То есть в этой экосистеме было мало ресурсов для средних травоядных и очень много — для крупных. Очевидно, что ранее данную территорию покрывали засушливые полупустынные кустарниковые степи; мало больших лугов и очень немного травоядных животных. Гигантские парацератерии объедали древесные кроны, а немногочисленные средние травоядные, похожие на антилоп, кормились кустарником (в современных саваннах и высокотравных степях, наоборот, обитает много мелких травоядных). Поскольку хищники были относительно мелкими, не ровня взрослому парацератерию, они определённо обгладывали скелеты мёртвых парацератериев и питались другой падалью, какую удавалось найти.
Причина вымирания парацератериев вызывает дебаты, но, вероятно, исчезновение этих животных произошло по двум причинам. На протяжении большей части олигоцена и раннего миоцена парацератерий господствовал в своей экосистеме, не сталкиваясь ни с крупными хищниками, ни с серьёзными конкурентами-травоядными. Затем, 20–19 млн лет назад, первые мастодонты покинули африканскую прародину и расселились по Евразии. Примерно в то же время они достигли Северной Америки. Современные слоны и их доисторические предки коренным образом повлияли на окружающую среду (в современной африканской саванне они валят деревья и прореживают лесные чащобы, способствуя распространению разнообразной растительности). Без их вмешательства ничто не мешало бы росту деревьев. Вероятно, с прибытием мастодонтов в Евразию там стали быстро деградировать леса, особенно вековой лес, необходимый парацератериям. Кроме того, вслед за мастодонтами в Евразию мигрировали охотившиеся на них хищники. Это были и амфиционовые размером с медведя (медведесобаки — вымершее семейство, не связанное ни с псовыми, ни с медвежьими) и гигантские гиенайлюры. Такие крупные хищники были способны задирать крупных мастодонтов и, вероятно, оказались непобедимы для парацератериев, давно не сталкивавшихся с опасными противниками. Как бы то ни было, парацератерии вымерли вскоре после того, как в Евразию пришли мастодонты и охотившиеся на них плотоядные.
Индрикотерий в массовой культуре
Будучи крупнейшим известным науке сухопутным млекопитающим, парацератерий фигурирует во многих фильмах и прочих источниках. Ему посвящён целый эпизод в сериале «Прогулки с чудовищами», рассказывавший об олигоцене в Восточной Азии. Аниматоры проделали отличную работу, восстановив повадки, которые могли бы быть у этих животных. Однако у нас имеются лишь кости парацератерия и ориентировочное представление о его биологии — по аналогии со слонами. На самом деле, мы ничего не знаем ни о масти этих животных, ни о том, какие звуки они издавали, ни как себя вели.
Очевидно, медленный мерный аллюр этих колоссальных зверей нашёл отражение не только в документалистике. Аниматор Фил Типпетт, разрабатывавший большую часть декораций и реквизита для ленты «Империя наносит ответный удар», второго эпизода сериала «Звёздные войны», явно вдохновлялся парацератериями, создавая модели высоченных грохочущих и шагающих танков AT-AT, присланных на ледяную планету Хот для подавления мятежа.
Лучше один раз увидеть!
Почти полные скелеты парацератерия экспонируются лишь в Палеонтологическом музее им. Ю. А. Орлова в Москве и в некоторых крупных китайских музеях. Наиболее хорошо сохранившийся череп парацератерия есть в Американском музее естественной истории (Нью-Йорк), другие выставлены в Лондонском музее естественной истории и Музее наук о земле им. Седжвика в Кембриджском университете.
Много лет модель парацератерия, выполненная из стекловолокна, экспонировалась в музее при Университете штата Небраска в городе Линкольн (см. рис. 23.3). Позднее она оказалась в Музее Риверсайд города Скоттсблафф (штат Небраска).
Дополнительная литература
Prothero, Donald R. Rhinoceros Giants: The Paleobiology of Indricotheres. — Bloomington: Indiana University Press, 2013.
24. Отражение обезьяны?
Древнейший ископаемый предок человека
Сахелянтроп
В следующий раз, когда будете в зоопарке, прогуляйтесь мимо обезьяньих клеток. Представьте обезьян почти без шерсти, а рядом с ними — клетку с несчастными людьми, голыми и неспособными говорить, но в остальном нормальными. А теперь попытайтесь угадать, насколько наш генофонд схож с генофондом этих обезьян. Каково, по-вашему, совпадение генетического кода у вас и у шимпанзе — 10 процентов, 50 процентов, 99 процентов?
Джаред Даймонд. «Третий шимпанзе»
Отражение обезьяны?
Тема эволюции человека, протекавшей параллельно с эволюцией животных, всегда вызывала полемику и эмоции. По религиозным причинам даже сегодня значительное количество людей не приемлет идею о том, что человек — представитель царства животных, просто ещё один вид. Хотя в большинстве развитых современных государств этот факт не подвергается сомнению. Некоторые опросы свидетельствуют, что многие американцы признают эволюцию растений и животных, но не эволюцию человека.
По иронии судьбы, эволюции человека посвящено так много исследований, и эта тема вызывает такой интерес, что именно человек — один из наиболее изученных примеров эволюции. Целая антропологическая дисциплина (физическая антропология и палеонтология человека) посвящена изучению палеонтологической летописи наших ближайших родичей. Тысячи учёных по всему миру исследуют целый комплекс тем в этой сфере — палеоантропологов больше, чем специалистов по динозаврам и любым другим доисторическим существам. Без малого сотни тысяч окаменелостей гоминин (представителей подсемейства Homininae) хранятся в музеях по всему миру. Количество экземпляров столь ошеломительно, а информация об эволюции человека так подробна, что, если бы мы говорили о любом другом биологическом виде с нашей планеты, человек был бы образцовым примером эволюции, документированным не хуже, чем развитие любого другого семейства животных. Но столько людей упрямо выдвигают к этой идее ненаучные претензии, что она препарируется с излишней щепетильностью, искажается и попросту отвергается. Если бы аналогичный массив доказательств был собран для решения любого иного вопроса, никаких сомнений уже не осталось бы.
И даже если бы у нас не было чрезвычайно полной палеонтологической летописи человека, остальные доказательства
всё равно крайне убедительны. Нам достаточно просто взглянуть в зеркало. Ещё в 1735 году основатель современной систематики Карл Линней дал людям научное наименование
Homo sapiens (человек разумный) и охарактеризовал наш вид греческой фразой
«Познай себя». В 1766 году Жорж-Луи Леклерк, граф де Бюффон написал в 14-м томе своей «Естественной истории», что обезьяна — «просто животное, но животное необыкновенное, на которое человек не может смотреть, не думая одновременно о себе». Другие французские натуралисты, в частности Жорж Кювье и Этьен Жоффруа Сен-Илер, отмечали исключительное анатомическое сходство между обезьянами и людьми, хотя не говорили прямо, что человек — вид обезьяны. Один из первопроходцев биологии, французский учёный Жан-Батист Ламарк, в 1809 году приводил следующую аргументацию в своей книге «Философия зоологии»:
Определённо, если бы определённая порода обезьян, особенно наиболее совершенные из них, утратили бы в силу обстоятельств или по какой-то другой причине привычку карабкаться по деревьям и хвататься за ветви ногами... и если бы представители этой породы, поколение за поколением, были вынуждены использовать ноги только для ходьбы, а на руках ходить бы перестали, то, несомненно... эти обезьяны превратились бы в двуруких существ, и... ноги их служили бы тогда исключительно для ходьбы.
Эта проблема имела высокую важность и в 1859 году, когда Дарвин опубликовал свою работу «Происхождение видов». Его книга и так была противоречивой, поэтому он пытался максимально сгладить проблему эволюции человека, оставив лишь одно замечание на данную тему: «Много света будет пролито на происхождение человека и его историю». Хотя Дарвин в тот момент отказывался распространяться по этому поводу, его сторонник, Томас Генри Гексли бросился на амбразуру и в 1863 году опубликовал книгу «О положении человека в ряду органических существ». В этой работе Гексли описал и проиллюстрировал анатомическое сходство всех костей, мышц и органов человекообразных обезьян и человека (рис. 24.1). Наконец, в 1871 году Дарвин изложил собственные мысли в книге «Происхождение человека», хотя уделил основное внимание таким темам, как половой отбор, даже не упоминая окаменелости — на тот момент ещё не было палеонтологических доказательств эволюции человека (кроме неандертальцев, но их интерпретировали неверно).
 Рис. 24.1.
Рис. 24.1. Иллюстрация Бенджамина Уотерхауса Хокинса, демонстрирующая исключительное сходство скелетов человека и человекообразных обезьян — вплоть до отдельных костей (из книги
Thomas Henry Huxley. Evidence as to Man’s Place in Nature. — London: Williams & Norgate, 1863)
Перенесёмся на 90 лет вперёд. Ещё один источник данных, не доступный ни Дарвину, ни другим учёным, работавшим до 1960-х годов, чётко демонстрирует наше родство с обезьянами и другими представителями царства животных. Речь о ДНК. Уже самые первые опыты по молекулярной биологии показали исключительное сходство ДНК человека, шимпанзе и гориллы. Если поместить сыворотку антител человека и этих обезьян в один и тот же раствор, иммунные реакции получаются сильнее, чем между сывороткой человека и любых других животных. Это значит, что гены человека и человекообразных обезьян в основном схожи.
Затем, в конце 1960-х, разработали метод под названием «гибридизация ДНК-ДНК». Раствор с ДНК обезьяны и человека нагревается до тех пор, пока две нити двойной спирали не расплетаются. После этого смесь остужается, и каждая нить сплетается с ближайшей. Некоторые получившиеся молекулы ДНК содержат и человеческую, и обезьянью нить. Некоторые нити обезьяньей ДНК сплетаются с обезьяньими, а некоторые нити человеческой ДНК — с человеческими, но нас
интересуют двойные спирали гибридной ДНК. Если повторно нагреть раствор с гибридной ДНК, то чем крепче сплетены нити гибридных молекул (то есть чем более они схожи), тем более высокая температура требуется, чтобы их разделить. Проделывая такой опыт с ДНК шимпанзе, горилл, других человекообразных обезьян, а также мартышек, лемуров и прочих животных (не приматов), можно примерно определить, насколько то или иное животное схоже с человеком. Этот опыт также свидетельствует, что ДНК человека и шимпанзе практически идентичны.
За последние 20 лет появились такие передовые технологии, как полимеразная цепная реакция (ПЦР). Они позволяют непосредственно секвенировать не только человеческую ДНК, но и ДНК других животных и растений. Геном человека был полностью секвенирован в 2001 году, а геном шимпанзе — в 2005-м. Сравнив их, мы получили такой же результат, как и при гибридизации ДНК-ДНК: геномы человека и шимпанзе совпадают на 98–99%. Как от шимпанзе, так и от горилл нас отличает 1–2% ДНК. Дело в том, что 60–80% нашего генома составляет так называемая «мусорная ДНК». Эти гены никогда не считываются и не используются, но пассивно передаются из поколения в поколение. Часть «мусорной ДНК» — эндогенные ретровирусы (ERV), обрывки вирусных ДНК, попавших в наш геном после инфицирования каких-то далёких наших предков и по-прежнему передаваемых потомству, хотя эти участки ДНК ничего не кодируют. Меньший процент приходится на структурные гены, кодирующие каждый белок и строение нашего тела, в том числе гены, которые мы давно не используем. 1–2% генов, отличающие нас от шимпанзе, — это гены-регуляторы, своеобразные «переключатели», сообщающие остальному геному, когда какие признаки должны быть выражены, а когда — нет. Именно поэтому люди сильно отличаются от человекообразных обезьян, хотя набор генов у них и у нас почти идентичен.
Например, и у людей, и у человекообразных обезьян есть структурные гены, отвечающие за отращивание длинного хвоста, но экспрессия этих генов не происходит, кроме редких случаев, когда в регуляторных генах возникает сбой. Если произойдёт такая ошибка, у человека вырастет длинный костистый хвост. У птиц тоже есть гены, кодирующие появление длинного костистого хвоста, как у динозавров (унаследованные от хищных предков-рептилий), но поскольку они не работают, у современных птиц вместо хвоста — сросшиеся кости гузки. Время от времени регуляторные гены отказывают, и птенцы вылупляются с длинным хвостом, как у динозавра. Аналогично, у современных птиц беззубый клюв, тогда как у их предков-динозавров имелись зубы (см. главу 18). Однако гены для отращивания зубов остались. Экспериментально удалось привить эмбриону курицы эпителиальную ротовую ткань, взятую у мыши, и вылупился зубастый цыплёнок. Но зубы у него были не мышиные, а динозавровые! Следовательно, в ДНК животных есть множество древних генов, которые «не включаются», но достаточно немного изменить генетическую регуляцию, чтобы возродить примитивные черты.
Исключительное генетическое сходство между человеком и двумя видами шимпанзе (речь об обыкновенном шимпанзе (
Pan troglodytes) и карликовом шимпанзе, или бонобо (
P. paniscus), само по себе — ошеломительное и очень убедительное доказательство нашего близкого родства. Несмотря на инстинктивное неприятие со стороны некоторых людей и религиозные идеи, человек — действительно отражение обезьяны. Вот как говорит об этом биолог Джаред Даймонд: «...допустим, на Землю прибыли инопланетные зоологи, и весь биоматериал, который оказался у них в распоряжении, — это ДНК. Они секвенировали геномы разнообразных животных, в том числе ДНК человека и двух шимпанзе. Опираясь лишь на эти данные, инопланетяне пришли бы к выводу, что человек — просто третий вид шимпанзе. Наше генетическое сходство с двумя другими видами шимпанзе выше, чем между любыми двумя видами лягушек, даже выше, чем между львами и тиграми».
Действительно, различия в ДНК всех человеческих рас меньше, чем аналогичные различия у отдельных популяций шимпанзе из разных регионов Африки (см. рис. 24.2)! Это позволяет сделать два вывода. Во-первых, генетические различия между человеческими расами незначительны, они менее существенны, чем кажется многим людям. Во-вторых, внешнее несходство между человеком и шимпанзе обусловлено минимальными изменениями в регуляторных генах, но результаты этих изменений грандиозны.
 Рис. 24.2.
Рис. 24.2. Молекулярная филогения человекообразных обезьян и людей, демонстрирующая генетическое расстояние между разными видами, в зависимости от их митохондриальной ДНК. Все человеческие расы больше похожи друг на друга, чем разные популяции горилл и шимпанзе (иллюстрация по образцу Sequences Show Diverse Evolutionary Histories of African Hominoids //
Proceedings of the National Academy of Sciences USA. — 1999. — Vol. 93. — Fig. 1B; © 1999, Национальная академия наук США)
Дело закрыто: человек — немного изменившаяся обезьяна. Генетические и анатомические доказательства более чем убедительны. ДНК в каждой клетке вашего тела — свидетельство и свидетельница нашего близкого родства с обезьянами, независимо от того, насколько неловко чувствуют себя от этого некоторые люди. Этот факт доказан и без окаменелостей, которые демонстрировали бы переход от обезьян к человеку. Но как давно разошлись эволюционные линии человека и обезьян?
Геологические часы
Учёные двумя путями подходят к вопросу о том, когда разделились эволюционные линии человека и человекообразных обезьян. Во-первых, ведут поиск окаменелостей, в которых постепенно становится всё больше обезьяньих черт и меньше человеческих. Эта стратегия развивается на протяжении всего времени, пока ведутся исследования, хотя её успех зависит от того, удастся ли найти подходящие породы подходящего возраста, и окажутся ли в них окаменелости примитивных гоминин. Человеческие кости фоссилизируются очень редко, поэтому даже в подходящих отложениях можно найти лишь считанные человеческие зубы или челюсти, хотя там же будут сотни окаменелостей, принадлежащих другим млекопитающим: антилопам, свиньям, мастодонтам. Тем не менее, как мы узнаем из главы 25, палеоантропологам понадобились годы раскопок, чтобы отыскать исчезающие и редкие человеческие окаменелости, поскольку на любом важном открытии можно сделать карьеру и имя.
Когда окаменелости гоминин найдены, следующая непростая задача — точно их датировать. Многие человеческие окаменелости обнаружили в пещерах и других местах, где нет материалов, которые облегчали бы датировку. Если экземпляр моложе 60 тыс. лет (последний ледниковый период до голоцена), органические ткани окаменелости можно датировать с помощью радиоуглеродного анализа (учитывает содержание углерода-14). Археологи активно пользуются этим методом для датировки возраста человеческих артефактов, большинство из которых моложе 60 тыс. лет, а также окаменелостей, сформировавшихся в конце ледникового периода. Например, окаменелости, найденные в битумных озёрах Ла-Брея близ Лос-Анджелеса не старше 37 тыс. лет, поэтому их многократно датировали в ходе радиоуглеродного анализа. Данный метод не работает с материалами старше 60 тыс. лет (в лучших современных лабораториях удаётся отодвинуть предел до 80 тыс. лет). Для работы с более древними окаменелостями больше подходит калий-аргоновый (K-Ar) метод либо его новая версия, когда для датировки используются изотопы
40Ar и
39Ar. В таком случае окаменелость нельзя датировать непосредственно, по материалу экземпляра или осадочным породам, в которых он был найден. Вместо этого сравниваются кристаллы, сформировавшиеся в породах, остывших после вулканических извержений. Это может быть древняя лава или древний вулканический пепел. Когда такие кристаллы застывают, в их атомной решётке остаётся нестабильный материнский изотоп калий-40. По мере старения кристалла часть атомов калия-40 спонтанно распадается, и образуется дочерний изотоп аргон-40. Скорость распада хорошо известна, поэтому, измерив соотношение между материнскими и дочерними изотопами, геолог может вычислить возраст кристалла.
Как в любом другом научном методе, здесь есть свои ограничения и подводные камни, которых нужно избегать. Поскольку датировка — измерение времени с момента застывания кристалла и попадания в него радиоактивных материнских атомов, калий-аргоновый метод работает лишь в породах, когда-то перешедших из жидкого состояния в твёрдое (они называются магматическими). Таковы, например, граниты и базальты. Хороший геолог скажет вам, что кристаллы из песчаника или другой осадочной породы непригодны для датировки: они попали в песчаник из более древних пород, их возраст не связан с возрастом осадочной толщи. Но геологи давно обошли эту проблему, найдя по всей Земле сотни мест, где пригодные для датировки пласты лавы или вулканического пепла перемежаются с осадочными породами, содержащими окаменелости, либо где интрузивные магматические породы вклиниваются в осадочные и позволяют определить их минимальный возраст. На основе таких отправных точек вычисляется возраст пород в геологических масштабах, причём метод настолько точен, что мы можем определить время большинства событий, произошедших в период от нескольких миллионов до ста тысяч лет назад.
Если из-за строения кристалла часть материнских или дочерних атомов каким-то образом его покинула либо атомы попали в кристаллическую решётку и загрязнили кристалл, соотношение материнских и дочерних изотопов искажается и уже не годится для датировки. Геологи всегда учитывают такой вариант и проверяют десятки образцов, чтобы точно установить возраст, а также делают перекрёстную проверку, прибегая к другим методам датировки. Новейшие методы и инструменты настолько точны, что опытный геолог способен уловить любую ошибку датировки и быстро отметает даты, не соответствующие строгим стандартам.
С помощью названных методов большинство окаменелостей, найденных в Африке, датировали очень точно, их возраст удалось определить, хотя они пролежали в земле более 25 млн лет (см. главу 25). Антропологи часто работают вместе с геохронологами и ищут свежие слои, где много неповреждённых кристаллов подходящих минералов (как правило, речь идёт о калиевых полевых шпатах либо о слюдах — мусковите и биотите). Эта работа не осталась без ошибок, но в целом хронология окаменелостей гоминин восстановлена хорошо. Кроме того, если в определённом районе нет залежей пепла, палеонтологи могут ориентироваться на хронологические различия комплексов окаменелостей, чтобы хотя бы примерно определить возраст данного местонахождения. Ведь в другом месте такой же комплекс окаменелостей может встретиться вместе с подходящим для датировки вулканическим пеплом.
Какова эта палеонтологическая летопись? История начинается с важных окаменелостей, которые были найдены в гряде Сивалик на территории Пакистана. Эта замечательная серия пород охватывает большую часть олигоцена, миоцена и плиоцена, причём невероятно богата окаменелостями. Залежи образовались на месте речных наносов, рассыпавшихся по Южной Азии во время медленного роста Гималаев: горы понемногу разрушались, и в результате возник Сивалик. Палеонтологи и геологи изучали породы с 1902 года, когда английский учёный Гай Пилгрим первым начал исследования в Южной Азии, которая тогда была колонией Великобритании.
В течение прошлого века на территории Сивалика собрали огромные коллекции окаменелостей млекопитающих, позволяющие подробно восстановить, как в Южной Азии протекала эволюция в период миоцена. Из-за напряжённости индо-пакистанской политики, а также в силу отношений США с обеими странами, Пакистан задолжал США миллионы долларов за приобретённое военное снаряжение. В результате, в период с 1970-х по 1990-е годы американские учёные получали массу грантов (особенно по программе Фулбрайта) на экспедиции в Пакистан и важные исследования. Многие палеонтологи воспользовались этим шансом, появились разнообразные работы по палеонтологии и геологии гряды Сивалик и прилегающих районов. Благодаря изобилию вулканического пепла и технологии, которая называется палеомагнитной стратиграфией, окаменелости Сивалика датированы исключительно хорошо. Разумеется, сегодня политическая обстановка в Пакистане так нестабильна, что очень немногие американцы рискуют туда отправиться. Даже исследователи из других стран, не имеющих тесных связей с США, опасаются обитающих во многих регионах Пакистана воинственных племён, связанных с Талибаном и Аль-Каидой.
А в 1932 году палеонтолог Дж. Эдвард Льюис из Смитсоновского института работал в долине реки Тинау, в непальской части Сивалика, и нашёл челюсть, которая очень напоминала челюсть примитивных гоминин. На ней были относительно небольшие клыки; по форме (сверху) она напоминала широкий полукруг (типично для человеческих челюстей). Напротив, челюсти человекообразных обезьян имеют U-образную форму и огромные клыки; также для них характерен плоский низкий подбородок и длинные параллельные задние кости. В 1960-е годы антрополог Дэвид Пилбим из Гарварда и приматолог Элвин Саймонс, работавший сначала в Йельском университете, затем в Университете Дьюка, а также другие учёные стали отстаивать точку зрения, что эта челюсть (её обладателя Льюис назвал рамапитеком) является древнейшей прачеловеческой окаменелостью. Рама — один из древнеиндийских героев, а слово «питек» в переводе с греческого означает «обезьяна». Также существуют приматы, названные в честь индийских богов Шивы и Брахмы. Поскольку возраст некоторых экземпляров, найденных в отлично откалиброванной серии пород Сивалика, составляет 14 млн лет, гоминины отделились от человекообразных не позднее этого периода. В 1960-е и 1970-е годы все студенты, изучавшие антропологию, эволюцию приматов и палеонтологию человека, усваивали, что рамапитек был «первым из гоминин».
Молекулярные часы
Наряду с радиоуглеродной и калиево-аргоновой датировкой существует ещё один метод, позволяющий определить, когда одна группа животных отделилась от другой, — молекулярные часы. В 1962 году легендарный молекулярный биолог Лайнус Полинг (лауреат двух Нобелевских премий) и Эмиль Цукеркандль одними из первых начали использовать молекулярные методы при построении дерева эволюционных взаимосвязей между организмами. Это были первые доказательства эволюции, добытые из наших собственных клеток и ДНК. Полинг и Цукеркандль заметили: количество различий аминокислот в молекулах гемоглобина не просто соответствует разделению эволюционных ветвей исследуемых животных; количество изменений пропорционально тому, как давно два организма отделились друг от друга. Год спустя ещё один из родоначальников молекулярной биологии, Эмануэль Марголиаш, отмечал:
Представляется, что количество остаточных различий между цитохромами-с двух любых биологических видов в основном обусловлено тем, как давно разошлись эволюционные линии, к которым относятся оба вида. Если эта гипотеза верна, цитохром-с всех млекопитающих должен в равной мере отличаться от цитохрома-с всех птиц. Поскольку рыбы отделились от основного эволюционного древа позвоночных раньше, чем птицы и млекопитающие, цитохром-с у птиц и млекопитающих должен в равной мере отличаться от цитохрома-с у рыб. Аналогично цитохром-с всех позвоночных должен в равной степени отличаться от белка дрожжей.
Все эти данные свидетельствуют о том, что молекулярные изменения накапливались со временем, по мере эволюционного расхождения разных групп животных, и что скорость изменения молекул пропорциональна давности разделения эволюционных линий.
Тем временем стали выясняться факты, свидетельствующие, что большая часть ДНК у животных является «мусорной» или как минимум нефункциональной. Огромная часть генома попросту никогда не считывается при экспрессии генов и, следовательно, остаётся невидимой для естественного отбора или адаптивно-нейтральной. Первопроходческие работы японского биохимика Мотоо Кимуры, в частности, показали, что большинство звеньев в ДНК не затрагиваются процессами, происходящими в организме. Эти «адаптивно незаметные» звенья могут спонтанно мутировать, и естественный отбор не будет отсеивать одни их варианты в пользу других. Со временем такие мутации продолжают накапливаться с регулярной скоростью — тикают, как часы. Поскольку естественный отбор не замечает таких изменений, тиканье молекулярных часов становится удобным инструментом для оценки времени расхождения между любыми двумя эволюционными линиями в геологических масштабах. Остаётся лишь откалибровать их, опираясь на хорошо известные моменты основных эволюционных расхождений, зафиксированные в палеонтологической летописи.
Вскоре многие молекулярные биологи стали плотно работать с молекулярными часами, оценивая историю расхождения разных групп животных и выверяя хронометраж. Работы покойного Винсента Сарича и Аллана Уилсона из Университета Беркли вновь и вновь демонстрировали: судя по молекулярным часам, эволюционные линии человека и шимпанзе разошлись лишь от 7 до 5 млн лет назад и не ранее 8 млн лет назад, а не 14 млн лет назад, когда жил рамапитек. Но палеонтологи держали оборону. Они скептически относились к молекулярному методу, как к недоказанному и ненадёжному, поскольку он время от времени выдавал очень странные и смехотворные результаты. (Это по-прежнему случается, и мы не всегда понимаем, почему.)
В 1970-е и 1980-е годы противоречия становились всё острее, авторитетные оппоненты вступали в пикировку на конференциях и вели жаркие споры на страницах научных журналов. Сарич и Уилсон были уверены в надёжности своих данных: наверняка что-то не так с рамапитеком либо с его возрастом. Сарич был дородным, высоченным, импозантным господином с окладистой бородой, обладал громким голосом и был неуступчив в спорах. Он без колебаний задевал и оскорблял людей, если считал нужным. В 1971 году он сказал: «Вы не имеете права называть гоминидом окаменелость старше восьми миллионов лет, плевать, как она выглядит». Разумеется, это удручало исследователей вроде Саймонса и Пилбима, которые продолжали настаивать, что рамапитек опровергает доводы молекулярных биологов.
Патовая ситуация разрешилась после ещё одного открытия, сделанного в Сивалике. В 1982 году Пилбим объявил о находке новых костей рамапитека, среди которых была не только полная нижняя челюсть, но и частично сохранившийся череп. Вместе с черепом это животное гораздо больше напоминало ископаемого орангутана, которого ещё в 1910 году назвал сивапитеком Гай Пилгрим, впервые занявшийся раскопками в Сивалике. Нижняя челюсть рамапитека принадлежала лишь ископаемому родичу орангутана, похожему на гоминин. Вскоре антропологи были вынуждены отступить и признать свою ошибку, а вместе с ней — победу Сарича, Уилсона и молекулярной биологии. Теперь, когда палеонтологи знали, что ископаемых гоминин 14 млн лет назад не существовало, встал вопрос:
какой ископаемый предок человека является самым древним? Действительно ли он не старше 8 млн лет, как предсказывали Сарич и Уилсон?
Тумай
В течение последних 25 лет палеоантропологи усердно работали по всему миру, прослеживая ископаемую историю гоминин до всё более древних отложений. Как рассказано в главе 25, первые люди появились в Африке, и все древнейшие окаменелости найдены именно там. Хотя первые раскопки велись в Южной Африке, затем — в Кении и Танзании, а с 1970-х годов усилия сосредоточились на ещё более древних пластах в Эфиопии.
С тех пор как в 1974 году открыли Люси (
Australopithecus afarensis), каждые несколько лет учёные находили более древние экземпляры. В 1984 году в Кении обнаружили окаменелости малоизвестного вида
Australopithecus anamensis. Эти образцы были старше Люси и имели возраст 5,25 млн лет. Затем в 1994 году в Эфиопии нашли ещё более примитивный вид, получивший наименование
Ardipithecus ramidus. Вплоть до 2009 года ископаемых образцов по нему почти не было, пока Тим Уайт с коллегами не объявили об открытии частично сохранившегося скелета и других ископаемых. Теперь известны многие фрагменты конечностей ардипитека и даже частично сохранившийся череп. Совсем недавно открыли ещё более древнего представителя этого рода —
Ardipithecus kaddaba, жившего 5,6 млн лет назад.
Тем временем франко-британско-кенийская команда под руководством Мартина Пикфорда работала на холмах Туген-Хиллс в Кении, породы которых гораздо старше, чем классические отложения Олдувайского ущелья и озера Туркана. В 2000 году они объявили об открытии ещё более древнего гоминина, названного
Orrorin tugenensis. В 2007 году сообщили о находке окаменелостей оррорина, сохранившихся гораздо лучше. Известно всего около 20 ископаемых фрагментов оррорина (задняя часть челюсти, передняя часть челюсти, отдельные зубы, фрагменты предплечья и бедренной кости, пальцы). Его зубы, насколько известно, очень похожи на обезьяньи, но околотазовая часть бедренной кости не оставляет сомнений, что оррорин был прямоходящим. Как и в других кенийских отложениях, на холмах Туген есть удобный для датировки вулканический пепел, позволяющий оценить возраст окаменелостей оррорина в 6,1–5,7 млн лет.
Следовательно, сегодня палеонтологическая летопись гоминин уходит минимум на 6 миллионов лет в прошлое, тогда как датировка по молекулярным часам свидетельствует, что гоминины отделились от человекообразных обезьян примерно 7–5 млн лет назад. Но где чуть более древние отложения, в которых также могли бы находиться окаменелости гоминин? К 1995 году французский палеонтолог Мишель Брюнэ много лет занимался миоценовыми млекопитающими со всего мира, специализировался на разработке наиболее опасных и удалённых залежей: в Афганистане он укрывался от проносящихся на бреющем полёте истребителей, в Ираке его арестовали, в Камеруне его коллега умер от малярии, а в Чаде учёный не раз попадал на мушку. В середине 1990-х Брюнэ часто занимался раскопками миоценовых отложений в Чаде (некогда французской колонии).
Условия в чадской пустыне Джураб — не для слабаков. Брюнэ было под шестьдесят, а работа в пустыне стала бы тяжёлым испытанием и для более молодого человека. Хотя жара могла достигать 43–49 °C, Брюнэ приходилось носить на голове повязку, надевать балаклаву и тёмные очки, чтобы уберечься от песка, забивающего глаза, уши, нос и рот. В тени могло так припекать, что бутылки с водой неожиданно взрывались. Когда Брюнэ с коллегами перелопачивали пустынный грунт в поисках фрагментов костей и зубов, им приходилось быть осторожными не только из-за адской жары, сильнейшего ветра и песка, но и потому, что эта земля была нашпигована минами, оставшимися после многочисленных племенных войн. 21 января 1995 года Брунэ нашёл челюстную кость примитивного гоминина возрастом 3,5 млн лет, первую подобную окаменелость за пределами Южной или Восточной Африки. Позже её назвали
Australopithecus bahrelghazali.
В июле следующего года он встретился с Тимом Уайтом в Аддис-Абебе, в Национальном музее Эфиопии, чтобы сравнить найденные им в Чаде окаменелости с экземплярами Уайта, добытыми в Эфиопии. Брюнэ сказал Уайту, что под той формацией, в которой он нашёл показанную челюсть, точно залегает ещё более старая. В ней были окаменелости вымерших песчанок и других млекопитающих, которым от 6 до 7 млн лет. Уайт отнёсся к этому скептически: он знал, что песчанки обитают в очень сухом климате, поэтому сомневался, что там же могли жить гоминины. Прямо в музее Брюнэ поспорил с Уайтом, что найдёт и более древних гоминид, так как докопался до более старых отложений. «Я выиграю», — сказал он.
Перенесёмся в 2001 год. Ещё шесть лет Брюнэ раскапывал более древние пласты, по возрасту относящиеся к миоцену и содержащие окаменелости, которым 6–7 млн лет. Вместе с коллегами он основал Франко-Чадскую палеонтологическую миссию (MPFT), коллаборацию Университета Пуатье и Университета Нджамены (Чад). Брюнэ и три члена чадской команды работали в адском пекле, в местечке Торос-Меналла. Вдруг Ахунта Джимдумальбайе нагнулся и присмотрелся к какой-то штуке, торчавшей из земли. Он позвал Брунэ и товарищей, и вскоре все убедились, что Джимдумальбайе нашёл очень важный экземпляр. Окаменелость напоминала череп обезьяны, но у неё были и черты гоминин (см. рис. 24.3). Находку быстро извлекли, пропитали консервантами и отнесли в лагерь.
 Рис. 24.3.
Рис. 24.3. Тумай, череп Sahelanthropus tchadensis (из работы
Michel Brunet et al. A New Hominid from the Upper Miocene of Chad, Central Africa //
Nature. — 2002. — July 11; предоставлено Nature Publishing Group)
Хотя Брюнэ не завершил анализ образца, доставленного в Университет Пуатье, слухи о нём уже распространились. Все спорили о том, что это за находка, гадая по немногочисленным утечкам от людей, которые видели зарисовки или слышали о черепе. Брюнэ ничего не оставалось, кроме как опубликовать предварительный анализ, пока не разлетелась ложная информация. 11 июля 2002 года его работа появилась в качестве главной статьи в авторитетнейшем научном журнале Nature. Брюне назвал свой экземпляр
Sahelanthropus tchadensis в честь региона Сахель на территории Чада, где его нашли, и самой республики Чад. Однако между собой исследователи прозвали находку Тумай, что переводится с чадского языка дазанга как «надежда жизни».
В настоящее время известен только череп сахелянтропа — ни челюсти, ни какой-либо другой части скелета. Кроме того, череп был сильно сдавлен и скошен по диагонали, поэтому выглядит асимметричным и неестественным. Инженеры и компьютерщики использовали специальную моделирующую программу, чтобы восстановить форму черепа и отобразить его истинную форму — прежде чем он оказался раздавлен и засыпан. Окаменелость сравнима с черепом шимпанзе, поэтому сахелянтроп должен был иметь размер примерно как эта обезьяна. Объём его черепа составляет от 320 до 380 куб. см (для сравнения: объём черепа современного человека — более 1350 куб. см). У него также имелись большие надбровные дуги, как у многих примитивных гоминин и человекообразных обезьян. Сахелянтроп обладал рядом других обезьяньих черт, в частности относительно примитивными зубами-молярами.
Но, как указывали Брюнэ и его коллеги, некоторые черты сахелянтропа определённо сближают его с гомининами, а не с шимпанзе или другими человекообразными обезьянами. У него, в отличие от любых человекообразных обезьян, было плоское лицо, почти не похожее на вытянутую морду. Клыки у сахелянтропа небольшие, не то что у человекообразных обезьян (хотя Тумай, видимо, — череп самца, а у большинства самцов человекообразных обезьян большие клыки). Таким образом, зубы сахелянтропа упорядочены на нёбе по контуру, напоминающему букву С, а не по продолговатому контуру вроде буквы U, как у большинства человекообразных обезьян. Наиболее важно, что большое затылочное отверстие в задней части черепа — через него проходит позвоночный столб и соединяется с головным мозгом — находится прямо под основанием черепа, а не скошено ближе к затылку. Таким образом, череп сидел на позвоночнике ровно, а не свешивался вперёд, как у шимпанзе и других человекообразных обезьян.
Последний момент очень важен. Как мы убедимся в 25-й главе, на протяжении всего XX века большинство антропологов склонялись к тому, что объём мозга — важнейший фактор, повлиявший на человеческую эволюцию, а другие признаки, например прямохождение, сформировались позже. Большая часть гоминин, чьи окаменелости найдены в течение последних 30 лет, от Люси до ардипитека и оррорина, явно были полностью прямоходящими животными, но имели небольшой мозг. Сахелянтроп — древнейший ископаемый представитель гоминин, известный на настоящий момент, и факты свидетельствуют, что его череп располагался прямо над позвоночником. Прямохождение — одна из первых адаптаций в эволюции человека, возникшая задолго до того, как наш мозг достаточно вырос.
Этот факт — наряду с плоским лицом, небольшими клыками и характерной для гоминин формой челюсти — позволяет поставить сахелянтропа к человеку ближе, чем любую человекообразную обезьяну. Хотя новые открытия совершаются постоянно, пока Тумай держит рекорд и остаётся древнейшим представителем гоминин. Он жил 6–7 млн лет назад; именно в этот период должны были разойтись эволюционные линии человека и шимпанзе, о чём молекулярные биологи говорят уже 40 лет.
Лучше один раз увидеть!
Подлинные окаменелости сахелянтропа, оррорина, ардипитека, австралопитека и других древнейших гоминин хранятся в специальных закрытых запасниках музеев стран, на территории которых они были найдены (в основном в Эфиопии, Кении, Танзании и Чаде). Лишь профессиональным исследователям разрешается осматривать такие коллекции и прикасаться к редчайшим сокровищам.
Во многих музеях есть экспозиции, посвящённые эволюции человека, где находятся превосходные слепки важнейших окаменелостей. В США следует назвать Американский музей естественной истории (Нью-Йорк), Музей естественной истории под открытым небом (Чикаго), Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия), Музей естественной истории округа Лос-Анджелес (Лос-Анджелес), Музей человека в Сан-Диего, Музей естественной истории им. Пибоди при Йельском университете (Нью-Хейвен, штат Коннектикут). В Европе такие экспонаты есть в Музее естественной истории (Лондон) и в Музее эволюции человека (Бургос, Испания). Из более дальних музеев следует упомянуть Австралийский музей в Сиднее.
Дополнительная литература
Даймонд Дж. Третий шимпанзе. Два процента генотипа, которые решили всё. — М.: ACT, 2013.
Gibbons, Ann. The First Human: The Race to Discover Our Earliest Ancestors. — New York: Anchor, 2007.
Huxley, Thomas H. Evidence as to Man’s Place in Nature. — London: Williams & Norgate, 1863.
Klein, Richard G. The Human Career: Human Cultural and Biological Origins. 3rd ed. — Chicago: University of Chicago Press, 2009.
Marks, Jonathan. What It Means to Be 98% Chimpanzee: Apes, People, and Their Genes. — Berkeley: University of California Press, 2003.
Sponheimer, Matt, Julia A. Lee-Thorp, Kaye E. Reed, and Peter S. Ungar, eds. Early Hominin Paleoecology. — Boulder: University of Colorado Press, 2013.
Tattersall, Ian. The Fossil Trail: How We Know What We Think We Know About Human Evolution. — New York: Oxford University Press, 2008.
Tattersall, Ian. Masters of the Planet: The Search for Our Human Origins. — New York: Palgrave Macmillan, 2013.
Wade, Nicholas. Before the Dawn: Recovering the Lost History of Our Ancestors. — New York: Penguin, 2007.
25. Люси в небе с алмазами
Древнейший человеческий скелет
Австралопитек афарский
Мы должны, однако, мне кажется, признать, что человек, со всеми его благородными качествами... всё ещё носит на своей телесной организации неизгладимую печать низкого происхождения.
Чарльз Дарвин. «Происхождение человека и половой отбор»
Происхождение человека
В книге «Происхождение видов», опубликованной в 1859 году, Чарльз Дарвин не затрагивает палеонтологическую летопись эволюции человека. Даже в книге «Происхождение человека», вышедшей в 1871-м, он не упоминает человеческие окаменелости. У него были веские причины для такого молчания: в середине XIX века имелось лишь несколько находок, позволявших задуматься о существовании доисторических людей. Первых хорошо описанных неандертальцев нашли в 1856 году в известняковом карьере долины Неандер близ Дюссельдорфа (Германия) — всего за три года до публикации «Происхождения видов».
Среди этих окаменелостей был лишь свод черепа и несколько костей конечностей, изначально ошибочно принятых за кости пещерного медведя. Позже распространилась другая неверная интерпретация: якобы кости принадлежали больному кавалеристу-казаку. Выдвигались и более причудливые ложные версии. Все полагали, что это кости современного человека, пусть и необычные. Так вышло, что первый полный скелет неандертальца, найденный в коммуне Ла-Шапель-о-Сен на территории Франции, принадлежал больному старику, страдавшему рахитом. Поэтому на первых реконструкциях неандертальцев изображали сутулыми и нескладными, а не стройными и атлетичными, какими в действительности были эти люди. Правда раскрылась позднее, благодаря многим скелетам, сохранившимся лучше.
Лишь в самом конце XIX века открыли останки ещё более примитивных людей, чем неандерталец. Голландский врач и анатом Эжен Дюбуа увлекался идеями Дарвина и был убеждён, что первые люди сформировались в Восточной Азии. Поэтому в 1887 году он завербовался в голландскую армию в качестве хирурга, чтобы отправиться в Голландскую Ост-Индию (современная Индонезия). И действительно, ему повезло после недолгих раскопок обнаружить настоящее сокровище. Оказалось, что в этом регионе встречались ископаемые останки гоминин. Между 1891 и 1895 годами Дюбуа и его яванская команда обнаружили несколько экземпляров, в том числе свод черепа, бедренную кость и несколько зубов (см. рис. 25.1). Дюбуа назвал открытый им вид
Pithecanthropus erectus (в переводе с греческого «обезьяночеловек выпрямленный»). Хотя окаменелости и оказались фрагментарными, бедренная кость не оставляла сомнений, что её обладатель был прямоходящим. Свод черепа очень примитивный, с ярко выраженными надбровными дугами; объём черепа примерно вдвое меньше, чем у современных людей.
 Рис. 25.1.
Рис. 25.1. Три окаменелости Яванского человека, зарисованные Эженом Дюбуа: свод черепа, коренной зуб, бедренная кость. Каждая окаменелость показана с двух сторон (иллюстрации из фонда Wikimedia Commons)
Дюбуа вернулся в Голландию и в 1899 году получил должность профессора. К сожалению, он плохо переносил нормальную жёсткую критику, обычную в научном сообществе. Многие антропологи скептически отнеслись к заявлениям Дюбуа, основанным на скудном материале, полагали, что эти окаменелости — деформированные останки обезьяны. В результате Дюбуа разозлился, обиделся и перестал участвовать в дебатах. Он спрятал свои находки подальше, отказался кому-либо их показывать и вступать в научную дискуссию. К 1920-м годам у него появились единомышленники, но он по-прежнему жил отверженным и оскорблённым, а в 1940 году скончался в возрасте 82 лет.
Из Евразии?
В 1871 году Дарвин предполагал, что корни человечества следует искать в Африке. Он рассуждал просто: там обитают все наши ближайшие родичи-приматы (шимпанзе и гориллы), поэтому логично, что общий предок обезьян и людей должен был появиться в Африке. Впоследствии большинство антропологов отвергали версию Дарвина и настаивали, что люди появились в Евразии. Приводились разные доводы, в том числе упоминались открытия Дюбуа, сделанные на Яве, а вообще эти взгляды были глубоко расистскими: африканские аборигены считались недочеловеками, представителями иного вида, чем европейцы. Идея о том, что все люди произошли от чёрных африканцев, в начале XX века казалась отвратительной многим европейским учёным.
Почти все антропологи и палеонтологи начала XX века считали, что прародиной человечества являлась Евразия. Знаменитый палеонтолог Генри Фэрфилд Осборн, директор Американского музея естественной истории, в 1920-е годы организовал и помог профинансировать легендарные центральноазиатские экспедиции в Монголию, которыми руководил Рой Чепмен Эндрюс: предполагалось, что там удастся найти древнейших ископаемых предков человека (см. главу 23). Таких окаменелостей не нашли, но открыли очень важные окаменелости динозавров (в том числе их первые яйца и гнёзда), а также по-настоящему интересных и необычных ископаемых млекопитающих. Открытия Эжена Дюбуа, совершённые на Яве, лишь подкрепили «азиатскую» версию.
В 1921 году, когда американская экспедиция приступила к исследованиям в Монголии, шведский палеонтолог Йоханн Гуннар Андерссон обнаружил близ Пекина пещеру Чжоукоудянь. Начатые им раскопки продолжил австрийский палеонтолог Отто Жданский, открыв удивительных представителей фауны ледникового периода, в том числе гигантскую гиену. Также он нашёл два зуба, принадлежавших гоминину. Жданский передал экземпляры канадскому анатому Дэвидсону Блэку, работавшему в Пекинском объединённом медицинском колледже. Блэк опубликовал информацию о них в 1927 году, назвав ископаемое существо
Sinanthropus pekingensis («китайский человек из Пекина»). Этот вид стал широко известен под названием Пекинский человек.
Когда удалось получить финансирование, раскопки продолжились, но за много лет работы удалось найти лишь ещё несколько зубов. Наконец, в 1928 году рабочие добыли нижнюю челюсть, фрагменты черепа и множество зубов, что позволило убедиться в примитивности вида. Благодаря дополнительным вложениям начали более масштабные раскопки, в которых участвовали преимущественно китайские рабочие и учёные. Вскоре нашли более 200 человеческих окаменелостей, в том числе шесть почти полных черепов (см. рис. 25.2). Блэк умер в 1934 году от сердечной недостаточности, а годом позже немецкий анатом Франц Вейденрейх продолжил начатые им исследования, в том числе описание окаменелостей. Хотя Блэк публиковал много предварительных описаний, по мере появления новых ископаемых находок, именно в подробных монографиях Вейденрейха они полностью документированы. При наличии такого материала вскоре стало ясно, что Пекинский человек очень похож на Яванского. Большинство антропологов причисляют их к одному и тому же виду
Homo erectus.
 Рис. 25.2.
Рис. 25.2. Один из наиболее полных черепов Пекинского человека, найденный в китайском Чжокоудяне (из фонда Wikimedia Commons)
Раскопки в Чжоукоудяне продолжались, а над миром сгущались тучи войны. Японская империя начала расширяться, аннексируя новые китайские территории. В 1931 году японцы вторглись в Маньчжурию — регион на северо-востоке Китая, превратив его в свою провинцию Маньчжоу-Го. Там Япония сформировала марионеточное правительство, во главе которого поставили Пу И, последнего китайского императора. В 1937 году началось второе японское вторжение в Китай, в ходе которого японцы захватили ещё один большой китайский регион, сражаясь с националистами Чан Кайши и коммунистами Мао Цзэдуна.
Позднее, в 1941 году, незадолго до нападения на Пёрл-Харбор, встревоженные учёные в Пекине чувствовали, что война близко. Палеонтологи, работавшие в Чжоукоудяне, опасались, что окаменелости попадут в руки японцев и станут трофеями, а не материалом для научных исследований. Все экземпляры, собранные в Пекинском объединённом медицинском колледже, уложили в два больших ящика, которые пытались доставить на грузовике на борт корабля ВМФ США и тайно вывезти из страны через порт Циндао. В условиях конспирации, призванной запутать японских оккупантов, ящики были утеряны. Есть версия, что их погрузили на корабль, который потопили японцы. По другой — ящики просто хорошо спрятали, и никто не знает, где они. Сторонники третьей версии считают, что ящики нашли китайские дельцы, часто уничтожавшие окаменелости («драконьи кости»), перетирая их в порошок и сбывая как средства традиционной китайской медицины. К счастью, почти со всего оригинального материала были сделаны точные слепки, которые сегодня находятся во многих музеях. Поэтому мы знаем, как они выглядели, вплоть до мельчайших деталей. Кроме того, во время следующих раскопок появились новые находки, и потеря не стала невосполнимой.
Из Британии?
Идею о том, что прародина человечества находится в Азии, впервые высказал знаменитый немецкий эмбриолог и биолог Эрнст Геккель, напористо отстаивавший эту точку зрения (задолго до того, как её удалось проверить на материале окаменелостей). Хотя Геккель был крупнейшим протеже Дарвина в Германии, он не соглашался с англичанином в том, что люди возникли и начали развиваться в Африке. Именно Геккель вдохновил Дюбуа, который, казалось бы, подтвердил его взгляды, найдя человеческие окаменелости на Яве. С Геккелем соглашались и другие учёные-первопроходцы, занимавшиеся палеонтологией человека, причём не только те, кто работал в Чжоукоудяне — Андерссон, Жданский, Блэк, Вейденрейх, — но и Осборн и его коллеги из Американского музея естественной истории. Это были Уолтер Грейнджер (крупный учёный, участвовавший в центральноазиатских экспедициях), Уильям Диллер Мэтью (считавший, что большинство отрядов млекопитающих возникли в Евразии, а оттуда распространились по всему миру) и Уильям Кинг Грегори.
На тот момент палеонтологическая летопись, казалось, подтверждала версию о евразийском происхождении человека: сначала были найдены неандертальцы, затем Яванский и Пекинский человек. Удивительно, что ещё одно открытие сделали в Англии, и оно тоже заставляло поверить, что именно Евразия являлась главным центром эволюции человека. В 1912 году на заседании Лондонского геологического общества коллекционер-любитель Чарльз Доусон заявил, что четырьмя годами ранее один рабочий передал ему фрагмент черепа, найденный в гравийном карьере близ Пилтдауна. Рабочий принял находку за окаменевшую кокосовую скорлупу и попытался её расколоть, а Доусон отправился в Пилтдаун и нашёл там новые фрагменты. Он показал их Артуру Смиту Вудварду из Британского музея естественной истории, и уже оба поехали в Пилтдаун. Доусон якобы добыл там новые фрагменты челюсти и свода черепа, Вудвард ничего не нашёл.
Вскоре Вудвард создал реконструкцию челюсти и черепа, основываясь на немногочисленных фрагментах, которые у него имелись (см. рис. 5.3). Экземпляры получились весьма любопытными. Голова была почти как у современного человека, с объёмистой черепной коробкой и небольшими надбровными дугами. Однако челюсть сильно напоминала обезьянью. Нужно отметить, что основание челюсти, а также лицо и многие фрагменты черепа были изломаны или не найдены, поэтому сложно сказать, правильно ли челюсть прикреплена к черепу. В августе 1913 года Вудвард, Доусон и французский священник и палеонтолог Пьер Тейяр де Шарден снова отправились на отвалы пустой породы в Пилтдауне, где Тейяр нашёл клык, прекрасно входивший в зазор между сломанными фрагментами челюсти. Клык был небольшой и напоминал человеческий, не походил на огромные клыки человекообразных обезьян.
 Рис. 25.3.
Рис. 25.3. Череп Пилтдаунского человека, реконструированный Артуром Смитом Вудвардом (из фонда Wikimedia Commons)
Открытие Доусона и реконструкция Вудварда вызвали критику (рис. 25.4). Анатом Артур Кит оспорил эту реконструкцию и сделал свою, более человекоподобную. Анатом Дэвид Уотерстон из лондонского Кингс-колледжа заключил, что две окаменелости не подходят друг к другу, и Пилтдаунский человек — просто человеческий череп, к которому прикрепили челюсть обезьяны. Такого же мнения придерживались французский палеонтолог Марселин Буль, описавший неандертальцев из Ла-Шапель-о-Сен, и американский зоолог Геррит Смит
Миллер. В 1923 году Франц Вейденрейх, описавший Пекинского человека, категорично заявил, что Пилтдаунский человек — это человеческий череп с обезьяньей челюстью, причём его зубы подпилены, чтобы уменьшить сходство с обезьяной.
 Рис. 25.4.
Рис. 25.4. Картина Джона Кука «Дискуссия о пилтдаунском черепе». На этой знаменитой картине изображены члены Британского антропологического общества, изучающие экземпляр Пилтдаунского человека. В первом ряду, в центре сидит сэр Артур Кит (главный сторонник подлинности экземпляра). В заднем ряду, слева направо: Ф. О. Барлоу, Графтон Эллиот Смит, Чарльз Доусон (автор мистификации) и Артур Смит Вудвард (главный консультант по геологии в Музее естественной истории, официально описавший экземпляр). Слева в переднем ряду: А. С. Андервуд, справа в переднем ряду: У. П. Пайкрафт и знаменитый анатом Рэй Ланкестер (иллюстрация из фонда Wikimedia Commons)
Хотя в 1920-е и 1930-е годы находились критики и скептики, сомневавшиеся в реальности Пилтдаунского человека, титаны британской палеонтологии, особенно Вудвард, Кейт и Графтон Эллиот Смит, твёрдо в него верили. Несмотря на проблемы, это ископаемое их удовлетворяло. Во-первых, оно подтверждало, что основным движущим фактором человеческой эволюции было увеличение мозга, начавшееся задолго до того, как наши предки утратили обезьяньи зубы, челюсти и стали прямоходящими. Тогда в палеоантропологии царила догма: первым делом развился крупный человеческий мозг и интеллект, а потом он стал двигателем человеческой эволюции. Во-вторых, сработал шовинизм. Англичане гордились тем, что недостающее звено найдено на их родине, что первый британец оказался ещё примитивнее Яванского и Пекинского человека. Складывалось впечатление, что Европа, особенно британские острова, были центром человеческой эволюции. Окаменелость так удачно соответствовала антропологическим мифам и предубеждениям того времени, что критики вскоре умолкли, и Пилтдаунский человек оставался каноническим экземпляром 41 год.
Лишь в конце 1940-х – начале 1950-х годов он вновь стал вызывать вопросы, поскольку к тому времени этот экземпляр уже не вписывался в палеонтологическую летопись, которая постоянно дополнялась. Особенно много находок поступало из Африки. Десятилетиями образцы Пилтдаунского человека хранились под замком, изучать можно было лишь сделанные с них слепки, поэтому немногим довелось рассматривать сами окаменелости. Только в 1953 году химик Кеннет Оукли, антрополог Уилфрид Э. Ле Гро Кларк и Джозеф Уэйнер исследовали оригинальные образцы. Они подтвердили, что Пилтдаунский человек — подделка: череп, принадлежавший современному человеку, извлекли из средневековой могилы, челюсть взяли у орангутана, зубы — у шимпанзе. Все экземпляры были искусственно состарены. Для этого их держали в растворе железной и хромовой кислоты. Кроме того, зубы были намеренно спилены, чтобы меньше походили на обезьяньи (как правильно догадывался Вейденрейх).
Обо всех лицах, причастных к этому заговору, говорят до сих пор. Чарльз Доусон «нашёл» все окаменелости, но выяснилось, что ранее он уже подделывал разные артефакты и человеческие окаменелости, поэтому мог быть единственным виновником. Некоторые полагали, что в одиночку, без экспертных консультаций анатома или антрополога, Доусон не смог бы изготовить столь реалистичную подделку и продуманно уничтожить все фрагменты, которые могли его выдать, и что череп и челюсть не связаны друг с другом. В разное время учёные предполагали, что к делу причастны Тейяр де Шарден, Артур Кит, зоолог Мартин Хинтон, мистификатор и поэт Гораций де Вир Коул и даже Артур Конан Дойл, прославившийся рассказами о Шерлоке Холмсе. С тех пор прошло более ста лет, но решающие доказательства так и не найдены. Мы лишь знаем, что Доусон был основным (и, возможно, единственным) мистификатором, который задолго до этого занимался подобными делами. Возможно, мы никогда не узнаем, помогал ли ему кто-нибудь.
Ребёнок из Таунга и миссис Плез
В то время как евразийская гипотеза происхождения человечества исследовалась в музеях и университетах Европы, Африка оставалась задворками научного мира. Большинство африканских городов были сонными колониальными местечками, без крупных университетов и музеев. Европейские учёные, особенно англичане, немцы, французы, португальцы и голландцы, приезжали в африканские колонии, забирали оттуда важные экземпляры для своих музеев, но ничего не оставляли местным жителям, которых считали грубыми колонизаторами или невежественными дикарями.
Одним из немногих исключений являлась Южная Африка. Поскольку морское сообщение вокруг южной оконечности Африки было стратегически важно, а также из-за исключительного богатства этого региона (здесь добывали золото, алмазы, драгоценные металлы), страна была заселена и европеизирована на века раньше остального континента. В результате там активнее строили современное государство по европейскому образцу. Этим занимались как британские колонизаторы, так и голландские поселенцы (буры). Кейптаун, Йоханнесбург, Дурбан, Претория, другие города были большими и развитыми. Они могли похвастаться собственными университетами и музеями, одними из немногих в Африке. Кроме того, Южная Африка стала вторым независимым государством на континенте: она потеряла статус колонии в 1910 году, тогда как большинство иных африканских территорий получили независимость лишь в 1950-е – 1960-е годы.
Среди работавших в Южной Африке учёных, имевших европейское образование, был молодой австралиец Рэймонд Дарт. Он получил медицинскую степень в Университетском колледже Лондона, а затем перебрался в Южную Африку, где стал работать на недавно образованном факультете анатомии в Витсватерсрандском университете Йоханнесбурга. По прибытии он с досадой убедился, что на факультете нет сравнительных коллекций обезьяньих и человеческих черепов, столь важной для изучения анатомии, и объявил среди студентов соревнование: кто принесёт в университет самые интересные кости. Одна девушка — единственная на всем курсе — сказала, что видела череп ископаемого павиана на каминной полке в доме у приятеля. Дарт усомнился, что это может быть павиан, поскольку в Африке южнее Сахары почти не находили ископаемых приматов. Однако, увидев череп, учёный понял, что студентка права. Оказалось, что череп лежал дома у директора компании Northern Lime, изготавливавшей цемент из известняка, который добывали в карьере под названием Таунг. Дарт попросил приносить ему любые другие окаменелости, которые обнаружат рабочие, взрывающие известняковые пустоты.
Однажды летним утром 1924 года Дарт отчаянно пытался застегнуть галстук-бабочку — приятель пригласил его на свадьбу шафером, а саму свадьбу собирались играть прямо в доме у Дарта. Вдруг он услышал шум: у двери поставили два деревянных ящика. Дарт решил сразу в них заглянуть. В первом не было ничего интересного, а приоткрыв второй, учёный обнаружил там прекрасную окаменелость черепной коробки с отчётливым отпечатком эндокрана. Он стал взволнованно рыться в ящике, чтобы найти лицо от этого черепа. Дарт уже определил, что хотя череп существа примерно как у шимпанзе, мозг гораздо больше. Тем временем друг-жених уговаривал его закончить туалет. Всю церемонию Дарту не терпелось вернуться к своим «сокровищам». В течение нескольких следующих месяцев он очищал и препарировал экземпляр. И наконец, очередным точным ударом (Дарт работал молотком и вязальными спицами) отколол твёрдый слой брекчии от лицевой поверхности.
Оказалось, что это лицо ребёнка, которому было примерно четыре года — у него даже сохранились все молочные зубы (рис. 25.5). Хотя череп примерно такой же, как у современного шимпанзе, у него есть ряд черт, свойственных гомининам: необычно крупный мозг, плоское лицо (без удлинённой морды), маленькие надбровные дуги, уменьшенные клыки; зубы ребёнка расположены на нёбе полукругом. Важнее всего, что большое затылочное отверстие в основании черепа, к которому подходит позвоночник, находится как раз под основанием черепа — таким образом, это существо держало голову прямо и, вероятно, ходило на двух ногах.
 Рис. 25.5.
Рис. 25.5. Череп австралопитека африканского, более известный под названием Ребёнок из Таунга. Вид сбоку (снимок из фонда Wikimedia Commons)
Дарт подготовил описание, анализ и в 1925 году опубликовал эти материалы в Nature — ведущем научном журнале мира. Он назвал экземпляр
Australopithecus africanus (от греческого «африканская южная обезьяна»). Находка явно свидетельствовала, что первые гоминины обитали в Африке, тем более что это был самый обезьяноподобный и примитивный гоминин, найденный на тот момент. Поскольку Дарт изучал эндокраниальные слепки мозга в медицинской школе, его особенно заинтересовала внутренняя поверхность черепа этого образца. Мозг австралопитека был крупнее, чем у любой обезьяны, хотя умещался в обезьяньем черепе. Более того, австралопитек явно имел высокоразвитую лобную долю мозга — напоминавшую скорее человеческую, чем обезьянью.
Дарт считал свои доказательства бесспорными и рассчитывал на хвалебные отзывы со стороны научного сообщества. Как же он был разочарован, когда все великие европейские антропологи раскритиковали его находку как обычного «детёныша обезьяны». Отчасти проблема заключается в том, что юные обезьянки действительно похожи на людей гораздо сильнее, чем взрослые особи. Однако прямую осанку, человеческие коренные зубы, уменьшенные клыки и полукруглое расположение зубов на нёбе, а также увеличенную лобную долю мозга нельзя было объяснить юным возрастом. Это неприемлемо. Артур Кит, один из главных творцов Пилтдаунского человека, писал: «Заявления (Дарта) нелепы, этот череп принадлежит молодой человекообразной обезьяне... и по столь разнообразным признакам сходен с черепами двух ныне живущих африканских антропоидов, горилл и шимпанзе, что мы можем без малейших колебаний отнести данную ископаемую форму к одной из этих современных групп».
Кроме того, существовали негласные моменты: империализм и расизм. Крупнейшие европейские учёные не доверяли выводам Дарта — невежественного южноафриканского анатома, хоть он и учился в Лондоне; не признанного эксперта, а «сельского пентюха». Его статья в Nature вышла очень краткой (таково требование редакции), поэтому ведущие антропологи могли судить об образце лишь по сжатой характеристике и нескольким крошечным иллюстрациям, сделанным от руки. Позже Дарт подготовил более подробное описание, уделив особое внимание мозгу. Но ни у кого не нашлось времени, денег или настроения отправиться в долгое морское путешествие, побывать в Южной Африке и самостоятельно ознакомиться с образцами.
Поэтому Дарт сам их привёз. В 1931 году он побывал в Англии, взяв с собой череп Ребёнка из Таунга, но тщетно. Британские антропологи не могли расстаться с расистскими предрассудками. Кроме того, в то время вся Европа была в состоянии эйфории от открытия Пекинского человека, как раз публиковались иллюстрации и описания Дэвидсона Блэка. Они подкрепляли версию евразийского происхождения — наряду с найденными ранее Яванским и Пилтдаунским человеком. Поэтому никто не заметил ни несчастного Ребёнка из Таунга, единственный африканский экземпляр, ни самого Дарта.
Дарту пришлось ждать ещё 20 лет, прежде чем европейское научное сообщество перестало от него отмахиваться и коллеги начали осознавать важность его находки. В 1947 году Кит признал: «Дарт был прав, а я ошибался». Но Дарт хорошо посмеялся последним. Он умер в 1988 году в возрасте 95 лет, уважаемый и прославленный благодаря своим открытиям, а также первопроходческому вкладу в современную палеоантропологию. Большинство его убеждённых соперников умерли гораздо раньше и сегодня уже забыты.
Но и в 1920–1930-е годы южноафриканские учёные были уверены, что Дарт прав и его критикуют несправедливо. Среди этих учёных был шотландец, доктор Роберт Брум (см. главу 19), который уже сделал себе имя в палеонтологии, открыв замечательные экземпляры пермских рептилий, а также древнейших предков млекопитающих в пустыне Кару. Некоторые собиратели окаменелостей, с которыми он сотрудничал, присылали ему ископаемые образцы, найденные в многочисленных южноафриканских карстовых пещерах. В 1938 году, работая в пещере Кромдрай, Брум нашёл очень массивный череп взрослого примата, назвал его
Paranthropus robustus (от греческого «массивный получеловек»). Позднее, в знаменитой системе пещер Сварткранса он обнаружил окаменелости, принадлежавшие более чем 130 особям
P. robustus. Недавний анализ их зубов показал, что среди этих массивных гориллоподобных людей нет ни одной особи старше 17 лет и что небогатый рацион робустусов состоял из орехов, семян и травы.
В том же 1938 году Брум получил слепок эндокрана ископаемого черепа объёмом около 485 куб. см — слишком большого для обезьяны. Он назвал этот экземпляр
Plesianthropus transvaalensis (от греческого «полуобезьяна из Трансвааля»). Затем до него дошли слухи об окаменелостях, найденных в пещере Стеркфонтейн. 18 апреля 1947 года Брум и Джон Т. Робинсон нашли полностью сохранившийся череп, принадлежавший, вероятно, взрослой самке (сейчас считается, что это был самец). Она имела явно человекоподобные черты, но была столь же примитивна, как Ребёнок из Таунга (рис. 25.6). Учёные назвали этот экземпляр миссис Плез. Их открытие продемонстрировало, что в Южной Африке встречаются окаменелости гораздо более примитивных гоминин, чем где-либо в Евразии. Вскоре в Стеркфонтейне были найдены и другие окаменелости, позволившие оценить изменчивость особей в популяции
Plesianthropus transvaalensis. В дальнейшем антропологи решили, что взрослые особи из Стеркфонтейна и Ребёнок из Таунга относились к одному и тому же виду, поэтому название
Plesianthropus transvaalensis теперь отменено в пользу оригинального таксона
Australopithecus africanus, предложенного Дартом.
 Рис. 25.6.
Рис. 25.6. В нескольких ракурсах показан череп австралопитека африканского, известный под названием миссис Плез (снимок из фонда Wikimedia Commons)
Эти открытия в Африке, наряду с полным отсутствием столь примитивных и древних окаменелостей в Евразии, заставили антропологов отказаться от азиатской версии человека. К 1947 году уже были описаны разные австралопитеки, и становилось всё очевиднее, что Дарвин был прав: люди возникли в Африке. В то же время отмирала идея, что основным движущим фактором человеческой эволюции было увеличение мозга и рост интеллекта, а другие признаки — мелкие зубы и прямохождение — возникли позднее. Вместе с ней вымирали и фашиствующие антропологи старой закваски. Все окаменелости, найденные до сих пор, демонстрируют, что сначала стала развиваться прямая осанка и сложные зубы, а мозг начал увеличиваться гораздо позднее. Поэтому в 1953 году вновь решили подробно изучить Пилтдаунского человека: он больше не вписывался в картину человеческой эволюции. Эта мистификация была разоблачена со смешанным чувством конфуза и облегчения.
Удача Лики
Ещё один сторонник африканской гипотезы — легендарный Луис С. Б. Лики (рис. 25.7,
А). По всеобщему признанию, это был харизматичный, энергичный, прямолинейный человек, который мог рассказать настоящую сагу о своих открытиях. Критики считали, что он безответственно и небрежно относится к науке, иногда покупался на сомнительные идеи, которые впоследствии не подтверждались. Тем не менее Лики навечно вписал своё имя в историю изучения эволюции человека — не только как первооткрыватель многих важных окаменелостей, но и как наставник своей жены Мэри, совершившей большинство открытий, сына Ричарда, превзошедшего отца, и многих других видных антропологов. Он также вдохновил приматологов Джейн Гудолл, Дайан Фосси и Бируте Галдикас потратить долгие годы на изучение шимпанзе, горилл и орангутанов соответственно.

 Рис. 25.7.
Рис. 25.7. (
А) Луис Б. Лики с окаменелостью в руке; (
Б) череп зинджантропа (современное наименование —
Paranthropus boisei), благодаря которому семья Лики стала всемирно известна, а центр палеоантропологических исследований вновь переместился в Африку ((
А) снимок из фонда Wikimedia Commons; (
Б) снимок предоставлен Национальным музеем Кении)
Лики родился в семье британских миссионеров на территории современной Кении, вырос среди нетронутой природы Восточной Африки, впитал культуру кикуйю — одного из крупнейших народов этого региона, научился бегло говорить на их языке. Хотя молодой человек получил лишь фрагментарное образование от кенийских учителей, после Первой мировой войны его отправили в Кембридж, где он зарекомендовал себя блестящим, старательным, но зачастую эксцентричным студентом. Лики выбрал путь антрополога и уже до тридцати лет опубликовал ряд работ по археологии Кении. В начале 1930-х его научная карьера едва не рухнула, когда он бросил первую жену Фриду, влюбившись в молодую художницу Мэри. Чтобы спрятаться от нападок академического сообщества, они с Мэри вернулись в Кению, где обнаружили несколько мест с окаменелостями примитивных обезьян — в Канаме, Канджере и на острове Русинга.
Живя в Кении в конце 1930-х – начале 1940-х годов, Лики играл важную роль в политической жизни региона, так как знал язык кикуйю и разбирался в культуре туземцев. Он был не только английским разведчиком в годы войны, но и переводчиком, посредником между британцами и кикуйю, когда между теми возникали трения. Также он являлся ключевой фигурой во время восстания мау-мау и в итоге помог разрешить конфликт. При этом Лики отказался вернуться в Европу, согласившись на мизерное жалованье в музее Кориндон в Найроби (в настоящее время — Национальный музей Кении).
При всей авторитетности Лики и важности его африканских находок, ему не удавалось совершить по-настоящему убедительное открытие, которое не только подтвердило бы, что человечество возникло в Африке, но и обеспечило бы карьерный рост, помогло бы добыть дополнительное финансирование для исследований. При всей важности экземпляров из южноафриканских пещер, их не удавалось датировать. Требовалось найти место, где человеческие окаменелости встречались бы вместе с останками других млекопитающих, период жизни которых был хорошо известен, либо с вулканическим пеплом, обеспечивающим точную датировку.
В 1913 году немецкий археолог Ганс Рек извлёк скелет вполне современного человека из пласта II в Олдувайском ущелье (территория современной Танзании). Его находка была неоднозначной, так как, судя по всему, относилась к среднему плейстоцену и была гораздо старше европейских человеческих окаменелостей, находившихся на том же уровне эволюции. В 1931 году к дискуссии подключился Лики, сумевший убедить коллег, что экземпляр Река — не современный человек, погребённый в древних породах. В 1951 году, когда отгремела Вторая мировая война, наладилась послевоенная политика и освободилось время для занятий антропологией, Луис и Мэри стали не покладая рук работать в самых нижних слоях Олдувайского ущелья. Там они нашли множество каменных орудий, свидетельствующих о существовании гораздо более примитивной культуры, нежели европейская, но не добыли убедительных окаменелостей.
Затем, в 1959 году, спустя восемь лет упорного труда (и через тридцать лет после того, как Луис приступил к раскопкам в Олдувае), Мэри нашла превосходный ископаемый череп (см. рис. 25.7,
Б). Он был гораздо более примитивным и грубым, чем любые подобные окаменелости, найденные в Южной Африке или где-либо ещё. Череп добыли в пласте I, самом нижнем слое отложений Олдувайского ущелья. Лики назвали его «дорогой мальчик». Черепу присвоили официальное название
Zinjanthropus boisei, или просто Зиндж (для краткости): родовое название дано по арабскому названию Восточной Африки, видовое — в честь Чарльза Бойса, профинансировавшего работы. Затем, в 1960 году, Джек Эвернден и Гарнисс Кёртис датировали слой пепла, расположенный под олдувайскими отложениями I, с помощью только что разработанного калий-аргонового метода и выяснили, что этим пластам 1,75 млн лет — гораздо больше, чем кто-либо мог предположить. На тот момент большинство учёных полагали, что весь плейстоцен продолжался не более нескольких сотен тысяч лет, но в 1960-е годы хронологическую шкалу перекалибровали на основании калий-аргонового метода. Вскоре Лики стали всемирно знамениты, получили поддержку и финансирование от Национального географического общества. Более того, в поисках новых экземпляров в Африку хлынули антропологи — ведь стало ясно, что эволюция человека протекала, по большей части, именно там, а в Евразию и на другие континенты люди мигрировали значительно позднее.
Наследие Люси
Вскоре лихорадочные поиски гоминин с «чёрного континента» уже велись по всей Восточной Африке, особенно в тех регионах, где имелись древние осадочные напластования, в сбросовых долинах вдоль Великого рифта. Ричард, сын Луиса Лики, поначалу не интересовался антропологией, но впоследствии продолжил дело отца. Он стремился выйти из отцовской тени, поэтому сам приступил к раскопкам на озере Рудольф (в настоящее время называется Туркана) на севере Кении. Там было найдено множество новых черепов, в том числе наиболее хорошо сохранившийся экземпляр
Homo habilis, древнейшего представителя нашего рода
Homo. Ричард сделал себе имя в кенийской политике, особенно как борец против хищнической охоты на носорогов и слонов. Его жена Мив, работая с местными жителями, продолжила дело Лики. Мэри, мать Ричарда, продолжала находить важные окаменелости и, в частности, обнаружила знаменитую цепочку человеческих следов в местечке Летоли в Танзании.
Кения и Танзания почти ежегодно упоминались в новостях, всякий раз в связи с удивительными находками Лики. В конце 1960-х Луиса Лики пригласили на обед, где также присутствовали президент Кении Джомо Кениата и император Эфиопии Хайле Селассие. Император спросил Лики, почему никаких открытий не сделано в Эфиопии. Луис быстро его убедил, что окаменелости найдутся, если император даст учёным добро на раскопки. Вскоре антрополог Ф. Кларк Хоуэлл из Беркли стал работать на северном берегу озера Туркана, где река Омо вытекает из Эфиопии. Хоуэлл и его коллега Глинн Айзек много лет собирали материалы в отложениях Омо, которые удобны для датировки благодаря изобилию вулканического пепла. К сожалению, эти залежи образовались при сходе внезапных паводков, от которых остались гравийные и песчаные наносы. В таких породах окаменелости быстро ломаются и истираются, поэтому хорошо сохранившихся останков гоминин там не обнаружили.
Тем временем другие молодые и перспективные антропологи желали совершать собственные открытия в регионе, который фактически оставался вотчиной Лики и их соратников. Двумя представителями нового поколения стали Дональд Джохансон и Тим Уайт. Оба хотели попытать профессиональную удачу в местах, не контролируемых семьёй Лики. От французских геологов Мориса Тайеба и Ива Коппена, а также антрополога Джона Кальба они узнали об отложениях Афарского треугольника — рифтовой долины, раскинувшейся между тектоническими плитами, где Аденский залив соединяется с Красным морем. В этих пластах уже находили многочисленные окаменелости млекопитающих, позволяющие предположить, что здешним отложениям около 3 млн лет. Потенциально эти окаменелости были старше любых ископаемых гоминин, найденных ранее в Кении и Танзании. Джохансон, Уайт, Тайеб и Коппен получили разрешение работать на этих пластах и в 1973 году начали раскопки в Хадаре.
Спустя многие месяцы исследований и разведочных работ в поисках окаменелостей (за это время было найдено несколько фрагментов костей гоминин), 24 ноября 1974 года Джохансон прервал свои полевые заметки и взялся помочь студенту Тому Грею, исследовавшему обнажение пород. Боковым зрением Джохансон заметил поблёскивающую кость, извлёк её и сразу понял, что кость принадлежит гоминину. Они стали откапывать новые кости, пока не добыли почти 40% скелета гоминина (рис. 28.5,
А). Это был первый гоминин старше неандертальцев позднего плейстоцена, известный не по отдельным фрагментам, а по относительно полному скелету. Той ночью они праздновали находку у костра, слушая кассету с песнями Beatles — так началась «Люси в небе с алмазами». Одна из участниц команды, Памела Олдерман, весело напевая, предложила назвать ископаемую находку Люси. Позже гоминин получил официальное название
Australopithecus afarensis в честь Афарского треугольника, где его нашли.

 Рис. 25.8.
Рис. 25.8. Люси, австралопитек афарский: (
А) скелет; (
Б) реконструкция внешнего облика ((
А) снимок предоставлен D. Johanson; (
Б) фотография автора)
Через год после открытия Люси команда вернулась в Хадар, где нашла целую россыпь костей австралопитека афарского. Эта находка, названная Первым семейством, стала первым большим образцом, содержавшим ископаемые останки как молодых, так и взрослых гоминин, добытым из отложений старше 3 млн лет. Антропологи смогли оценить типичную изменчивость особей в одной популяции. Эти данные могут быть важны при определении, относится ли новая окаменелость, слегка отличающаяся от известных ранее, к самостоятельному виду, это представитель нового рода либо просто член той же изменчивой популяции.
Проанализировав Люси, Джохансон и Уайт определили, что скелет принадлежал взрослой самке ростом около 1,1 м (см. рис. 28.5,
Б). Ключевое значение имеют её коленные суставы и тазовые кости, чьи важнейшие признаки свидетельствуют: австралопитек афарский был прямоходящим, его ноги полностью находились под телом, как у современного человека. У него был относительно небольшой мозг (380–430 куб. см) и маленькие клыки, как у более высокоразвитых гоминин. При этом у него ещё ярко выраженная морда, а не плоское лицо. Это был очередной удар по антропологической теории «сначала вырос мозг», которая в середине 1970-х годов ещё считалась авторитетной. Лопатка, руки и кисти похожи на обезьяньи, то есть австралопитек афарский по-прежнему лазал по деревьям, даже будучи полностью прямоходящим. Однако на ступне нет признаков хватательного большого пальца, то есть ноги и ступни австралопитека афарского были целиком приспособлены для ходьбы по земле, а не для того, чтобы цепляться за ветки.
С тех пор как в середине 1970-х годов нашли Люси, палеоантропологи совершили много новых удивительных открытий. Люси была первым древним гоминином (старше 3 млн лет), известным по относительно полному скелету, а не по фрагментам черепа или отдельным костям конечностей. В 1984 году Алан Уолкер и группа Лики нашли на западном берегу озера Туркана Мальчика из Нариокотоме. Этот скелет, которому 1,5 млн лет, сохранился почти на 90% и по сей день является самым полным экземпляром древних гоминин из всех известных. Возможно, он относится к виду
Homo erectus или
H. ergaster (вопрос остаётся открытым). В 1994 году Уайт и его команда нашли в Эфиопии почти полный скелет
Ardipithecus ramidus, которому примерно 4,4 млн лет.
Итак, палеонтологическая летопись гоминин улучшается год за годом, новые экземпляры обнаруживают постоянно. За минувший век мы проделали огромный путь: ранее нам были известны только неандертальцы, Яванский и Пекинский человек, а Пилтдаунский человек воспринимался вполне серьёзно. Сегодня известно шесть родов гоминин, кроме
Homo (
Ardipithecus,
Australopithecus,
Kenyanthropus,
Orrorin,
Paranthropus и
Sahelanthropus), и более 12 валидных видов. Вместо упрощённой идеи о том, что с течением времени развивался один человеческий род, палеонтологическая летопись позволила выявить сложное и ветвистое эволюционное древо — несколько эволюционных линий сосуществовали в одних и тех же местах.
Всего 30 тыс. лет назад на планете остался единственный доминирующий вид человека. Сегодня
Homo sapiens угрожает не только всем остальным видам, но и самому себе — мы можем вымереть, как любые ископаемые виды, описанные в этой книге.
Лучше один раз увидеть!
Подлинные окаменелости ардипитека, австралопитека, человека умелого, человека прямоходящего и других древнейших гоминин хранятся в специальных закрытых запасниках музеев стран, на территории которых они найдены (в основном в Эфиопии, Кении, Танзании и Чаде). Лишь профессиональным исследователям разрешается осматривать такие коллекции и прикасаться к этим редчайшим сокровищам.
Во многих музеях есть экспозиции, посвящённые эволюции человека, где находятся превосходные слепки важнейших окаменелостей. В США следует упомянуть Американский музей естественной истории (Нью-Йорк), Музей естественной истории под открытым небом (Чикаго), Национальный музей естественной истории Смитсоновского института (Вашингтон, округ Колумбия), Музей естественной истории округа Лос-Анджелес (Лос-Анджелес), Музей человека (Сан-Диего), Музей естественной истории им. Пибоди при Йельском университете (Нью-Хейвен, штат Коннектикут). В Европе такие экспонаты есть в Музее естественной истории (Лондон) и в Музее эволюции человека (Бургос, Испания). Из более дальних музеев следует упомянуть Австралийский музей в Сиднее.
Дополнительная литература
Boaz, Noel T., and Russell T. Ciochon. Dragon Bone Hill: An Ice-Age Saga of Homo erectus. — Oxford: Oxford University Press, 2008.
Dart, Raymond A., and Dennis Craig. Adventures with the Missing Link. — New York: Harper, 1959.
Johanson, Donald, and Maitland Edey. Lucy: The Beginnings of Humankind. — New York: Simon & Schuster, 1981.
Johanson, Donald, and Blake Edgar. From Lucy to Language. — New York: Simon & Schuster, 2006.
Kalb, Jon. Adventures in the Bone Trade: The Race to Discover Human Ancestors in Ethiopia’s Afar Depression. — New York: Copernicus, 2000.
Klein, Richard G. The Human Career: Human Cultural and Biological Origins. 3rd ed. — Chicago: University of Chicago Press, 2009.
Leakey, Richard E., and Roger Lewin. Origins: What New Discoveries Reveal About the Emergence of Our Species and Its Possible Future. — New York: Dutton, 1977.
Lewin, Roger. Bones of Contention: Controversies in the Search for Human Origins. — Chicago: University of Chicago Press, 1997.
Lewin, Roger. Human Evolution: An Illustrated Introduction. 5th ed. — New York: Wiley-Blackwell, 2004.
Morell, Virginia. Ancestral Passions: The Leakey Family and the Quest for Humankind’s Beginning. — New York: Touchstone, 1996.
Reader, John. Missing Links: In Search of Human Origins. — Oxford: Oxford University Press, 2011
.
Sponheimer, Matt, Julia A. Lee-Thorp, Kaye E. Reed, and Peter S. Ungar, eds. Early Hominin Paleoecology. — Boulder: University of Colorado Press, 2013.
Swisher, Carl C., III, Garniss H. Curtis, and Roger Lewin. Java Man: How Two Geologists Changed Our Understanding of Human Evolution. — Chicago: University of Chicago Press, 2001.
Tattersall, Ian. The Fossil Trail: How We Know What We Think We Know About Human Evolution. — New York: Oxford University Press, 2008.
Tattersall, Ian. Masters of the Planet: The Search for Our Human Origins. — New York: Palgrave Macmillan, 2013.
Wade, Nicholas. Before the Dawn: Recovering the Lost History of Our Ancestors. — New York: Penguin, 2007.
Walker, Alan, and Pat Shipman. The Wisdom of the Bones: In Search of Human Origins. — New York: Vintage, 1997.
Примечания
1
Согласно последним исследованиям, вода на Марсе есть (см., напр.
http://www.nature.com/ngeo/journal/v8/n11/full/ngeo2546.html). (
Примеч. пер.)
(обратно)
2
trilobos (
греч.) — трёхдольный. (
Примеч. пер.)
(обратно)
3
В России тоже можно отыскать трилобитов. Например, несколько мест указаны в статье журнала «Наука и жизнь» по ссылке
http://www.nkj.ru/archive/articles/18991/. (
Примеч. пер.)
(обратно)
4
30 см (
Примеч. ред.)
(обратно)
5
9,5 кг (
Примеч. ред.)
(обратно)
6
320 км (
Примеч. ред.)
(обратно)
7
2,7 м (
Примеч. ред.)
(обратно)
Оглавление
Предисловие
Благодарности
01. Планета ила
Первые окаменелости
Криптозоон
Дилемма Дарвина
Криптозоон: ещё одна ложная тревога?
Эврика!
Лучше один раз увидеть!
Дополнительная литература
02. Эдиакарский сад
Первые многоклеточные организмы
Чарния
От одной клетки ко многим
Окаменелости хребта Флиндерс
Камо грядеши, Эдиакара?
Лучше один раз увидеть!
Дополнительная литература
03. Мелкие раковинные ископаемые
Первые раковины
Клаудина
Строители раковин
Появление «ракушечек»
Прогнозы Престона Клауда
Клаудина
Тлеющий фитиль
Дополнительная литература
04. Хочу я туда, где трилобитов стада...
Первые крупные панцирные животные
Оленеллус
Посланцы далёких времён
Кто такие трилобиты
Оленеллус и первые трилобиты
Что произошло с трилобитами
Лучше один раз увидеть!
Дополнительная литература
05. Это червь или членистоногое?
Происхождение членистоногих
Галлюцигения
Диковины сланцев Бёрджесс
Галлюцинация в камне
Кто такие членистоногие
Бархатные черви и членистоногие
Макроэволюция от типа к типу
Лучше один раз увидеть!
Дополнительная литература
06. Это червь или моллюск?
Происхождение моллюсков
Пилина
Недостающие звенья найдены
Первые моллюски
Галатея и переворот в биологии
Дополнительная литература
07. Растущие из моря
Происхождение наземных растений
Куксония
Первые наземные растения
Высокие первопроходцы: сосудистые растения
Открытие Изабель Куксон
Планета начинает зеленеть
Лучше один раз увидеть!
Дополнительная литература
08. Ихтиолог на палеорыбалке
Происхождение позвоночных
Хайкоуихтис
Хью Миллер и древний красный песчаник
Эпоха рыб
Удочка в прошлое
Соединяя звенья
Нерыбоподобное звено
Лучше один раз увидеть!
Дополнительная литература
09. Мегачелюсти
Самая крупная рыба
Мегалодон
Прогулка на холм Шарктус
Кишащие акулами миоценовые моря
Вот такая рыба!
Гроза морей
Документальная фантастика
Лучше один раз увидеть!
Дополнительная литература
10. Рыба, вышедшая из воды
Происхождение земноводных
Тиктаалик
Из воды на сушу
Ваша внутренняя рыба
Лучше один раз увидеть!
Дополнительная литература
11. «Лягомандра»
Происхождение лягушек
Геробатрахус
«Человек — свидетель потопа»
Жизнь по обе стороны
Сокровища красноцветных отложений
Когда амфибии правили миром
В поисках «Лягомандры»
Лучше один раз увидеть!
Дополнительная литература
12. Черепаха с половиной панциря
Происхождение черепах
Одонтохелис
Черепахи до самого низа
Промежуточные черепахи
Первая пресноводная черепаха
Черепаха с половиной панциря
Под пирамидой черепах
Лучше один раз увидеть!
Дополнительная литература
13. Ходячие змеи
Происхождение змей
Хаасиофис
Батюшки, живые змеи!
Откуда взялись змеи
Лучше один раз увидеть!
Дополнительная литература
14. Царь рыбоящеров
Крупнейшая морская рептилия
Шонизавр
Она продаёт ракушки на морском берегу
«Рыбоящеры»
Происхождение ихтиозавров
Киторептилии триасового периода
Лучше один раз увидеть!
Дополнительная литература
15. Гроза морей
Величайший морской монстр
Кронозавр
Океаны в глуши
Морское чудовище на краю света
Царь морских чудовищ
Морские длинношеи
Происхождение морских чудовищ
Лох-несское чудовище?
Лучше один раз увидеть!
Дополнительная литература
16. Чудовищный хищник
Крупнейший хищник
Гиганотозавр
Царь тиранических ящеров
Из Африки
Крупнее королевского тираннозавра
Лучше один раз увидеть!
Дополнительная литература
17. Страна гигантов
Крупнейшее сухопутное животное
Аргентинозавр
Образ жизни древних гигантов
Размер имеет значение!
Живой динозавр в Конго?
Лучше один раз увидеть!
Дополнительная литература
18. Перо в камне
Первая птица
Археоптерикс
Творения природы
Как повезло Дарвину
Всё больше экземпляров
Птица... или динозавр?
Птицы взлетают
Лучше один раз увидеть!
Дополнительная литература
19. Почти млекопитающее
Происхождение млекопитающих
Тринаксодон
Протомлекопитающие
Великая Кару
Горгоновы морды, ужасные головы и двойные собачьи зубы
Ухочелюсти
Развитие тринаксодона
Лучше один раз увидеть!
Дополнительная литература
20. Уходящие в воду
Происхождение китов
Амбулоцет
Китовые истории
«Великий морской змей»
Эволюция и киты
«Ходячий плавающий кит»
Кратко о бегемотоподобных
Род гиппопотамов
Лучше один раз увидеть!
Дополнительная литература
21. Ходячие ламантины
Происхождение сиреновых
Пезосирен
Русалки!
Наука о русалках
Морские коровы уходят в море
В дебрях Ямайки
Из Африки
Стеллерова корова
Лучше один раз увидеть!
Дополнительная литература
22. Лошадь зари
Происхождение лошадей
Эогиппус
Цок-цок
«Лошадь зари»
Как же их называть?
Откуда же взялась лошадь?
Лучше один раз увидеть!
Дополнительная литература
23. Гигантские носороги
Крупнейшее сухопутное млекопитающее
Парацератерий
Зыбучие пески!
Монгольские чудовища
Откуда пошли носороги
Биология гигантских носорогов
Страна гигантов
Индрикотерий в массовой культуре
Лучше один раз увидеть!
Дополнительная литература
24. Отражение обезьяны?
Древнейший ископаемый предок человека
Сахелянтроп
Отражение обезьяны?
Геологические часы
Молекулярные часы
Тумай
Лучше один раз увидеть!
Дополнительная литература
25. Люси в небе с алмазами
Древнейший человеческий скелет
Австралопитек афарский
Происхождение человека
Из Евразии?
Из Британии?
Ребёнок из Таунга и миссис Плез
Удача Лики
Наследие Люси
Лучше один раз увидеть!
Дополнительная литература
*** Примечания ***