А. С. Серебровский
Гибридизация животных
Предисловие
Настоящая книга является опытом сведения в систему чрезвычайно разнородных к часто отрывочных знаний, связанных с проблемой гибридизации животных. Будучи убеждены, что гибридизации животных предстоит сыграть большую роль в социалистическом хозяйстве, мы считали такую сводку полезным, хотя и трудным делом. Руководствуясь гениальным тезисом Маркса, «философы лишь различным образом
объясняли мир, но дело заключается в том, чтобы
изменить его», — мы расположили изложение вокруг вопросов овладения процессами гибридизации как одного из путей переделки животного организма. Рассматривая здесь преимущественно общие проблемы гибридизации, мы оставляем вопросы «частной гибридизации» для второй части сводки, для написания которой время однако еще не вполне созрело.
Ввиду обилия и разнообразия затронутых вопросов в книге несомненно найдется немало пробелов и оплошностей. В ряде случаев мы с благодарностью воспользовались указаниями тт. С. Н. Боголюбского, П. К. Денисова, П. А. Косминского, М. Л. Левина, С. Г. Левита, В., К. Милованова и мн. др. Мы также признательны ряду указанных в тексте лиц, любезно разрешивших нам воспользоваться еще не опубликованными данными и иллюстрациями.
Автор будет признателен за все указания читателей, которые просит направлять в лабораторию генетики МГУ.
Автор
Глава I. Гибридизация как наука
Гибридизация и гибриды
Поскольку предметом настоящей книги будет проблема овладения процессами гибридизации, необходимо прежде всего условиться относительно содержания этого термина. Мы будем говорить о гибридизации как о
межвидовой гибридизации и соответственно называть гибридами межвидовых гибридов независимо от того, относятся ли скрещенные виды к одному роду или к разным родам, семействам и пр. Таким образом под гибридизацией мы будем разуметь процессы, связанные со скрещиванием различных видов (типа линнеонов) и с получением первого и следующих поколений гибридов, возникающих в результате этих скрещиваний.
Такое определение гибридизации может нас удовлетворить несмотря на то, что попытка более точного определения приводит к большим трудностям. Эти трудности возникают прежде всего на того, что в наше определение мы вводим указание на различные
виды. До Дарвина это указание клало бы резкую грань между продуктами скрещивания различных видов и различных особей внутри вида, поскольку природа межвидовых и внутривидовых различий считалась совершенно разной.
Порожденное дарвинизмом учение о виде решительно изменило это понимание, сняло это принципиальное различие, и гибриды между особями, расами, разновидностями, видами, родами и т. д. стали в один непрерывный ряд. Разделение в наше время этого ряда на две части — на внутривидовую и межвидовую гибридизацию — требует уже специального обоснования, так как какой-либо резкой грани между скрещиванием например видов и скрещиванием разновидностей не существует. Если поэтому мы продукт скрещивания видов назовем гибридом, то почему, спрашивается, мы не можем назвать гибридом продукт скрещивания подвидов?
Мало того, известно, что систематики придают сплошь и рядом совершенно различное таксономическое значение различным близким формам. Например тундряная куропатка (Lagopus rupestris) американскими авторами разбивается на десяток видов или подвидов, другие считают все эти виды за один, а третьи объединяют этот вид с еще некоторым соседними L. hyperboreus, L. mutus и др. Помеси между всеми этими формами приходилось бы считать то гибридами то нет в зависимости не от объективных особенностей процессов скрещивания их друг с другом, по от более или менее субъективных взглядов систематиков на положение нижней границы вида, на границы между видом и подвидом.
Еще более существенным является то, что генетика показала отсутствие
резких граней между разными категориями скрещиваний и в физиологическом отношении. Еще в начале XX в. принимали, что наследственность в межвидовых и во внутривидовых скрещиваниях подчиняется различным законам. Полагали, что законы наследственности, открытые Менделем и распространенные после 1900 г. генетиками на огромный круг организмов, действительны лишь для признаков, которыми различаются особи внутри видов (доминирование, расщепление и пр.), наследование же при скрещивании видов идет по особому типу «промежуточного наследования», где нет ни доминирования ни расщепления. Однако феномен промежуточности был расшифрован как лишь более сложный случай менделевской наследственности, и рядом работ было показано, что характерные явления менделизма могут быть обнаружены в любом межвидовом скрещивании, но лишь в различных осложненных формах.
Термин «гибрид»
[1] стал прилагаться одинаково и к гибридам рас и особей, даже самых близких, различающихся лишь одним-двумя генами; в зависимости от числа генов, которыми различаются родители, мы говорим вслед за де Фризом о моногибридах, дигибридах, тригибридах, полигибридах.
Нетрудно видеть однако, что наличие непрерывных и постепенных переходов между различными категориями скрещивании (особей, рас, подвидов, видов и т. д.) вовсе не должно закрывать собой наличия глубоких качественных различий на отдельных звеньях этой цепи. Если мы, базируясь на некоторой условности в определении границ видов и условности в определении границ между простой и осложненной менделистической наследственностью, будем отрицать целесообразность и правомерность выделения межвидовой гибридизации в более или менее специфическую область, то мы рискуем совершенно выхолостить все понятие о гибридах. В самом деле в менделизме гибриды противопоставляются гомозиготам чистых линий. Но, как известно, среди животных чистые линии или хотя бы отдельные вполне гомозиготные особи не существуют, так как вряд ли бывает случай, чтобы какое-либо животное имело родителей совершенно одинакового генотипа или получило от них совершенно одинаковые геномы, а среди людей и домашних животных этого уже никогда не бывает. Таким образом нам придется вообще всех животных называть гибридами, любые спаривания самца с самкой называть гибридизацией, хотя бы и «внутривидовой гибридизацией». Если из соображений теории эту гибридность любого животного и приходится иногда подчеркивать, то будет все же нелепостью на этом основании приравнять всю зоологическую систематику к систематике гибридов. Совершенно очевидно, что гибрид и гибридизация являются специфическими объективными категориями и наличие переходов от гибридов к обычным животным и от гибридизации к обычному размножению нас смущать не должно. Отсюда будет правильным обозначать просто гибридизацией именно межвидовую гибридизацию, а внутривидовую сопровождать, если нужно, этим прилагательным. Полезно наряду с гибридами и гибридизацией говорить о
субгибридах (
субгибридизации), когда речь идет о скрещиваниях форм, более близких, чем виды (подвиды, расы, породы), и
об ультрагибридах (
ультрагибридизации) в случаях гибридизации, более отдаленной, чем примерно, семейства (скрещивания представителей разных отрядов, классов, типов).
Гибридизация по сравнению с нормальным размножением характеризуется совокупностью целого ряда особенностей. Прежде всего зачатие гибрида происходит с большими или меньшими трудностями самой разнообразной природы, которые мы будем рассматривать в III и следующих главах. Некоторые из этих препятствий легко устранимы, другие трудно или почти вовсе не устранимы. Развитие гибридов может сопровождаться разными аномалиями и даже вовсе не итти или, наоборот, гибриды могут обнаруживать особенно пышное развитие (гетерозис). Далее в огромном большинстве гибриды оказываются с разными степенями бесплодия. Соединение в клетках разных наборов хромосом приводит к особенностям и неправильностям в передаче признаков по наследству, и потомство гибридов оказывается чрезвычайно многообразным и часто с большим процентом ненормальных, уродливых или вообще нежизнеспособных особей даже при нормальных по жизнеспособности гибридах первого поколения. Наконец это потомство оказывается нередко обладателем совершенно неожиданных свойств. Каждое из этих свойств, взятое в отдельности, может встречаться и у негибридов. Пони-жеребца трудно скрестить с крупной кобылой. Отдельные уродства, даже нежизнеспособность, могут наследоваться и в качестве моногибридных признаков. Бесплодие также может встречаться у негибридов. Изредка встречается у нормальных животных неправильность распределения хромосом и т. п. С другой стороны, у некоторых гибридов могут отсутствовать некоторые из перечисленных свойств. Но тем не менее если говорить о типичных гибридах, то для них
совокупность перечисленных особенностей является совершенно характерной, и с другой стороны, у нормальных животных этой
совокупности свойств не бывает никогда. Чем дальше в таксономическом отношении отстоят друг от друга скрещенные виды, тем более резкие гибридные черты они имеют, и хотя от нормального животного до ультрагибрида существует непрерывный переходный ряд, тем не менее при известном удалении родителей наступает уже это повое качество потомства, которое мы не можем называть нормальным животным, а вынуждены называть гибридами. Эта степень удаленности соответствует примерно видовой удаленности, почему мы и считаем достаточно обоснованным ввести в определение гибрида указание на его межвидовую природу.
Необходимо ввести и еще одно ограничение в понятие о гибридах, ограничение временного характера. Как мы увидим подробнее в следующей главе, наши основные домашние животные (собака, корова, овца, свинья, лошадь) ведут свое происхождение от нескольких видов. Иными словами, их эволюция протекала с участием гибридизации и в этом отношении их можно было бы назвать гибридами. Однако мы думаем, что и такое, распространительное употребление терминов гибрид и гибридизация будет нецелесообразным. Если гибриды охарактеризованы нами выше целой совокупностью признаков, то в настоящее время ни один из этих признаков уж не приложим к нашим домашним животным (кроме мулов). Они совершенно свободно размножаются, процент оплодотворения и жизнеспособность их нормальны, механизм наследственности действует вполне нормально, характерного для гибридов многообразия потомства у них нет и т. д. Если они и обладали когда-либо этими особенностями, то в настоящее время они ничем не отличаются от нормальных животных и следовательно уже перестали быть гибридами. Указанный вывод, что гибриды при их дальнейшем размножении могут утрачивать свои свойства, характерные для гибридов (дегибридизация) и тем самым переставать быть гибридами, для нас очень важен, так как он вносит большую точность в объем понятия гибридов и позволяет нам не называть обычное разведение домашних животных гибридизацией. Поэтому данное выше определение гибридов мы должны дополнить указанием, что гибридами являются продукты скрещивания видов на протяжении такого числа поколения от момента скрещивания, пока эти формы сохраняют свойства гибридов. Для разных случаев это число поколений может быть весьма различным.
Проблема овладения гибридизацией. Краткий очерк
Что необходимо для того, чтобы мы имели возможность в полной мере овладеть процессами гибридизации животных и направлять их на пользу социалистическому строительству? Что нужно знать и узнать для этого? Каковы должны быть в общих чертах программа и содержание этой
науки об управлении процессами гибридизации или, как мы предложили ее назвать (Серебровский, 1933),
гибридагогии[2].
Здесь прежде всего можно выделить три основные раздела, которые должны входить в эту науку: 1) знание мировых ресурсов, 2) умение поставить задачи работы и 3) овладение методами достижения поставленных задач.
Для того чтобы иметь возможность широко ставить проблему гибридизации животных для различных сельскохозяйственных (например создание новых форм домашних животных) и других целей (облегчения акклиматизации дичи и пушнины и др.), необходимо почти заново разработать вопрос о наличных «мировых ресурсах». Под мировыми ресурсами гибридизации мы разумеем те дикие виды и домашние породы, которые могут быть попользованы для тех или иных гибридизационных целей. В силу этого подлежащими охвату изучением в качестве «мировых ресурсов» формами будут конечно далеко не все виды животных, но в первую очередь те, которые более или менее близко примыкают прежде всего к нашим 20–25 видам домашних животных: копытные, примыкающие к быкам, козам и овцам, к северному, оленю и маралу, к лошади и ослу, к верблюду, свинье; хищные, примыкающие к собаке, лисице и кошке; грызуны, примыкающие к кролику, морской свинке, мышам и крысам, белке, нутрии, ондатре; куриные и утиные птицы, некоторые группы рыб, пчелиные, шелкопряды и т. п. С другой стороны, эти группы должны быть подвергнуты несравненно более детальному изучению, чем это сделано до сих пор. Должны быть изучены их анатомия, гистология, экология, физиология и особенно генетика, они должны быть оценены под зоотехническим углом зрения и выяснено, какие перспективы может открыть гибридизация их друг с другом и особенно с нашими домашними животными.
Само изучение мировых ресурсов гибридизации должно однако вовсе не ограничиваться только зоологическим изучением видов. Оно должно вестись в трех плоскостях: 1) ресурсы видов, 2) ресурсы признаков и 3) ресурсы генов.
Знание видов, их родственных отношений должно нам дать указания на то, кого мы вообще можем вовлечь в гибридизацию, кого непосредственно можно скрестить например с домашней овцой, кого использовать в качестве «гибридизационного моста», при помощи которого можно будет перекинуть мост между нескрещиваювщимся видами. Для этого изучения должно быть привлечено и использовано все, что накоплено в этой части систематикой, морфологией, экологией, зоогеографией, палеонтологией, археологией и др. и особенно должна быть заново развернута разработка тех вопросов, которые этими науками еще не были разработаны, а для нас особенно важны.
Не менее важно изучение ресурсов
признаков, так как несомненно, что не столько сами виды, сколько отдельные признаки этих видов будут представлять основной интерес для гибридизатора. Поэтому этот отдел изучения мировых ресурсов должен будет дать детальные монографии примерно такого типа: «Шерстный покров копытных сем. Bovidae», «Обзор величины строения и химического состава яиц Anatidae», «Обзор врожденного иммунитета в сем. свиней», «Респирационные константы и особенности обмена веществ в отр. куриных» и т. д. — систематический, детальный, глубокий анализ и сопоставление всех могущих интересовать гибридизатора признаков: анатомических, гистологических, физиологических, биохимических рефлексологических и др.
Наконец уже высший интерес будут представлять мировые списки
генов, свойственных различным диким видам и обусловливающих различия между разными видами. Во всяком межвидовом скрещивании следует различать «предсказуемую» часть результата и «непредсказуемую» часть, вернее предсказуемую лишь после того, как частная генетика этих видов будет более или менее изучена. Действительно, сравнивая хотя бы яка с бизоном, мы можем предсказать пока лишь возможность скомбинировать например оба типа их оброслости. Но поскольку разница в оброслости этих двух быков несомненно обусловлена многими генами, предсказать сейчас возможные типы оброслости, которые в таком скрещивании смогут возникнуть во втором и следующих поколениях в результате перекомбинировки этих неизвестных нам пока генов, мы не можем. Так как таких комбинаций несомненно будет иметься (теоретически) миллионы, то практически даже в очень большом втором поколении мы из них получим лишь ничтожную часть и может быть самых интересных для. нас не получим. Лишь составив на базе изучения этого расщепления представление о том, какие именно гены принимают в нем участие, мы в состоянии будем делать предсказания о возможных дальнейших результатах этой гибридизации. А так как умение предварительно рассчитать возможные результаты того или иного скрещивания будет иметь для всей работы крупнейшее значение, то и само изучение мировых ресурсов генов будет иметь для гибридизатора крупнейшее значение.
Чрезвычайно серьезной и совершенно новой является вторая часть теории гибридизации, именно разработка теории того, что можно назвать «проектированием животных форм). С вопросом о таком проектировании связаны по нашему мнению самые существенные черты отличия всей работы гибридизатора от работы обычного селекционера. В самом деле, каков основной метод работы селекционера? Селекционер имеет перед собой какую-нибудь породу животных, обладающих определенными показателями: данной средней молочностью, данной средней величиной, скороспелостью, склонностью к заболеваниям и т. д. При этом по универсальному закону изменчивости различные особи породы уклоняются от этих средних величин в плюс или минус сторону. Наряду с данной средней молочностью имеются различные — вплоть до крайних — плюс-варианты, которые привлекают к себе естественно внимание селекционера, и он ставит перед собой задачу передвинуть среднюю молочность в сторону плюса вплоть до крайнего возможного предела. Точно так же по проценту например смертности потомства внимание селекционера привлекают крайние минус-варианты, дающие наименьшую смертность, и он ведет селекцию в направлении передвижения средней в минус-сторону до возможного предела. Если в пределах породы его работа начинает исчерпывать возможности или становится слишком медленной, он обращает внимание на какую-нибудь иную породу у которой молочность еще выше, и путем скрещивания привлекает ее к повышению изменчивости и к расширению возможностей дальнейшей селекции.
Иными словами, селекционер, исходя из данных качеств своих животных, ставит себе сравнительно очень простую (теоретически) задачу путем отбора постепенно усиливать желательные признаки и ослаблять нежелательные. И вся сложность его работы состоит в технической реализации селекции, в возможной быстроте работы, в том, чтобы, погнавшись за одними признаками, не упустить или не испортить других и т. п.
В значительной степени иначе приходится работать, когда мы ставим себе задачу путем более или менее смелых скрещиваний пород создать новую по известному проекту. В наивысшем развитии этот путь работы стоит перед гибридизатором различных видов, и поэтому мы можем считать его характерным именно для
гибридизатора и от него именно требовать разработки теории «проектирования новых форм животных».
О каком же проекте здесь идет речь? Вот перед гибридизатором лежат доступные ему «мировые ресурсы» (возьмем в качестве примера быков и предположим, что технические вопросы гибридизации достаточно разрешены) — различная оброслость от буйвола до яка, совершенно различное соотношение передней и задней части тела, различный характер жира и мяса, различная кожа, несколько различный обмен веществ, температура тела, химия крови и пр. Все эти признаки могут быть различным образом комбинируемы и в результате возможно создание животных, чрезвычайно сильно отличающихся от наших обычных коров. Спрашивается, как должен гибридизатор распорядиться этими ресурсами и какое животное он захочет — или должен будет — создавать?
Ясно, что работать на-авось, «авось что-нибудь да выйдет интересное», он не может, не имеет права, как архитектор не имеет права на-авось складывать кирпич, балки и т. д. в надежде, что авось выйдет здание, для чего-нибудь пригодное. Очевидно, что подобно архитектору
гибридизатор должен тоже создать некоторый проект и, создав и хорошенько обсудив и оценив этот проект, приступить к его выполнению.
Какие вопросы должны быть проработаны при составлении этого проекта? Прежде всего очевидно должно быть оценено, каким потребностям социалистического хозяйства должно будет удовлетворять это животное. Этот вопрос может оказаться одним из труднейших. В самом деле, мы так привыкли к нашей корове, овце, лошади, что нам может казаться, что, собственно говоря, они уже заполнили все наши потребности и всякое новое животное в конце концов должно быть. или второй коровой, или второй овцой, или второй лошадью. Конечно это не так. У человека слишком много потребностей, а бывший перед ним выбор животных для одомашнения был слишком ничтожен, чтобы была хоть тень вероятности того, что человеку никакое другое животное кроме его коровы, овцы, лошади и т. п. не нужно. Вспомним, что растениеводы работают с несколькими тысячами видов растений и каждому находят его незаменимое место и все еще ищут новые виды для введения их в культуру. Таким образом теоретически проработать вопрос о том, какие еще формы быкообразных животных могли бы оказаться чрезвычайно полезными человеку — задача чрезвычайно важная, она дала бы мощный толчок всей работе гибридизации. Здесь должны быть учтены например потребности различных районов и климатов, неудовлетворенные потребности промышленности, различные проблемы комбинации направлений продуктивности (например молочно-шерстные быки, мясные лошади для полупустынь и т. д.).
Следующая проблема проектирования — проблема соединимости признаков. Из элементарного менделизма мы знаем о возможности комбинирования признаков — можно например скомбинировать любую форму гребня курицы с любой окраской и любой формой оперения. Но совсем иначе обстоит вопрос о возможности комбинирования взаимно исключающих или мешающих друг другу признаков и свойств. В зоотехнической литературе уже много дебатировался вопрос о возможности скомбинировать выдающуюся молочность с выдающейся мясностью — вопрос этот еще чрезвычайно далек от разрешения. особенно если мы возьмем признаки в таком размахе, в каком мы их встретим в наших мировых ресурсах. Для этого должен быть по-новому освещен вопрос об общей энергетике организма, о физиологической основе тех или иных его свойств, о возможности параллельного существования в одном организме этих процессов и т. д. — ряд вопросов, восходящих очевидно до самых глубин познания общей физиологии и морфологии организма.
 Рис. 1. Клювы различных видов и родов гусей.
Рис. 1. Клювы различных видов и родов гусей.
Далее значительна проблема о важных и неважных, необходимых и не необходимых частях организма. Нужно ли в таком проекте предусматривать и ушную раковину, и слепую кишку, и четыре соска, а не два, нужно ли столько позвонков, сколько их есть, или можно дело упрощать и здесь и т. д. Проработка этих вопросов позволит нам ориентироваться в возможных пределах реорганизации животного тела, полученного нами в наследство от стихийного процесса органической эволюции. Совершенно очевидно, какое огромное поле науки
должно быть здесь вспахано, прежде чем мы сможем дать обоснованные ответы на подобные вопросы. Здесь должны быть использованы физиология, сравнительная анатомия, все вообще эволюционное учение, проведен длинный ряд генетических и хирургических экспериментов и т. д.
Все подобные вопросы придется иметь в виду при проектировании и отдельных органов. Маленький пример — мы задавали нескольким биологам различных специальностей следующий вопрос. У разных уток и гусей имеется большой выбор различных клювов — длинные и короткие, широкие и узкие, прямые и с крючками, твердые и мягкие и др. (рис. 1 и 2). Какой клюв наиболее целесообразно дать нашей домашней утке? Или у нее уже «самый лучший» клюв? Ответ на этот вопрос был в общем одинаков — современная наука совершенно не подготовлена к ответу на этот вопрос! Для этого должна быть изучена экология диких видов под этим специальным углом зрения и выяснено более точно экологическое значение различных форм клюва. Далее должна быть изучена современная экология домашней утки, выяснены перспективы изменения этой экологии, и только тогда мы сможем дать ответ на поставленный вопрос.
 Рис. 2. Клювы представителей различных подсемейств утиных: широконоски, гаги фишеровой, крохаля, пеганки и лебедя.
Рис. 2. Клювы представителей различных подсемейств утиных: широконоски, гаги фишеровой, крохаля, пеганки и лебедя.
Мы видим таким образом, что проблема проектирования новых форм животных должна будет разрастись но всеобъемлющую теорию, синтезирующую достижения всех почти ветвей биологии. Для возможности этого синтеза придется разработать множество вопросов, которые биологией пока вовсе не ставились, и не потому, что биология не могла бы их осветить, но потому, что эти вопросы просто не ставились со стороны производства. Они естественно вытекают из запросов гибридизации и только специалистами этого типа будут разрабатываться.
Для того чтобы эти перспективы не показались слишком «страшными», следует сказать, что мы здесь имели в виду так сказать высшую форму проектирования. В конкретных условиях, особенно в начале работы по гибридизации, наши проекты будут гораздо более скромными и посильными. Составить проект короны с сильной оброслостью, скомбинированной из оброслости яка и бизона, или что-либо подобное будет в наших силах конечно скоро. Не так трудно составить проект собаки с шерстью лисицы или лисицы с размерами и поведением собаки, а дальнейшие уточнения и коррективы к такому эскизу вносить в ходе самой работы. Но в ходе этой работы будет разработана и сама теория подобного проектирования, которая позволит в дальнейшем ставить более глубоко и смело проблему реконструкции животного организма.
На сегодня однако более очередными являются многочисленные вопросы третьего раздела гибридагогии — раздела о методах гибридизации. Подлежащие здесь рассмотрению вопросы более ясны и могут быть перечислены короче.
Первый круг вопросов связан с проблемой получения гибридов и преодоления возникающих здесь затруднений. В тех случаях, когда виды более или менее свободно скрещиваются друг с другом, достаточно просто привести их в одно место и дать возможность такому скрещиванию осуществиться. Основное внимание в этом случае должно быть сосредоточено на самом разведении и содержании различных диких видов, которые часто оказываются привезенными из других стран, из других климатов, и гибридизатору придется неизбежно овладеть всей своеобразной техникой содержания и разведения диких животных. Иными словами, мы должны заняться теоретически всем множеством вопросов, возникающих в зоологических садах, и в частности — проблемой акклиматизации животных под тем специфическим углом зрения, что гибридизатору необходимо этих животных не просто содержать, а содержать их так, чтобы они обнаруживали стремление к размножению и находились в соответствующем физиологическом состоянии. Это, как известно, является одной из самых трудных задач, стоящих перед зоосадами, по крайней мере по отношению к целому ряду видов, которые прекрасно живут в зоосадах, но никогда или очень редко размножаются (пример краснозобые казарки, соболь и пр.).
Если скрещивания встречаются с затруднениями, то перед гибридизатором встает в добавление к только что указанным длинный ряд новых проблем, подлежащих преодолению самыми разнообразными методами. Здесь прежде всего могут встретиться затруднения биологического характера, например несовпадение сроков размножения скрещиваемых видов, особенно если они привезены из разных широт и разных полушарий. Совершенно ясно, что разрешить задачу — свести сроки размножения двух видов к совпадению — можно, исходя из очень различных отправных пунктов, например искусственным вызыванием течки и другими эндокринными воздействиями, теплым или холодным содержанием, различным кормлением, постепенным из поколения в поколение оттягиванием сроков родов или замедлением или ускорением полового созревания и т. д. вплоть до длительного хранения спермы одного вида до начала половой деятельности самок другого вида.
При совпадении сроков размножения может все же возникнуть длинный ряд дальнейших затруднений: механические затруднения для скрещивания, рефлексологические затруднения («нежелание» самца спариваться с чужой самкой, воинственное поведение самца по отношению к самке и пр.). Для преодоления этих препятствий уже указывались различные приемы, но в настоящее время несомненно, что самым радикальным способом этого преодоления должно явиться искусственное осеменение, и потому разработка вопросов искусственного осеменения применительно к самым разнообразным млекопитающим, птицам, насекомым должна привлекать большое внимание гибридизатора.
Комплекс вопросов здесь очень велик и разнообразен: специфическое изучение анатомии половых органов, изучение биохимии и биофизики спермы и различных жидкостей тела самки (вагинальные, маточные, трубные и брюшинные выделения), физиологии акта эякуляции, механические, электрические, хирургические приемы получения спермы, тщательное изучение физиологии течки разных видов, включая овуляцию, конкретная техника искусственного осеменения и его аппаратура, хранение и разбавление спермы и т. д.
Однако, преодолев проблему гетерогенного осеменения, мы подходим вплотную к еще более важной проблеме — проблеме гетерогенного оплодотворения. Эта проблема в настоящее время изучена гораздо глубже, чем предыдущая, но тем не менее здесь мы имеем еще огромные пробелы и поэтому огромное поле для исследований. К тому же пока вопрос разрабатывался в отрыве от производственных задач, и конкретная задача заставить чужой сперматозоид оплодотворить чужое ему яйцо если и ставилась, то лишь по отношению к животным, менее всего интересующим производство, — иглокожим, молюскам и пр. (исключением являются рыбы). По линии млекопитающих и птиц здесь не сделано ничего, и даже едва начата разработкой та техника, на базе которой эта проблема может разрешаться, например первые работы по культивированию яиц млекопитающих in vitro и получение оплодотворения их под микроскопом в условиях культуры тканей. И ясно, что для того чтобы иметь возможность управлять гетерогенным оплодотворением, содействовать ему, необходимо на конкретных объектах, интересующих животноводство, и на подсобных объектах поднять разработкой огромную область биологии и биохимии, культуры тканей и т. д.
Еще более важной проблемой гибридизации — можно без преувеличения сказать —
основной ее проблемой — является проблема преодоления бесплодия гибридов. Она является основной потому, что уже в настоящее время с большим или меньшим трудом мы имеем возможность получать огромное разнообразие гибридов. Например по группе уток и гусей можно предполагать, что мы можем получить не менее 40 000 разнообразных парных гибридных сочетаний из тех 200–250 видов, которые входят в эту группу, но подавляющее число их бесплодно. А так как основной интерес гибридагога направлен естественно на второе и дальнейшие поколения гибридов, то сейчас остается для этого лишь ничтожная часть уже получающихся гибридов. Преодоление бесплодия было бы огромной, решающей победой на фронте гибридизации, Но сложность этой проблемы ясна. Она связана, с генетикой пола, с проблемой управления митозами, в которых принимают участие гетерогенные хромосомы. Она связана с такими например вопросами, как необходимая температура и другие условия для сперматогенеза млекопитающих, и наконец с обширной областью эндокринологических вопросов, так как возможно, что именно на этом пути будет найдено разрешение проблемы: вопросы воздействия на организм гибрида с ранних стадий эндокринными препаратами, вопрос трансплантации гибридных гонад в полноценные в половом отношении взрослые организмы, вопросы культивирования in vitro гаметогенной ткани, проблема полиплоидии и т. д.
Ясно, какой обширный арсенал самых разнообразных знаний и методик должен быть мобилизован для разрешения этой проблемы из проблем, разрешение которой сделает нас подлинными творцами новых форм животных.
Можно было бы еще дальше излагать круг проблем, разработка которых должна быть организована для того, чтобы гибридагогия действительно дала нам в руки ключ к управлению гибридизацией в интересах социалистического строительства. Мы ограничимся однако уже сказанным, упомянув в заключение еще об одной интересной проблеме — проблеме экологии гибридов. В самом деле, когда мы скрещиваем два вида, различающиеся своей экологией, то очевидно «нормальная» экология гибрида должна быть какой-то тоже гибридной — может быть промежуточной, может быть атавистической (см. гл. IX), может быть вовсе новой. Для того чтобы воспитывать, выращивать полученных гибридов, получать от них максимум, который они могут дать, мы очевидно должны научиться как-либо предугадывать или отыскивать эти новые экологические потребности.
Совершенно ясно, что развитие всякой науки стимулируется развитием и потребностями производства, а не субъективными интересами отдельных исследователей. Как заметил Энгельс, «до сих пор хвастались тем, чем производство обязано науке, но наука бесконечно большим обязана производству». Поэтому для уяснения перспектив нашей пауки в СССР необходимо уяснить себе те перспективы, какие-открываются для гибридизации животных в условиях социалистического хозяйства, чему и будет посвящена следующая глава.
Глава II. Роль гибридизации животных в прошлом и ее перспективы
Для того чтобы правильно оценить перспективы гибридизации животных и ее значение для различных отраслей народного хозяйства, очень важно проследить историю гибридизации, выяснить, что она уже дала, как использовалась она в различные периоды человеческой истории, что стимулировало и что тормозило это использование. Анализ истории гибридизации позволит нам правильнее оценить ее будущее, особенно в тех новых условиях, которые создаются построением социализма.
Гибридизация в истории домашних животных
К сожалению восстановление истории гибридизации животных человеком в наиболее интересных главах встречается с очень большими трудностями. С одной стороны, несомненно, что человек начал гибридизировать попавших в орбиту его хозяйства животных с чрезвычайно древних времен. Уже одомашнивая в разных географических областях свое первое животное — собаку, — человек тем самым одомашнил целый круг различных форм ее предков — от типичного северного волка до индийских волков и может быть шакалов, Вовлекая эти одомашненные формы в процесс своих переселений, человек приводил в соприкосновение друг с другом такие формы собак, которые естественно никогда не встречались, будучи географически удалены и изолированы друг от друга. В условиях одомашнения это соприкосновение легко приводило к гибридизации, создавая ту глубокую изменчивость, которая несомненно сыграла крупную роль в усовершенствовании пород собак. То же самое можно сказать и относительно ряда других животных — свиньи, овцы, козы и др.
Однако эти соображения слишком общи. Для оценки роли гибридизации в прошлом необходимо выяснить следующие вопросы: 1) какие именно животные были подвергнуты гибридизации и какие не были гибридизированы вовсе, 2) какие виды из числа гибридизированных были гибридизированы настолько глубоко, что несколько диких видов приняли примерно равное участие в создании пород, и какие виды лишь частично, местами или недавно восприняли примесь генофонда других видов, 3) как (хотя бы приблизительно) проходила эта гибридизация хронологически и 4) в качестве итога, как различные степени, характер и хронология гибридизации отразились на темпе эволюции соответственного вида животных. Ясно, что. ответить на вопрос о том, какую роль сыграла гибридизация в эволюции того или иного животного, можно лишь осветив предварительно намеченные вопросы. При этом должна быть строго учтена и специфика эволюции домашних животных, так как темпы эволюции разных видов вовсе не являются, как иногда полагают, более или менее прямой функцией степени изменчивости данного вида. Если например у голубей имеется множество пород, а у кошек — очень мало, то это вовсе не значит, что физиологическая наследственная изменчивость у голубя сильнее, чем у кошки. Дело может обстоять и обратно
[3], но условия отбора голубей по ряду причин и биологических и социально-экономических несравненно более благоприятны, чем условия отбора кошек. Оба эти вида развивались в одомашненном состоянии без гибридизации.
Ответить на перечисленные вопросы однако вовсе не так легко. Начало эволюции многих домашних животных восходит к древнейшим эпохам человеческой истории, и восстанавливать ее приходится косвенными и часто малонадежными методами. Дикие предки таких животных, как корова, уже истреблены. Различные исследователи допускают подобных же вымерших прародителей и для других видов домашних животных (лошадь, коза и пр.), и у нас нет в настоящее время надежного метода для того, чтобы ограничивать это «предкотворчество». Как известно, некоторые исследователи доводили это предкотворчество до абсурда, предполагая почти для каждой породы домашних животных особого вымершего дикого предка. Так, Фитцингер предполагал особых диких предков для хохлатых, для пятипалых и прочих пород кур, насчитывая таких диких предков почти два десятка. Развитие мутационного учения и изучение генетики этих признаков (хохла, полидактилии и пр.) сделали очевидной абсурдность. подобной точки зрения. Тем не менее вопрос о том, для каких видов следует допустить существование вымерших предков, остается еще далеко невыясненным, и подобные предки принимаются для ряда наших домашних животных: Bos nomadicus наряду с реальными Bos primigenius — для крупного рогатого скота, Capra prisca — для коз, Equus abeli — для лошади и пр. В этом вопросе несомненно имеются элементы порочного круга: невыясненность числа предков затрудняет понимание закономерностей эволюции домашних животных, а невыясненность закономерностей эволюции затрудняет выяснение числа предков. И хотя в эти вопросы и вносится постепенно все большая и большая ясность, тем не менее многие важные детали остаются еще темными. И наконец положение (с точки зрения интересующего нас сейчас вопроса) осложняется еще и тем, что даже в том, случае, когда исследователи более или менее договариваются относительно предков, как например Bos nomadicus и Bos primigenius для крупного рогатого скота, возникает важный вопрос, в какой степени эти предки являются действительно различными видами или же лишь географическими формами, так как относительно вымерших предков мы ничего кроме их остеологии, часто даже краниологии, не знаем — нам не известен ни их хромосомный аппарат, ни физиологические особенности; не известно, — с какой легкостью они скрещивались друг с другом (так ли например легко, как зебу и корова, или же гораздо труднее, как як и корова), были ли в этих скрещиваниях элементы нарушения плодовитости и т. д. — вопросы для оценки характера и роли гибридизации чрезвычайно важные.
Современное состояние вопроса о происхождении домашних животных в кратких чертах сводится к следующему. Прежде всего довольно длинный ряд домашних животных несомненно имел монофилетическое происхождение. Таковы прежде всего птицы — голуби, куры, утки, индейки, цесарки, канарейки с их предками Columba livia, Gallus bankiva, Anas platyrhynchus, Gallopavo meleagris. Относительно голубя и курицы делаются попытки доказать их гибридное происхождение. Так, Гиджи (Ghigi, 1922), основываясь на возможности скрещивания Columba livia с Columba leuconota, отстаивает участие также последнего вида в образовании домашних голубей, а основываясь на возможности гибридизации домашних кур с пестрым яванским петухом Gallus varius — участие последнего в образовании пород кур. К этим попыткам однако следует отнестись критически. Сам факт возможности плодовитого скрещивания вовсе не служит аргументом в пользу причисления данного вида к предкам. Известно например, что домашние козы дают плодовитых гибридов с альпийским козерогом (Capra ibex), но никому не приходит в голову причислить козерога к предкам коз, хотя бы уже по совершенно иному строению рог.
Точно так же на Яве и других Зондских островах любители бойцового спорта скрещивают Gallus varius с домашними курами, и гибриды оказываются (всегда? часто?) плодовитыми (Beebe, 1918–1922). Возможно поэтому, что местные куры в некоторой степени имеют примесь генов G. varius. Однако отсюда еще очень далеко до возведения G. varius в ранг предков кур, как наличие в некоторых районах гибридов крупного рогатого скота с яком вовсе еще не дает права возводить яка в ранг одного из предков коров. У пестрого петуха имеется. целый ряд очень своеобразных и красивых признаков (хвост, перья, грива и пр.), и если бы он действительно принимал участие в происхождении кур, то хотя какие-либо из признаков этого вида встречались бы и среди домашних кур. Правда, Гиджи делает попытку принять за такой признак черную окраску кур, основываясь на том, что при гибридизации кур с G. varius извлекаются черные формы. Однако слабость этого аргумента несомненна, она воскрешает позиции Фитцингера. Нужно доказать, что черный цвет (если даже будет доказано, что он генетически один и тот же у кур и у G. varius) не мог возникнуть у кур мутационно, как он возник у индюшек, уток, голубей и множества других видов диких птиц, где меланизм является одним из самых обычных мутационных признаков.
Из млекопитающих монофилетическое происхождение можно считать несомненным для следующих видов (Hilzheimer 1912/13; Klatt, 1927): кошка (от египетской буланой кошки Felis maniculata), домашний хорек (от Putorius eversmanbi) (Dubois, 1925; Miller, 1912), лама (от гуанако), северный олень (от Rangifer tarandus), буйвол (от индийского буйвола), кролик (от южноевропейского Oryctolagus cuniculus), морская свинка (от Cavia cuthleri), лисица и другие недавно прирученные пушные животные: нутрия, ондатра.
Из домашних рыб несомненно монофилетичными являются карп (от сазана Cyprinus carpio) и золотая рыбка.
Для большинства перечисленных форм характерно весьма незначительное породное разнообразие, однако крупнейшими исключениями являются птицы — куры и особенно голуби, глубина породного разнообразия которых стоит пожалуй на одном из первых мест среди всех домашних животных, соперничая даже с собакой.
Что касается домашних животных
полифилетического происхождения, то здесь на первом месте по ясности картины нужно поставить свинью и гуся, так как для них, с одной стороны, несомненно наличие двух предков у каждого и, с другой стороны, эти предки живут и ныне и нет основания предполагать каких-либо вымерших предков.
Свинья была несомненно одомашнена и в Европе, где диким предком явился кабан (Sus scrofa), и в Юго-восточной Азии, где одомашнению подверглась полосатая свинья (Sus vittatus)
[4]. Уже с античных времен началась гибридизация европейских домашних свиней с пришедшими в Европу восточно-азиатскими, и этот процесс идет и по сие время, зайдя настолько глубоко, что в чистой форме потомков кабана среди домашних свиней более культурных стран уже едва ли возможно найти. В соединении с интенсивным отбором и мутационным процессом этот гибридизационный процесс, особенно в последнее столетие, способствовал глубочайшим изменениям анатомии и физиологии свиньи, доводя ее нередко до явно патологических степеней.
Несколько иной является история
гуся. Здесь также несомненно, участие двух доныне живущих диких видов — серого гуся (Anser cinereus), одомашнивавшегося повидимому на очень широкой территории, и гуся-сухоноса (Cygnopsis cygnoides) — на Дальнем Востоке и в Юго-восточной Азии. Оба эти гуся еще яснее, чем названные виды свиней, являются различными видами, так как серого гуся и сухоноса с достаточными основаниями относят даже к разным родам. Что касается рода Anser, довольно полиморфного, то здесь возможна Примесь и некоторых других близких и достаточно легко скрещивающихся видов (например A. albifrons), хотя основным видом несомненно является именно серый гусь. В отличие от свиней гибридизационный процесс у домашних гусей зашел не так далеко, и большинство пород домашних гусей является чистыми потомками серого гуся. Однако Блитсом для гусей Индии уже был отмечен их смешанный гибридный характер, и эта гибридизация могла итти и на территории России, где ареалы обоих диких видов встречаются в Сибири (см. также Westwood, 1842, и его ссылку на Atheneum, 1836, р. 636). А с недавним проникновением к нам домашних пород китайского гуся, происходящего от сухоноса, эта гибридизация получила широкое распространение. Одна из лучших пород гусей, возникшая в России — холмогорский гусь, — несет на себе явные следы своего гибридного происхождения.
Два других вида гусей, одомашненные человеком — нильский гусь (во времена Египта) и канадская казарка (в недавнее время в Сев. Америке), — не отразились на эволюции домашних гусей благодаря бесплодию гибридов с ними.
Более сложной и менее ясной является история гибридизации остальных домашних животных, прежде всего собаки.
Что в образовании
домашней собаки, наиболее многообразного домашнего животного, приняло участие несколько диких видов, не подлежит сомнению, хотя исследователи еще далеко не достигли единства мнения относительно списка этих форм. Семейство собак довольно богато видами, распространенными по всему земному шару, и многие представители его сравнительно легко и вероятно многократно приручались человеком и даже скрещивались с ранее одомашненными собаками.
Однако, как мы только что упомянули, одной возможности скрещивания еще недостаточно, чтобы отнести данный вид к числу видов, действительно принимавших реальное участие в образовании домашних пород, и выяснение этих реальных предков среди возможных кандидатов представляет тем более сложную задачу, чем больше этих кандидатов. Дарвин, детально проработавший этот вопрос, считал «весьма вероятным, что домашние собаки, сколько их есть на свете, произошли от двух хорошо разграниченных видов волка (именно Canis lupus и С. latrans
[5]), еще от двух или трех сомнительных видов (именно европейского, индийского и североафриканского волка), по крайней мере от одного или двух южноамериканских видов собак, от нескольких рас или видов шакала и быть может от одного или нескольких вымерших видов».
Несомненно отпадают из числа предков собак такие роды, как азиатский красный волк (Cyon), африканская гиеновая собака (Lycaon) и многочисленная группа лисиц и песцов. По существенным краниологическим отличиям эти ветви можно проследить до олигоцена и даже эоцена (Гекели), и каких-либо указаний на примесь их генотипа к породам собак мы не находим. Менее ясен вопрос с южноамериканскими дикими видами, являющимися в известных отношениях связующими звеньями между лисицами, с одной стороны, и волками и шакалами — с другой. К сожалению древние домашне породы южноамериканских собак недостаточно изучены. Индейцы Гвианы скрещивали своих собак с диким Canis cancrivurus. Однако Лоннберг (Lönnberg, 1919), изучивший собаку индейцев Огненной Земли, не нашел у нее никаких признаков родства с южноамериканскими дикими видами, так что неясно, скрещиваются ли южноамериканские дикие собаки с завезенными из Старого Света или только с своими одомашненными потомками. К тому же невидимому и гибриды собак с южноамериканскими дикими видами бесплодны.
Таким образом круг возможных предков домашней собаки значительно суживается до группы собак в собственном смысле слова — волков и шакалов. Из них наиболее вероятными кандидатами следует признать индийского Canis pallipes, волка С. lupus и египетского С. lupaster.
Основываясь на грациозной, долихосомной форме абиссинского волка (род Simenia), ряд авторов в последнее время (особенно Келлер) отстаивал в нем предка борзых собак. Однако Штудер (Studer, 1903, ср. Heller, 1903) и Гильцхеймер (1908) возражают против этой возможности, а последний, следуя Гекели, относит Simenia к группе лисиц. Также недостаточно ясным остается вопрос с австралийским динго, которого однако правильнее считать за одичалую собаку, завезенную сюда первобытным охотником австралийцем.
Для оценки степени гибридности собак особое значение имеет вопрос об участии в их образовании шакалов. Хотя указанные выше волки и относятся систематиками к нескольким диким видам, однако они настолько близки друг к другу и настолько связаны переходами, что могут быть истолкованы в качестве географических популяций единого большого вида, несколько диференцировавшегося в соответствии с климатическими и прочими особенностями отдельных частей огромного ареала
[6].
Иначе дело обстоит с видами шакалов, которых относят уже к отдельным родам. Эти виды встречаются в тех же географических районах, как и некоторые волки, не смешиваясь уже с ними, так что их видовая самостоятельность несмотря даже на наличие некоторых переходных форм не подлежит сомнению. Точно так же и отличия их от волков, хотя бы по размерам, гораздо значительнее, чем в пределах волков. Доказать, что в собаке мы имеем смешение обоих обширных генофондов, волков и шакалов, было бы поэтому чрезвычайно важным. До недавнего времени большинство авторов причисляло шакалов к предкам ряда мелких пород собак. Однако в последнее время здесь появились сомнения. Так, Гильцхеймер, ранее признававший шакалов предками собак, в последние годы занял скептическую позицию, так же как и Антониус и ряд других авторов. В частности Штудер, Schame (1922) и Г. Миллер (1912) подчеркивают ряд особенностей в строении зубов шакалов, особенно верхних коренных, которые якобы не встречаются у собак. Однако Klatt (1927), придавая большое значение работе Миллера, отмечает, что в его материале некоторые черепа по этим признакам вполне совпадают с черепами серых шакалов из Эритреи, ясно отличаясь от волков, хотя значительное большинство собак, как больших, так и мелких, действительно соответствовало по зубам волчьему, а не шакальему типу. С другой стороны, не известно, у всех ли форм шакалов имеется этот шакалий тип зубов. Поэтому если даже отбросить из числа предков; чапрачного и полосатого шакалов, то остаются чрезвычайно многоформные серый и золотистый, образующие на своем обширном ареале ряд форм, от мелких южных до крупных более северных, по ряду признаков приближающихся и переходящих к волкам (Hilzheimer, 1908). Поэтому все же надо думать, что больше данных говорит не против, а за участие шакалов в образовании генофонда-собак, и подлежит уточнению лишь вопрос об удельном весе этого участия.
Несколько иначе обстоит вопрос о гибридизации домашних
овец. Еще Линней, отнеся большинство овец к виду Ovis aries, выделил цакелей в особый вид за своеобразную горизонтальную форму рогов. Впоследствии Фитцингер и другие принимали множество вымерших родоначальников для разных пород овец. Подобные деления лишены оснований. Однако, прослеживая историю овец, большинство авторитетов склоняется к тому, что в создании овец принимали участие несколько видов, — как европейские муфлоны, так и азиатские уриалы и аргали. Келлер и некоторые другие авторы отстаивали участие и гривистого барана, относимого теперь к особому роду Ammotragus (A. lervia). Это мнение они основывают на сходстве оброслости некоторых африканских и древнеегипетских пород с оброслостью гривистого барана. Однако если даже гривистый баран и мог быть, одомашнен и дал начало некоторым африканским породам, то считать его в числе предков обычных домашних овец невозможно уже потому, что скрещивание его с домашними овцами не удается (попыток скрестить его с предполагаемыми его домашними африканскими потомками повидимому никем не делалось. Повидимому науке не известны попытки скрестить эти африканские породы и с обычными овцами). Таким образом этот вид отпадает, и в качестве родоначальников овец остаются лишь представители рода Ovis. Однако сказать, какие именно дикие виды приняли участие в образовании домашних овец, не только трудно, но повидимому и невозможно. Дело в том, что систематика самих диких баранов чрезвычайно затруднительна и при желании «можно насчитать столько видов, сколько имеется бассейнов горных рек на огромном ареале распространения рода Ovis». Так как, надо думать, все виды этого рода способны свободно смешиваться друг с другом, то только перерывы ареала [например американский толсторог (О. montanum), отделенный от азиатских] создают некоторые более резкие различия. Но такие границы разной степени и силы создаются и хребтами и равнинами, и в результате почти каждое стадо может представлять некоторые особенности. К тому же все видовые отличия принимаемых (разными авторами в разном числе) видов настолько нерезки и со времени одомашнения овец, могли даже несколько измениться, что в настоящее время уже невозможно решить, какие же из этих видов и в какой степени примешались к генофонду домашних овец. В Америке происходит (незначительное) проникновение в домашних овец американского толсторога. И возможно, что в различной степени, но более или менее все дикие виды имели возможность подметаться к домашней овце, встречаясь с нею хотя изредка в горных районах. Если в новейшие-эпохи, когда, с одной стороны, дикие бараны сильно истреблены, а с другой — домашние овцы сильно изменены отбором, подобные встречи становятся все более и более затруднительными, то в более древние периоды (а овцы одомашнены уже много тысячелетий), когда диких баранов было больше, а домашние еще мало отличались от них, эти встречи могли происходить конечно гораздо легче и чаще.
Нет необходимости входить в подобные же детали относительно всех остальных животных.
Переходя к выводам из современных данных об участии гибридизации в эволюций домашних животных, можно их сформулировать так:
1. Несомненно, что целый ряд — и притом наиболее важных— домашних животных подвергся в течение своей истории гибридизации, представляя, подобно самому человеку, продукт смешения генофондов нескольких видов, образовав то, что систематики называют conspecies. Однако столь же несомненно, что в большинстве случаев эти смешавшиеся виды были ближайшими, очень слабо разошедшимися видами, скорее подвидами и географическими расами (типа природных рас львиного зева, исследованных Бауром, и даже более близкими). Только в немногих случаях (гусь, свинья, собака) смешавшиеся виды представляют собой более ясно диференцированные виды (серый гусь и сухонос, кабан и полосатая свинья, волк и шакал). Однако даже и в этих случаях смешавшиеся виды были настолько физиологически близкими, что их гибридизация происходила совершенно свободно без всякой специальной помощи со стороны человека и без всех специально характерных для гибридов явлений — трудностей осеменения и оплодотворения, пониженной плодовитости и т. д. Поэтому эта гибридизация по своему биологическому характеру почти не отличалась от скрещивания различных пород (субгибридизация).
2. Однако и эта субгибридизация, начавшаяся с древнейших времен и идущая и до сего времени, носила в основном стихийный, бессознательный характер, не велась во имя достижения четко поставленных целей. И сейчас например в США иногда домашние овцы, заходя в горы, покрываются дикпми горными баранами, и тем самым генофонд Ovis montana продолжает медленно дифундировать в генофонд домашних овец. И сейчас также домашние собаки нередко скрещиваются и с волками и с шакалами, продолжая ассимиляцию и смешение обоих генофондов (т. е. влияя может быть на сами дикие виды). И лишь в немногих новейших случаях мы имеем сознательное и более или менее планомерное применение такой гибридизации, как в случае скрещивания обыкновенных и китайских гусей, зебу с крупным рогатым скотом в Америке и др.
В соответствии со стихийным характером происходившей гибридизации она осуществлялась и на совершенно неудовлетворительном генетическом уровне. Не могло быть конечно и речи о правильном получении F
1, F
2, F
b и других необходимых поколений гибридов, при помощи которых работают например современные гибридизаторы-ботаники, начинающие широко применять подобные же скрещивания географических рас. Подобного правильно поставленного в научном отношении гибридизационного опыта у животных мы не имеем еще ни одного. Поэтому можно смело утверждать, что гибридизационные возможности, предоставляемые даже уже использованными формами, использованы человеком еще далеко не нацело, и поэтому какое-либо скрещивание волка с шакалом или домашних коров с зебу, произведенное на новом техническом уровне, еще сможет дать немалые результаты. Нужно помнить, что при различии между видами по крайней мере в несколько десятков генов (а- именно так расценивает например Баур разницу между видами львиного зева, а у животных они вряд ли слабее) число возможных комбинаций быстро достигает неисчерпаемого многообразия.
При различии в 10 генов число комбинаций — 1 024
в 20 генов — больше 1 000 000
в 30 генов — 1 000 000 000
в 50 генов — 1 000 000 000 000 000
т. е. числа, в миллионы раз превышающие число всех животных, когда-либо виденных человеком на протяжении всей его истории.
3. Еще значительно реже человек применял более отдаленную гибридизацию уже со всеми ее характерными особенностями. Таковы в истории животноводства скрещивания:
лошадь × осел
крупный рогатый скот × як
крупный рогатый скот × бизон
курица × фазан
утка × мускусная утка
гусь × канадская казарка
Наибольшее распространение и крупное хозяйственное значение имела, как известно, гибридизация лошади с ослом, дававшая мулов, — форма разведения, имеющая давность не в одно тысячелетие. Это гибридное скрещивание уперлось однако в бесплодие гибридов мулов и лошаков и не пошло дальше первого поколения. И характерной иллюстрацией непонимания огромных перспектив, открывающихся при переходе от F
1 к F
2 или даже к F
b является то, что даже тогда, когда отмечались случаи плодовитости мулиц и лошачих, эти случаи остались точно не констатированными, а сами плодовитые мулицы и лошаки и их потомство не были использованы для дальнейшего с их помощью перекидывания моста между обоими видами (поэтому в настоящее время мы так и не знаем окончательно, имелись ли действительно реальные случаи такой плодовитости).
В такое же бесплодие уперлись и все три перечисленные, многократно повторенные гибридные сочетания птиц. Это бесплодие, во всех трех случаях полное, и появлявшиеся сообщения о плодовитых гибридах фазанов × кур несомненно ошибочны.
Более поучительна гибридизация крупного рогатого скота, т. е. говоря точнее, домашнего тура с бизоном (Boyd, 1914) и яком. Гибридизация с бизоном в довольно широких масштабах проводилась в США в XIX в., причем гибриды ценились за их выносливость по отношению к степным зимним буранам. Гибридизация эта имела место на более примитивном уровне американского скотоводства, и какой-либо систематической работы по освоению ценных элементов генофонда бизонов путем гибридизации не велось. Когда же США стали заполняться первоклассными европейскими породами (голландцы, шортгорны, герефорды и др.), то в условиях капиталистической конкуренции эта гибридизация оказалась ликвидированной, тем более что она была конечно значительно сложнее, чем гибридизация с зебу в пироплазматических районах.
Гибридизация с яком в Центральной Азии носила также конечно стихийный характер. Дать однако оценку ее результатов сейчас невозможно, так как необходимы значительно более детальные исследования для того, чтобы сказать, в какой степени стада крупного рогатого скота — тура — этих областей восприняли в себя элементы генофонда яков. Возможно, что этот стихийный процесс прошел гораздо дальше чем сейчас эта представляется, так как при бесплодии самцов F
1 самки F
1, F
2 и пр. плодовиты, и на протяжении веков таких гибридов дальнейших поколений получалось конечно значительное количество (ср. Schlagintweit, 1857).
4. На основании данных о происхождении различных домашних животных и их относительного многообразия сделать вывод о значении гибридизации в этом многообразии в настоящее время мы не можем. Правда, если ограничиться одними млекопитающими, то подобный вывод напрашивается, так как наиболее многопородными и разнопородными домашними млекопитающими являются как раз наиболее гибридизированные (собака, овца, корова, лошадь, свинья) в противоположность сравнительно однообразным монофилетическим видам (верблюды, ламы, северный олень, кролик, морская свинка, кошка), и среди подвергшихся гибридизации наиболее многообразной оказывается собака, гибридизированная паиболее сильно вплоть до участия двух родов. Однако необходимо иметь в виду, что первая группа принадлежит к числу древнейших животных, одомашненных человеком, а собака является наиболее-, древним. Чем. древнее прирученное животное, тем очевидно в течение большего срока работает с ним человек, тем, ceteris paribus, большего многообразия он смог достичь. Это соображение имеет очень существенное значение. Но не все объяснимо и этой ссылкой. Верблюд тоже древнее животное, но он не типичен и для монофилетических видов, ибо до сего времени не вполне ясно, являются ли одно-и двугорбые верблюды породами одного вида или двумя видами. Гибридизация между этими формами несомненно происходила, хотя в новейшее время верблюдоводы и останавливают ее преимущественно на первом поколении.
 Рис. 3. Мулы из США.
Рис. 3. Мулы из США.
Несомненную роль играла и широта распространения предков первой группы видов. Дикие виды собачьих — космополиты, и человек, расселяясь, мог не только привозить одомашненную собаку с собой, но и в случае нужды акклиматизировать ее с помощью скрещивания с местными видами. Предки коровы водились как под тропиками (предки Bos indicus), так и в умеренном климате (В. primigenius). Наоборот, и дикий олень и верблюды имеют сравнительно узкие ареалы и не могли поэтому всюду следовать за человеком, не могли превратиться в основных домашних животных.
Гибридизация в капиталистический период
Гибридизация в капиталистический период характеризуется: 1) чрезвычайно низким удельным весом ее в животноводстве, где однако наряду с продолжающейся местами стихийной гибридизацией появляются отдельные новые случаи гибридизации целеустремленной, более или менее научно поставленной, и 2) широким развитием научной и «любительской» гибридизации животных.
Что касается гибридизации хозяйственной, то кое-где она продолжает стихийно протекать, преимущественно в районах наиболее отсталых. Так, в Центральной Азии, в районе распространения яка, более или менее стихийно проходила гибридизация яка с крупным рогатым скотом, будучи однако уже осознана именно как гибридизация. (Гибриды имеют специальные названия и оцениваются особо, до некоторой степени различаются поколения гибридов.) Что здесь однако дело идет еще о почти стихийной гибридизации, видно из того, что человек еще очень слабо может регулировать ее (что обнаружилось например в виде усиления гибридизации после прошедшей на Алтае коллективизации благодаря простому перемешиванию стад). Примеры такой же, но более скромной по масштабу гибридизации, идущей в наше время — собаки с волками, с шакалами, домашних овец с дикими, — мы уже приводили. Интересно, что почти так же стихийно, как в седой древности, на наших глазах проходит гибридизация двух новых одомашниваемых животных — страуса и охотничьего фазана.
В процесс одомашнивания страуса вовлечено не менее двух видов (Strutio camelus и S. molibdophanes) и в создание паркового охотничьего фазана не менее 4 видов — Phasianus colchicus, Ph. torquatus, Ph. mongolicus и Ph. versicolor) (Cronau, 1902). Если первые три вида фазанов различаются между собой ничтожными признаками окраски или украшением головы, то последний вид окрашен уже совсем иначе.
 Рис. 4. Гибрид фазана и курицы. Изображение 1763 г. (из Поля по Фришу).
Рис. 4. Гибрид фазана и курицы. Изображение 1763 г. (из Поля по Фришу).
В фазанниках Западной Европы (и у нас в Аскании-Нова) эти виды в разных комбинациях настолько перемешалось, что сейчас уже редко где можно найти чистые формы, хотя нередко эта гибридизация ставилась любителями нарочито, но в большинстве случаев носила чисто стихийный характер. Владельцы фазанников, приобретая в разных местах птицу для разведение, мало различали эти виды, свозили в один парк несколько видов, не изолировали их, и в результате в каждом фазаннике создались свои собственные гибридные популяции, при наличии которых уже стало невозможным избегать дальнейшей гибридизации даже в тех случаях, когда ее пытались избегать.
Из старых форм сознательной гибридизации сохранилось и даже пышно расцвело (Франция, США) муловодство. Оно прогрессировало однако лишь в том отношении, что была проведена значительная селекционная работа с ослами и лошадьми, благодаря чему в этих странах мулы поднялись до превосходного качества (рис. 3).
Интересно возникновение «муловодства» в области птицеводства. Последние столетия дали два примера этого применения гибридизации, кратко упомянутой выше: фазан × курица в Европе и гусь × казарка в Америке. Обе эти гибридные формы совершенно бесплодны и поэтому могут иметь только мясное значение для хозяйства. Но зато их мясные качества оказались выше мясных качеств чистых родительских форм. Возможно, что это стоит в связи именно с бесплодием этих гибридов, являющихся до некоторой степени естественными каплунами.
В помещичьих хозяйствах, особенно В; XVIII в., гибриды фазанов с курами разводились нередко (рис. 4). Впоследствии однако эта отрасль сошла напет по той простой причине, что, с одной стороны, получение этих гибридов было очень, невыгодно, так как фазаны давали с курами очень низкий процент оплодотворенных яиц, а с другой стороны, значительно прогрессировали мясные качества самих кур.
 Рис. 5. Канадская казарка.
Рис. 5. Канадская казарка.
В последнее время в Америке распространилось получение гибридов домашних гусей с диким, частично одомашненным американским гусем с канадской казаркой (Brenta canadensis, рис. 5). Этот сравнительно крупный гусь вместе с его более мелкими евразийскими родственниками — белошеей и черной казаркой — относятся совсем к иному роду гусей Brenta. Тем не менее казарка довольно легко скрещивается с домашними, особенно с китайскими гусями, ведущими начало от сухоноса (Cygnopsis cygnoides). Эта комбинация представила выгоды в двух отношениях: во-первых, китайские гусыни отличаются исключительной яйценоскостью, давая до 80 яиц против нескольких яиц, сносимых самкой казарки; с другой стороны, по мясным качествам эти гибриды ценятся на американском рынке значительно выше китайских гусей.
Наибольшее значение в последнее десятилетие имела получившая широкую известность и в научной литературе гибридизация домашнего скота с зебу (рис. 6) в пироплазматических районах США, в Ю. Америке и Сев. Африке. Здесь например по берегам мексиканского залива пироплазмоз свирепствовал настолько, что значительные районы оставались совершенно недоступными для скотоводства несмотря на подходящие климатические и экономические условия (Nabours, 1913). Тогда американцами было ввезено сюда несколько быков зебу из пироплазматических районов Индии. Скот зебу несмотря на то, что его мясные и молочные достоинства значительно ниже лучших американских пород, оказался необыкновенно ценным вследствие своей иммунности к пироплазмозу.
 Рис. 6. Бантенги (слева) и зебу (в центре и справа) в степи Аскании-Нова.
Рис. 6. Бантенги (слева) и зебу (в центре и справа) в степи Аскании-Нова.
Относительно причин этой иммунности в настоящее время мнения расходятся и даже оспаривается иммунность зебу (Якимов, 1932, 1934). Но-несомненным явилось то, что не только сами индийские зебу, но и их гибриды с домашним скотом оказались способными жить в пироплазматических районах. И например по берегам Мексиканского залива в короткое время были созданы значительные массивы этих гибридов. К сожалению, что характерно для капиталистического хозяйства, вокруг этого вопроса создалась рекламная шумиха, усиленно раздувавшаяся звероторговыми фирмами, преимущественно в Германии, транспортировавшей значительное количество зебу в Ю. Америку, и поэтому не всегда легко учесть, где кончаются действительные плюсы этой гибридизации и где начинается реклама.
Сама гибридизация проходила без необходимого участия научных сил, и хотя ей и посвящен ряд работ в генетической и зоотехнической; литературе, но должного освещения ее мы все же не имеем. Несомненно, что и в селекционном отношении работа шла на недостаточной высоте, протекая преимущественно в экстенсивных хозяйствах… В этом отношении могла бы представлять значительно больший интерес новейшая гибридизация зебу молочными породами (Hammond, 1931). Также интересна была работа с яком на Аляске, начатая на опытной станции, но она ведется в ничтожных масштабах, а сейчас в связи с кризисом вероятно ликвидирована вместе со станцией.
Если таким образом в области хозяйственной гибридизации животных капиталистический мир дал очень мало ценного, то значительно большие результаты мы находим начиная с XIX в. и в XX в. в области научной гибридизации. С результатами этой работы нам предстоит познакомиться в следующих главах. Здесь мы отметим лишь общие черты этой работы.
Научная гибридизация
Оторванная от проблем практического животноводства научная гибридизация шла в следующих направлениях:
1) изучение естественной гибридизации (в природе), проводимое главным образом систематиками и фаунистами;
2) экспериментальная гибридизация в связи с проблемой происхождения домашних животных (Кюн в Галле и Др.);
3) гибридизация, большей частью случайная и бессистемная, в зоологических садах, дававшая обильный материал для музеев, отмечавшийся и в научной литературе;
4) любительская, незаметно однако переходящая в научную, гибридизация в парках, птичниках, аквариумах, давшая, с одной стороны, богатый материал для научной литературы (с огромным ба-ластом ошибок и вздора), а с другой стороны — живой гибридный материал вроде упомянутых гибридов фазанов, мышей (в наших лабораторных мышах смешаны не менее двух видов Mus: Mus niusculus и Mus wagneri) и некоторых аквариумных рыбок;
5) многочисленные, начиная с 80-х годов экспериментальные работы по отдаленной гибридизации, проводившиеся под углом зрения проблем механики развития, теории оплодотворения и пр.;
6) многочисленные, большей частью экспериментальные гибридизационные работы энтомологов, преимущественно лепидоптерологов, получавших гибридов бабочек для своих коллекций (а также для обмена и даже торговли), частью однако повышавшиеся до работ высокого научного значения (напр. Штандфусса и др.);
7) наконец также уже довольно многочисленные работы по гибридизации, проведенные генетиками и исходившие из задач разработки современной теории наследственности, сравнительной генетики, проблемы пола, цитологии гибридов и пр.
Все эти многочисленные работы накопили огромный фактический и теоретический материал, уже с трудом поддающийся обозрению. Таковы десятки первоклассных экспериментальных работ, печатавшиеся преимущественно в Archiv für Entwicklungmechanik, посвященные гибридизации иглокожих (см. главу V). Работы по естественной и искусственной гибридизации рыб, птиц, бабочек и др. насчитываются сотнями. Не менее многочисленны работы по гибридизаций домашних животных. И наконец нужно иметь в виду, что для освещения многих вопросов гибридизации животных крупнейшее значение имеет и огромная литература по гибридизации растений.
В процессу этих работ были получены многие сотни различных гибридных комбинаций (ср. по птицам Hopkinson, 1926). К сожалению однако изучение этих гибридов за редкими исключениями (Штандфусс и другие энтомологи) было совершенно неудовлетворительно. В качестве яркого примера можно сослаться на пользовавшихся наибольшим вниманием гибридов птиц. Несмотря на то что число гибридов птиц, попавших в руки ученых, огромно, их исследование ограничивалось описанием окраски и монтированием шкурок и чучел. Лишь с трудом находим 1–2 описания скелетов (Schufeldt, 1890; а по млекопитающим Noack, 1908, Филипченко, 1916) и уже вовсе не находим более детального изучения даже анатомии, не говоря о физиологии и пр. Даже в работах Поля, Гайера и др., давших очень интересные исследования, разбирались лишь отдельные вопросы (атавизм, причины бесплодия и некоторые другие).
Вторым крупнейшим недостатком этих работ с точки зрения, разбираемой в настоящей книге, является то, что несмотря на плодовитость многих из получившихся гибридов капиталистическая наука и хозяйство оказались неспособными их сохранить в виде живых гибридных линий. В богатой Англии например Бонхоту (Bonchote, 1902–1908) удалось получить несколько поколений гибридов утки и шилохвости. Генофонды этих двух видов оказались слитыми, но ничего кроме статей и музейных шкурок от этого крупного достижения не осталось. Ничего не осталось от получившихся плодовитых гибридов кавказского фазана и серебристого, откуда несомненно можно было бы почерпнуть много ценного для птицеводства. Ничего не осталось в домашнем состоянии от интереснейших плодовитых гибридов домашних коз с диким альпийским козерогом — видом, несомненно не принимавшим участия в образовании домашних коз; гибридов крупного рогатого скота с бизонами и т. д. Наконец ничего живого не осталось от интересных гибридологических работ Кюна в Галле и Юарта в. Шотландии. Капиталистическому миру оказалось не под силу сохранить эти не дававшие прибыли достижения своей науки, в то время как СССР в труднейшие годы гражданской войны сумел спасти и сохранить наиболее ценных, хотя иногда также еще не представляющих непосредственной хозяйственной ценности гибридов Аскании-Нова.
Гибридизация в СССР
До революции гибридизация животных в России носила тот же характер, как и, только что описанная работа в других капиталистических странах, только в еще более скромных масштабах. В капиталистическом животноводстве она не играла никакой роли, если не считать некоторых процессов гибридизации домашнего скота с яком на Алтае и в Киргизии, проникновение элементов зебу в Закавказье и Ср. Азию и получение гибридов двугорбого верблюда с одногорбым, лошади с ослом. Известна (впрочем из вторых уст) скромная попытка завоза зебу в пироплазматический район Сухума одним помещиком. Проблемы гибридизации в науке были представлены тоже очень скромно. Такие направления биологии, как механика развития, генетика, стали проникать в Россию лишь незадолго до войны. Правда, еще, Паллас обсуждал роль гибридизации в происхождении собак, а в 1933 г. исполнилось 50-летие небольшой статьи Н. А. Северцова (Severzow, 1883), описавшего (с цветной таблицей) гибрида утки с чирком и обсуждавшего некоторые вопросы гибридизации, а еще ранее в Ornis Caucasica Радде (Radde, 1884) (тоже с цветной таблицей) описал застреленного в 1873 г. на воле в Закавказье гибрида домашней утки с мускусной. Однако какого-либо большого, научного значения эти и подобные им работы не имели.
Интересно все же вспомнить, что в 1857 г. Врасский, основатель Никольского рыбоводного завода, открывший так наз. «русский» или сухой способ оплодотворения икры рыб, почти немедленно применил его к опытам гибридизации рыб — повидимому впервые. Эти опыты также позднее продолжал Гримм (1881). В 1869 и 1870 гг. в Петербурге нашумели гибридизационные опыты с рыбами Кноха, частью оказавшиеся ошибочными (ср. Богданов, 1883).
 Рис. 7. Зеброид (гибрид зебры и лошади Пржевальского) с матерью-зеброй в Аскании-Нова.
Рис. 7. Зеброид (гибрид зебры и лошади Пржевальского) с матерью-зеброй в Аскании-Нова.
Довольно значительный интерес представили также опыты по гибридизации, начатые с конца XIX века Ф. Э. Фальц-Фейном в его крупном поместье на Украине — Аскании-Нова. Среди других помещиков, заводивших по примеру Западной Европы себе зоологические парки (вроде полудегенерата А. С. Хомякова, устраивавшего в своем имении под Тулой соревнования в беге между крестьянскими парнями и страусами), Фальц-Фейн выделился более серьезной постановкой работ по гибридизации. В основном и она носила любительский характер, особенно по птице, где были получены некоторые очень интересные гибриды, вроде полярного гуся с серым и др. и спрятаны в музей без всякого описания, — но особое внимание уделялось все же гибридам лошадей с зебрами (рис. 7) и домашнего скота с зубрами и бизонами (рис. 8), по отношению к которым обсуждалась проблема их хозяйственного значения. Кастраты гибрида с бизонами испытывались в качестве рабочих волов, причем были обнаружены их выдающаяся сила и выносливость; зеброиды ходили в упряжи. Наибольший интерес представили первые попытки применения для целей гибридизации искусственного осеменения, благодаря которым были получены некоторые зеброиды. Исследовались причины бесплодия гибридов, краниология гибридов (Иванов, Филипченко). И хотя в конечном счете масштаб этих работ был очень скромен и за 20 лет работы было получено например лишь десятка два гибридов тура с зубрами и бизонами, тем не менее эти работы выделились на фоне мировой науки и привлекли широкое внимание. Еще не успев превратиться после революции в советское хозяйство, Аскания-Нова попала в фронтовую обстановку гражданской войны, многократно переходя из рук в руки. Естественно, что она при этом сильно пострадала — разными белыми бандами был захвачен скот, угнаны гибриды-зеброиды и т. д. Однако в самый разгар военных действий советская власть приняла меры к спасению ценностей Аскании-Нова. Началось восстановление, а затем сильное расширение зоопарка и хозяйства. Из гибридов были сохранены группы зубро-бизоно-туров, оленей, муфлонов и фазанов. При недостаточной систематичности, а часто и случайности проводившейся гибридизации не все эти группы имели одинаковое значение, тем более что погибла и часть записей. Но для гибридизационных работ не меньшую ценность представили спасенные группы диких животных — лошадей Пржевальского, антилоп-канн, гну и др.
 Рис. 8. Зубро-бизоны в Аскании-Нова.
Рис. 8. Зубро-бизоны в Аскании-Нова.
В последующие годы по мере восстановления хозяйства началась организация и научной работы. Сменив несколько раз организационную форму, Аскании-Нова превратилась в 1931/32 г. в Институт гибридизации и акклиматизации животных, с крупным бюджетом и многочисленным штатом научных и технических работников. Вместе с тем совершенно иные масштабы приняла и работа по. гибридизации животных, достигнув невиданного размаха. Достаточно сказать, что в 1932 и 1933 гг. в одних лишь опытах по гибридизации тура, зебу и бантенга участвовало несколько сот коров как в самой Аскании, так и в группе окружающих ее совхозов.
Помимо Аскании. работы по гибридизации ведутся или организуются и в других местах. Опытная станция возле Улалы (Ойротия, Алтай), правда, еще слабая, в значительной степени посвящена яку (рис. 9) и его гибридам; аналогичные работы ведутся в Киргизии (Любимов и др.). В Московском зоопарке проводилось в течение ряда лет скрещивание зебу × як (Завадовский). Гибридов яков на месте обследовали Лус, Гершензон, Волохов, Власов, Поляков, Кажарин и др. По гибридизации птиц работает кафедра генетики МГУ в Москве и Аскании, где с 1932 г. были получены искусственным осеменением гибриды кур с павлином, цесарок с курами, гибриды уток и др. Отдельные работы ведутся и в других местах.
Основные вехи работ по гибридизации сформулированы в известном постановлении (по инициативе наркома Я. А. Яковлева) Коллегии НКЗема СССР от 27–29 сентября 1931 г. (см. Приложение к главе), явившемся крупным стимулом для развития гибридизации в СССР, и в резолюциях Всесоюзной конференции по планированию генетикоселекционных исследований во второй пятилетке в июне 1932 г.
Развернутое строительство социализма в СССР, в частности ликвидация частнохозяйственной стихии в животноводстве и создание крупнейшего в мире совхозно-колхозного животноводства, теснейшая связь, установившаяся между животноводством и наукой, открывает перед гибридизацией совершенно новые перспективы.
 Рис. 9. Яки (Берлинский зоосад).
Рис. 9. Яки (Берлинский зоосад).
Действительно, необходимым условием для серьезного и эфективного развертывания работ по гибридизации животных являются именно 1) возможность оперирования крупными группами животных в течение достаточно долгого ряда лет, на протяжении которых работы эти могут не давать непосредственного народнохозяйственного эфекта, 2) непосредственное участие зоотехнической и биологической науки в этих крупных и сложных работах. Поскольку только в социалистической обстановке, где животноводство, быстро крепнущее, освобождено от кризисов и направлено на производство необходимой продукции, а не прибыли и где крупнейшие ученые считают для себя делом чести участие в соцстроительстве и содействие ему— эти необходимые условия реализованы, — постольку мы и должны считать перспективы гибридизации небывало благоприятными. Естественно, что эти перспективы будут тем шире, чем больше будет крепнуть и развиваться наше животноводство.
Нам остается более детально обсудить эти перспективы.
Гибридизация и освоение мировых зоологических ресурсов
Несмотря на то что человек издревле широко и многообразно использует животных путем охоты и рыболовства и путем животноводства, необходимо констатировать, что это использование, еще чрезвычайно слабо в видовом отношении и примитивно. Это положение вытекает из следующих соображений. Несомненно, что действие тельно эфективное и рациональное использование достигается лишь тогда, когда человек вполне овладевает данным видом организма, одомашнивает его, начинает рационально кормить и размножать его, совершенствует (повышает его продуктивность) путем селекции и пр., начинает лечить его и вообще снижать те огромные потери, которые каждый вид несет в естественном состоянии. И если сейчас корова или курица превращены в мощные «фабрики» или «аппараты» для продукции молока и яиц, далеко превосходящие по своей производительности своих диких предков, то мы вправе с удовлетворением сказать, что мы действительно научились использовать данное животное.
И здесь нам приходится прежде всего констатировать количественно (по числу видов) совершенно ничтожные успехи человечества за всю его историю в деле одомашнивания, особенно если принять во внимание колоссальный видовой состав животного мира.
Если сравнить число используемых видов животных с числом видов растений, которые серьезно используются человеком в качестве культурных, то различие становится особенно разительным. Только в СССР ботаники ведут селекционную работу не менее чем со 100 видами растений. Количество видов растений, охваченных культурой, исчисляется многими тысячами. И это в то время, когда видовой состав растений неизмеримо скромнее состава животных. И уже совершенно ничтожно одомашнение животного мира помимо млекопитающих и птиц.
Правда, помимо одомашненных человек использует много диких видов путем охоты и рыболовства, однако этот метод использования можно сопоставить с использованием растений путем сенокоса, заготовкой дров, сборами диких ягод и орехов, водорослей, выброшенных морем, сбора лекарственных трав и пр. Но при таком счете и список используемых растений расширится почти до пределов всего растительного мира.
Весьма интересно, что как раз в капиталистическую эпоху различие между освоением животного и растительного мира стало особенно сильным. В то время как в области ботанической идет интенсивная работа по интродукции и во всяком случае по распространению по земле хотя бы и старых, но узкоместных культур, в области освоения животного мира мы имеем совершенно ничтожные успехи. Из числа новых видов, одомашненных человеком за последние столетия, можно назвать лишь канарейку, страуса, мускусную утку, канадскую казарку и еще ряд декоративных воробьиных птичек, фазанов и декоративных уток, степень одомашнения которых еще очень ничтожна. Из млекопитающих можно назвать опоссума, лисицу, нутрию. Из рыб — декоративных аквариумных рыбешек, из которых лишь о Lebistes и гамбузии, используемых в борьбе с малярией, можно говорить как о видах, начатых освоением. Ничего значительного таким образом капиталистическая эпоха здесь не дала. Оставив без внимания задачу освоения нового, она занялась усовершенствованием путем селекции основных сельскохозяйственных видов животных — корова, овца, свинья, где и дала блестящие результаты.
Какова. причина столь ничтожного охвата доместикацией видового состава животных, особенно по сравнению с растениями? Значит ли это, что животный мир не представляет больше никакого интереса для
доместикации и для других форм аналогичного использования их человеком? Конечно не значит.
Причины слабого освоения животного мира разнообразны и серьезны.
Прежде всего огромное значение имеют чисто технические трудности разведения животных, незнакомые растениеводам, которые имеют возможность за сравнительно редкими исключениями (орхидеи, грибы) без особых технических приемов разводить и размножать любые дикие растения. Даже климатические припятствия и те сравнительно легко, по крайней мере в известном диапазоне, преодолеваются при разведении растений. Наоборот, для животных возможность их разведения в домашних условиях является скорее исключением, чем правилом, и огромное количество видов даже в условиях зоологических садов либо вовсе не размножается либо размножается в виде редких случаев, регистрируемых литературой, благодаря чему животный мир зоосадов приходится непрерывно пополнять поимками новых диких животных. Немногие исключения (антилопа-канна, гну, фазаны, волнистые попугайчики, аквариумные рыбки и пр.) очень мало нарушают общую картину, и например недавние “достижения СССР- в области разведения соболя показывают, что разрешение задачи для каждого вида требует значительной исследовательской работы.
Отсюда не следует конечно, что человек, уже в древности одомашнивший десятка два видов, впоследствии
утратил свое искусство. Наоборот, это указывает, что распространенное мнение о том, что человек сознательно избрал ряд нужных ему диких видов и одомашнил их, требует решительного пересмотра. Новейшее изучение вопроса показывает, что в процессе одомашнения значительную роль играли и сами одомашниваемые животные, их биологические особенности. Даже по отношению к растениям старых культур все более обосновывается мнение, что в значительной степени они сами вошли в культуру: «редька влезла в огород», овес вытеснил пшеницу при продвижении на север, внедрившись первоначально в посевы в качестве злостного сорняка и утратив затем в процессе почти естественного отбора ломкость колосков и превратившись в культурное растение. Точно так же одомашненными оказались только те животные, которые обладали для этого специальными биологическими предпосылками. Не случайно, что например все одомашненные животные являются стадными и полигамными видами. С точки зрения интересов человека например дикие лисицы и дикие шакалы по крайней мере равноценны, вернее даже лисица представляет больший интерес и как пушное и как охотничье животное. А тем не менее шакал, назойливо «влезающий» в лагери путешественников, гораздо более общительный и легко приручающийся, оказался действительно одомашненным уже много тысячелетий назад, а лисица лишь в самое последнее время и с большими техническими затратами и трудностями волка в хозяйство человека, да и сейчас далеко еще не может быть названа действительно одомашненной.
Как увидим ниже, это обстоятельство придает сразу огромное значение гибридизации как
методу освоения зоологических ресурсов, так как, гибридизируя уже одомашненные виды с трудно поддающимися одомашнению дикими, мы можем либо облегчить доместикацию, либо использовать генные ресурсы диких видов, минуя необходимость доместикации последних.
Эта техническая причина однако не является решающей. Она безусловно являлась решающей в древние эпохи, но в новейшее время, с развитием науки и техники, человек несомненно уже имел бы возможность преодолеть это препятствие по отношению по крайней мере ко многим видам животных. Пример лисицы, страуса, канарейки подтверждает это. И если мы видим, что именно в эпоху вооруженного техникой и наукой капитализма освоение зоологических ресурсов путем доместикации имеет ничтожное развитие, то основными причинами здесь являются причины уже иного, социально-экономического характера.
Животное по сравнению с растением является гораздо менее выгодным путем освоения солнечной энергии, так как оно перерабатывает растения, усваивая лишь небольшую долю энергии. Лишь в особо благоприятных случаях для наиболее культурных пород домашних животных мы имеем коефициент 1:3; обычно же он гораздо ниже. Поэтому животное оказывается выгодным лишь тогда, когда дает особенно ценный продукт — мясо, яйца, шерсть, тяговую силу и пр., которые нельзя или трудно заменить растительными продуктами.
Вместе с тем особое значение приобретают для судьбы животного те успехи, которые достигнуты в повышении его продуктивности. Если лучшие породы свиней усовершенствованы человеком настолько, что могут дать 1 кг мяса за 3 кг корма, то конкурировать со свиньей какому-либо новому, только что одомашненному виду по линии мясной продукции уже совершенно невозможно. Если курица может давать сейчас свыше 300 яиц в год, а в виде цыплят и огромную мясную продукцию, то конкурировать с ней какой-либо дрофе, если бы мы ее одомашнили, дающей 1–2 яйца в год, уже невозможно. Это и приводит к тому, что в эпоху капитализма оказались одомашненными животные или дававшие совершенно новый продукт, не имевший конкурента, например страус, или дававшие продукт, который мог конкурировать по разным другим причинам, как например путные животные (лиса, опоссум, нутрия).
Капиталистическое животноводческое хозяйство, как и всякая иная отрасль капиталистического хозяйства, в высокой степени заинтересовано в норме прибыли. В сельском хозяйстве нормы прибыли вообще ниже, чем в промышленности, и повышения этой нормы капиталист добивается различными искусственными способами. В племенном животноводстве мы видим особенно ясные результаты этих стараний. Различного рода реклама, мероприятия в виде племенных книг, выставочных премий и чемпионатов, в виде стандартов, которыми объединение разводчиков какой-либо породы ограждает свою монополию от других хозяев, приводят к тому, что выпускаемая ими продукция в виде племенных чистопородных животных имеет стоимость, значительно превышающую действительную их стоимость, отвечающую количеству затраченного на их производство необходимого труда, что и дает возможность существования племенных животноводческих хозяйств рядом с промышленными предприятиями. В условиях капиталистической конкуренции на эти спекулятивные цены идут и потребители, так как, приобретая за непомерно вздутую цену какого-либо выставочного чемпиона, более или менее независимо от его действительных племенных достоинств, этот новый владелец вписывает имя чемпиона в свои проспекты и в родословные своего молодняка, получая возможность и его продавать также по значительно вздутым ценам.
Естественно, что в этих условиях все внимание капиталиста-животновода, производящего племенной материал, сосредоточивается на разведении лишь первоклассных чистых пород, при работе с которыми норма прибыли оказывается наибольшей, и эта тенденция становится тем сильнее, чем больше обостряется борьба за прибыль. Число разводимых пород все больше и больше сокращается. Разведение не только беспородных, но даже малопопулярных пород, если бы даже они были не хуже мировых пород, скрещивание пород в надежде или даже в правильном расчете на создание новой породы, а тем более всякая гибридизация, ощутительные результаты которой могут быть получены лишь через ряд поколений, — все это становится для капиталистического животноводства уже недоступным, и с каждым годом все недоступнее (кроме таких отдельных исключительных случаев, как например случай с зебу, давший эфект уже в первом поколении).
Сказанное необходимо иметь в виду, чтобы правильно отвечать на вопрос о том,
не исчерпал ли себя уже метод гибридизации? Если иметь в виду, что, с одной стороны, начиная с древнейших времен, гибридизация сыграла крупную роль в создании наших важнейших домашних животных, а с другой стороны, что последние века ничего существенного в этом отношении уже не дали, такой вопрос является вполне законным. Но достаточно, с одной стороны, понять, что условия капиталистического животноводства оказались тормозящими гибридизацию, а с другой стороны, вспомнить, что в растениеводстве, в котором благодаря дешевизне материала этот тормоз отсутствует, гибридизация в последние столетия получила, наоборот, сильнейшее развитие, с каждым годом все усиливающееся, чтобы ответ на поставленный вопрос стал ясен: ни о каком исчерпании метода гибридизации животных не может быть и речи, но просто капиталистическому хозяйству он не под силу. И на протяжении тысячелетий гибридизация животных не только не исчерпана, но осталась едва лишь затронутой.
Однако необходимо подчеркнуть, что блестящие достижения капиталистического племенного животноводства, поднявшие качества домашних животных на огромную высоту, создали для гибридизации известные трудности, которые сохраняются и при переходе к социалистическому животноводству. Нас теперь интересует не прибыль, а производительность труда. Как бы ни были высоки качества наших нынешних животных, но они не идеальны. И поэтому естественен вопрос, что для пас целесообразнее — продолжать ли дальнейшую работу по улучшению наших лучших животных или заниматься созданием новых пород или даже видов домашних животных с применением гибридизации. Если бы вопрос стоял так, то надо было бы ответить определенно: необходимо дальше улучшать лучшее. Действительно, будет безнадежно глупо, если, имея шортгорнов и симменталов, мы начнем скрещивать дикие виды быков и антилоп в расчете «догнать и перегнать» качества этих пород. Однако вопрос так вовсе не стоит, во всяком случае должен быть в большинстве случаев поставлен вовсе не так. Необходимо работу по улучшению лучших построить так, чтобы в ней нашли применение все возможные средства, в том числе и гибридизация. Не антилопами пытаться заменить скот, но гибридизацией бантенгов, зубров, антилоп
с первоклассными породами скота добиваться либо создания совершенно новых форм либо улучшения нынешних видов, памятуя твердо, что улучшить лучшее можно не только при помощи еще более лучшего, но и при помощи «худшего», если в этом худшем имеются отдельные высокоценные свойства или даже гены. Хотя в общем зебу и хуже первоклассных пород обычного скота, но если из их генотипа мощно будет выделить например ту часть, которая повысит иммунность домашнего скота, это будет несомненным улучшением последнего. Золотая рыбка вообще менее вынослива к нашему климату, чем карп, но если верен упомянутый ниже опыт (стр. 43), то гибриды карпа с золотой рыбкой оказались выносливее к суровой зиме, чем даже карпы, что генетически объяснимо совершенно легко. Отсюда крупнейшей задачей гибридизатора является отыскание среди множества диких видов отдельных интересных свойств, хотя бы мелких черточек, аналогичных генов и тому подобного ценного материала, который можно было бы путем гибридизации использовать для дальнейшего улучшения наших животных.
В значительной степени этим же путем идут и гибридизаторы-ботаники, давшие уже много очень ценных результатов, преимущественно в области садоводства и плодоводства, но в последнее время внедряющие этот метод и в другие отрасли растениеводства. Всем: известны имена Бербанка (США) и Мичурина, которые путем часто-самых рискованных скрещиваний добивались блестящих результатов. Обычно одним из родителей, вводимым в скрещивание, является ценное культурное растение, а другим — дикий вид, интересный в каком-либо одном отношении (например сильная морозоустойчивость некоторых диких сибирских видов, использованных Мичуриным; кактус, мелкий и невзрачный, но лишенный шипов у Бербанка и т. п.). Первые поколения гибридов обычно бывают мало интересными в хозяйственном отношении, т. е. качества культурного родителя более или менее резко снижаются (например известный редично-капустный гибрид Карпеченко не имеет ни утолщенного корня редьки ни кочана капусты). Но во втором поколении, если его удается получить, и притом в достаточном объеме, наступает комбинирование признаков обоих родителей, в процессе чего и могут быть извлечены особи, соединяющие ценные качества культурного родителя с ценным специальным признаком дикаря.
Ценный культурный сорт оказывается улучшенным при помощи «невзрачного» дикаря.
Применение метода гибридизации
Применение метода гибридизации для разрешения народнохозяйственных проблем может быть весьма разнообразным во всех тех случаях, когда возникает необходимость в резком повышении наследственной изменчивости животных. Можно различить следующие разделы хозяйственной деятельности человека, в которых такая потребность имеется налицо: 1) при доместикации животных, 2) при акклиматизации, 3) при селекции и реконструкции домашних и других разводимых животных и 4) при воздействиях на взаимоотношения животных видов друг с другом, с растениями и микроорганизмами.
Гибридизация и доместикация
Гибридизация открывает возможность двоякого воздействия на доместикацию. Для форм, ближайших к нашим домашним видам, но трудно поддающихся доместикации (вроде лисицы и т. п.), можно ставить задачу путем гибридизации с одомашненными, добиться устранения этих инстинктивных, и других препятствий введением в трудно одомашниваемые виды тех частей генотипов домашних видов, которые облегчают процесс доместикации. С другой стороны, можно использовать повышение наследственной изменчивости в F
2 гибридов в расчете на то, что при скрещивании двух диких видов в F
2 произойдет наследственное расшатывание их рефлекторных свойств, возникает многообразие, которое сделает возможным из этого многообразия выбрать формы, уже легче одомашниваемые.
Хотя конкретных зарегистрированных примеров этой роли гибридизации в истории одомашнивания привести и невозможно, но можно думать, что в процессе создания например наших ручных овец от диких баранов
[7] такое «расшатывание» в результате гибридизации могло сыграть свою роль. Очень важным указанием на это служит нередко отмечавшееся гибридизаторами сильное повышение «дикости» гибридов, указывающее на разницу в соответствующих частях генотипов скрещиваемых видов и позволяющее тем самым рассчитывать на получение форм, наоборот, менее «диких» (см. гл. IX). К сожалению относящиеся сюда наблюдения производились недостаточно точно и объективно.
Гибридизация и адаптация (акклиматизация)
При перенесении животных в новые для них условия климата и пр. для их филогенетической адаптации должен пройти известный процесс естественного отбора, который отобрал бы из имеющихся в популяции различных экотипов те именно, которые наиболее соответствуют этим новым условиям. Для успешности и для ускорения этого процесса филогенетической адаптации чрезвычайно важное значение должно иметь максимальное повышение наследственного многообразия акклиматизируемого вида, что особенно легко может быть достигнуто именно — методом гибридизации.
В недавнее время например большое значение в борьбе с малярией получило разведение маленькой рыбки. гамбузии (Gambusia), перенесенной в Европу из американских тропиков. В 1924 г. гамбузия привезена из Италии в СССР и успешно распространяется в закавказских водоемах, истребляя личинок малярийного комара. Рыбка эта отличается рядом чрезвычайно ценных биологических свойств, и широкое распространение ее в водоемах СССР было бы чрезвычайно ценным достижением в борьбе не только с малярией, но и с комарами вообще. Однако у гамбузии имеется с этой точки зрения крупнейший недостаток в ее термофильности, не позволяющей ей продвигаться в более холодные районы. Гамбузия размножается при 22°, а при 10° уже зарывается в ил. Снижение ее термофильности на каждый градус открывало бы для нее обширные районы СССР.
Между тем гамбузия относится к семейству зубастых карпов (Cyprinodontidae), очень богатому родами и видами, да и сам род Gambusia тоже богат. видами. К этому семейству относятся такие аквариумные рыбки, как Platypaecilus и Xiphophorus, между которыми любители и биологи многократно получали межродовых гибридов, оказывавшихся плодовитыми и дававшими в F
2 весьма многообразные и своеобразные расщепления, о которых мы будем подробнее говорить ниже. Можно не сомневаться в том, что в этом многообразии не остается незатронутой и термофилия и что отсюда можно будет извлечь формы, выносящие и более низкие температуры. И поэтому можно выдвинуть как вполне реальную задачу гибридизацию гамбузии с другими видами и родами этого семейства с тем, чтобы в результате, снизив термофильность гибридов, дать СССР рыбу-истребительницу комаров, способную жить и в более холодных водоемах.
В семидесятых годах Фишер (Ficher, 1880) получил гибридов карпа (Cyprinus carpio) с золотой рыбкой (Carassius carassius auratus). Эти гибриды оказались менее чувствительными к холоду, чем карпы. В суровую зиму 1879/80 г. погибли все карпы в числе 350 экземпляров, а 300 гибридов остались целы и здоровы. Так как золотая рыбка тоже довольно чувствительна к холоду, то мы имеем здесь поучительнейший пример того, что холодостойкость более северной формы карпа оказалась повышенной благодаря скрещиванию с более нежной формой — случай на первый взгляд пародоксальный, но без труда объясняемый генетически.
Аналогичный пример можно привести и относительно зебу. Ряд свойств зебу (некоторых рас по крайней мере) представляет для нас высокий интерес. Не повторяя сказанного о пироплазмозе, отметим очень высокую. жирномолочность, доходящую до 7–9 % жира, наблюдающуюся в группе коров аравийского зебу в Аскании-Нова (Никольский). Но эти животные с их нежной кожей и слабым волосяным покровом, а может быть и по другом причинам очень зябки и начинают дрожать при температуре, которую свободно еще выносят обыкновенные коровы. Эта зябкость наследственна и сохраняется еще у гибридов F
1. Но поскольку эти гибриды плодовиты, можно будет системой скрещивания с крупным рогатым скотом, а тем более с яком сделать их холодоустойчивыми и дать возможность созданным линиям продвигаться в холодные районы.
Гибридизация при воздействии на взаимоотношения животных
Гибридизация при воздействии на взаимоотношения животных видов друг с другом и с растениями также должна будет найти свое место. В настоящее время например получает все большее развитие биологический метод борьбы с вредными насекомыми, дли чего обычно размножают другие виды насекомых, паразитов данного вредного вида, или хищников, живущих за — его счет (наездников, мух-тахин, божьих коровок, жужелиц и пр.). Однако любая система «хозяин — паразит», существующая в настоящее время, так взаимно адаптирована, что только известная часть хозяина может быть истреблена, а часть все же ускользает от поражения паразитом. Очевидно, что процесс эволюции «нападения» паразита и эволюции «защиты» хозяина по своим темпам совпадают с некоторым отставанием паразита (оставляя в стороне более сложные системы с участием сверхпаразитов, различных хозяев и пр.). Поэтому если бы можно было
ускорить эволюцию паразита, то он смог бы выработать «орудия нападения», более мощные, чем защита хозяина, и истребил бы последнего нацело. Можно думать, что в некоторых случаях гибридизация разных видов паразитов друг с другом, повысив наследственную. изменчивость, смогла бы двинуть быстрее вперед эволюцию паразита и сделать его уже неотразимым для хозяина, подлежащего истреблению из фауны.
В обратном случае можно воздействием на эволюцию хозяина помочь ему выработать более сильные средства защиты против нападения паразита. Приведенный выше пример с гибридизацией в экстенсивных хозяйствах крупного рогатого скота побережья Мексиканского залива с индийскими зебу относится как раз к этой категории, Введение в генотип Bos taurus некоторых наследственных элементов Bos indicus дало первому возможность пройти более быстрым темпом ту эволюцию, которая позволила зебу превратиться в вид, стойкий по отношению к пироплазмозу. Хотя вообще говоря, несомненно, что вызывающая пироплазмоз пироплазма или ее переносчик — клещ — тоже проходят свою эволюцию, постепенно приспособляясь к поражению быков, но в данном примере в филогенезе Bos taurus гибридизацией был произведен такой скачок, который оказался непревзойденным паразитом, и хозяин ускользнул от него, выставив более сильную защиту против нападения оставшегося (пока) на прежнем уровне паразита. Приводя этот пример, оговоримся все же, что некоторые авторы отрицают иммунность зебу к пироплазмозу и пытаются объяснить мексиканский и другие случаи иначе.
Гибридизация для селекции и реконструкции домашних животных
Гибридизация для селекции и реконструкции домашних животных имеет сейчас наибольшие перспективы. Но производить обзор даже главнейших проблем в этой области было бы здесь слишком громоздко и его придется отложить до соответствующих глав частной гибридагогии, посвященных отдельным видам животных. К тому же стоящие здесь задачи могут иметь широкий диапазон от вполне реальных сегодня до почти фантастических, которые при кратком изложении могут быть приняты за пустые фантазии. А между тем пр нашему убеждению некоторый элемент научной фантастики должен быть обязательным в работе гибридизатора, ставящего вопрос о реконструкции животного организма, так как только при этих условиях можно будет серьезно поставить эту проблему.
Разберем такой пример. На наших глазах птицеводство переживает исключительную по глубине техническую революцию. Что осталось — или остается — от романтической наседки с дюжиной пуховых шариков-цыплят и петуха, орущего на плетне? Мощные инкубатории, выбрасывающие в сезон миллионы цыплят, рассылающие за сотни километров по почте и аэропланами однодневных цыплят десятками тысяч ящиков; брудерные батареи-этажерки (рис. 10), где со всех сторон несется писк десятков тысяч цыплят, сидящих в одной комнате, через которую просасывает воздух мощный вентилятор, по которой в тележках развозят корм. И эти цыплята, которые уже никогда не выходят из своих металлических этажерок, порой совершают в них весь свой жизненный цикл! Такому кардинальному изменению форм птицеводства конечно должна была бы отвечать и коренная реконструкция всей птицы. В самом деле, зачем этим этажерочным жителям перья, раз в комнате и без того тепло и не бывает ни дождя ни ветра? Зачем им ноги, раз нм не нужно уж ничего разгребать, можно почти не двигаться по клетке, разве лишь для необходимого моциона? Зачем крылья, зачем уши, зачем голос, звук которого только утомляет рабочих этих фабрик? С другой стороны, развитие и внедрение в селекцию метода искусственного осеменения выдвигает свои новые требования, например в смысле удобства и количества получения спермы. у петухов (пли у самца другого вида) и легкости и быстроты введения ее в организм самки. Даваемый птицей продукт — яйцо — правда, уже близок к совершенству, но почему не может быть поставлен вопрос о том, чтобы курица несла не одно, а два яйца в сутки, а может быть давала бы и какой-либо новый вид продукта, богатые витаминами или гормонами образования, вроде гребешков, которые могли бы быть распространены и дальше, назад вдоль головы и шеи и т. д.
 Рис. 10. Батарейное воспитание цыплят.
Рис. 10. Батарейное воспитание цыплят.
Гибридагогу чрезвычайно важно под этим углом зрения иметь полные сведения о тех возможных направлениях изменений, которые уже свойственны данной группе животных. Если бы у нашего петуха не было гребня, то не так легко было бы представить себе вообще возможность существования подобного образования у птиц. А сейчас, зная, что у петуха имеются гребень и сережки, у индюка своеобразные кожные наплывы и кавернозный придаток над клювом, у фазана разрастающееся образование вокруг глаз и т. д., можно уже строить себе довольно многообразные «фантазии» о том, каким образом может быть реконструирована голова птицы, и притом в полной уверенности, что приемами гибридизации (если будут разрешены стоящие здесь проблемы осеменения, оплодотворения и т. д.) эти «фантазии» смогут быть конечно не только осуществлены, но и перекрыты во много раз. Гибридизатор должен вспомнить, какие удивительные неожиданные видоизменения и «реконструкции» цветов получил Баур при гибридизации двух близких видов Antirrhinum majus и A. molle. Здесь в F
2 возникли формы, которые можно было бы «нафантазировать», лишь привлекая значительно более широкий запас образов, чем тот который давали оба скрещиваемые и даже другие виды этого рода.
В частности следует отметить значение в ближайшее время гибридизации для пополнения данного; подвергающегося селекции вида сигнальными генами, т. е. такими генами, которые, не имея сами по себе какого-либо важного физиологического значения, но четко менделируя, могут быть использованы для наблюдения за важными, но трудно прослеживаемыми генами, если они окажутся локализованными в той же паре хромосом, вблизи места локализации сигнального гена. В особенности такие гены, как гены окраски, гены различных мелких особенностей оперения или волосяного покрова (хохлы, пучки волос и пр.), не мешающие отбору по основным направлениям, удобны в качестве сигнальных, так как для них прежде всего удается определить их локализацию и дать эскиз плана хромосом (ср. например работу по составлению плана хромосом курицы, для которого использованы именно такие гены). Для животного с большим числом хромосом, — а такими являются все. млекопитающие и птицы — для составления плана хромосом необходимо число генов, в несколько раз превышающее гаплоидное число хромосом, т. е. порядка 1–2 сотен. Поэтому заимствование сигнальных генов от других видов путем гибридизации чрезвычайно важно. Так например путем гибридизации был пополнен список генов мышей (заимствование гена пегости и «танца» от мышей, происходящих от Mus wagneri). Для оценки значения гибридизации, как пути освоения новых генов большой интерес представляет вывод, к которому пришел Баур при сравнительно-генетическом изучении культурных рас и диких видов львиного зева (Baur, 1932). Оказалось, что большинство генов, которыми различаются
культурные расы, не играет роли в образовании
межвидовых различий. Основываясь на несомненных фактах возникновения параллельных (гомологичных). мутаций, как например возникновения многих гомологичных мутаций у Drosophila melanogaster, simulans и др., или на замечательных, почти несомненно гомологичных мутаций у кобылок — Apotettix, Paratettix и Acridium (Nabours, 1933 и др.) и т. п. можно было бы ожидать мутационного возникновения в пределах данного; уже освоенного человеком вида и всех тех генов, которыми от него отличаются другие соседние виды. Если теоретически эта возможность и вероятна, то из указанных многолетних исследований Баура следует, что перспективы этого ожидания вовсе не велики
[8].
Поэтому надо считать, что освоение генов соседних видов путем гибридизации является самым верным, надежным и самым быстрым и едва ли может быть заменено путем отыскания новых мутаций в пределах данного вида.
Приложение к главе II
Выдержка из постановления коллегии НКЗема СССР от 28 сентября 1931 г.
Заслушав доклад проф. Е. Ф. Лискуна и проф. А. С. Серебровского о работе-Всесоюзного института животноводства по гибридизации с.-х. животных, Коллегия НКЗема СССР констатирует, что в то время как в растениеводство гибридизация в работах Мичурина, Бербанка и многих других дала блестящие результаты в смысле быстрого выведения новых полезных форм, животноводство почти lie использовало итого пути.
Учитывая, что в СССР помимо основных социально-экономических условии развертывание работы по гибридизации и смелое выведение новых форм домашних животных значительно облегчаются крупнейшими достижениями советской науки в технике искусственного осеменения и наличием уже некоторого гибридного-материала (яка с крупным рогатым скотом на Алтае и в Средней Азии, гибридов с горбатым скотом в Закавказье и Средней Азии, стада гибридов зубра, бизона и крупного рогатого скота и некоторых других в госзаповеднике «Чанли» (Аскания-Нова) и небольших групп гибридов в Московском зоопарке), Коллегия НКЗема СССР постановляет:
1. Предложить Всесоюзному институту животноводства:
А. Развернуть следующие работы по гибридизации с.-х. животных в масштабах, обеспечивающих быстрое выяснение перспектив и получение хозяйственных результатов:
1) по гибридам яка с местными и культурными породами молочного скота, имея в виду в основном жирномолочность и холодовыносливость гибридов;
2) по гибридам зебу с крупным рогатым скотом в направлении иммунности
к пироплазмозу для юго-восточных районов Союза, учтя опыт США;
3) по гибридам зубро-бизонов, бантенга, гаяла и гаура со скороспелыми породами скота в основном в направлении мясности и кожи;
4) по гибридам зубро-бизонов с яком в направлении шерстности, кожи и мяса;
5) по гибридам овец с крупными дикими видами в направлении мясошерстности;
6) по гибридам свиней в направлении иммунности;
7) по гибридам лошадей, ослов и зебр в отношении тяговой силы и поисков новых направлений;
8) по гибридам гусей и уток в направлении качества мяса, яйца, пуха и пера.
Б. Разработать вопросы:
а) о возможности восстановления плодовитости бесплодных гибридов, в первую очередь мулов, зеброидов и куриных;
б) о возможности болев отдаленных гибридизаций, в первую очередь коров с-буйволами, овец с козами, собак с лисицами и северных оленей с другими видами оленей.
2. В развитие и дополнение постановления Коллегии НКЗема СССР от 28/IX 1931 г. (протокол № 46) о гибридизации с.-х. животных организовать на базе зоопарка и соответствующей части зоотехнической станции Аскании-Нова Институт с.-х. гибридизации и степной акклиматизации животных, включив его в систему Академии с.-х. наук им. Ленина через Всесоюзный институт животноводства, как институт степной зоны СССР, проводящий свою работу под общим плановым и научно-методическим руководством Всесоюзного института животноводства.
3. Поручить Институту животноводства совместно с директоратом Аскании-Нова в декадный срок разработать план работ Института с.-х. гибридизации и степной акклиматизации животных, положив в его основу разрешение следующих проблем:
а) выяснение и установление пределов возможной гибридизации диких и домашних видов животных и создание для этой цели необходимых коллекций;
б) предварительное выяснение возможности и целесообразности хозяйственного использования диких животных и различных гибридов;
в) проведение опытов по межпородной и межвидовой гибридизации в целях отыскания наилучших хозяйственно-полезных комбинаций;
г) акклиматизация животных в условиях степной зоны СССР (изучение влияния внешних факторов среды, отыскание мер борьбы с вредными воздействиями среды, экология паразитов и возбудителей болезней животных и т. д.).
Глава III. Проблема пределов гибридизации
Прежде чем перейти в следующей главе к обсуждению мировых ресурсов для гибридизации важнейших животных, необходимо составить себе хотя бы приблизительное представление о том, в каких пределах систематического родства гибридизация возможна или вероятна. Выяснение этих пределов гибридизации вместе с тем определяет и возможности этой гибридизации, так как очевидно, что чем шире окажутся пределы возможной гибридизации, тем богаче будут ресурсы ее и шире перспективы.
До последнего времени на этот счет имелись нередко преувеличенные представления. Если даже оставить в стороне мифы вроде мифа о Леде и лебеде, то и тогда мы встретим в старинной научной литературе совершенно фантастические сообщения. Интересно, что даже блестящий биолог Реомюр допускал эфективность ухаживаний кролика за голубем. В Москве весьма распространено мнение о возможности гибридизации кролика с кошкой, т. е. представителя отряда грызунов с представителем отряда хищных. Любопытно, что еще в 1883 г. А. П. Богданов в своей «Медицинской зоологии» отмечает существование в Москве этой легенды. В Дагестане распространено мнение о частой гибридизации ослов с коровами и даже, правда всегда из третьих рук, приводят описание этих гибридов
[9], и интересно, что Спалланцани и Боннэ допускали существование подобных гибридов, так называемых юмаров (Iumar).
Говорят и пишут о гибридах овец с козами, собак с лисицами, утки с курицей
[10], курицы с птицей-лирой (Hopkinson, 1922) и т. п.
Где здесь кончается действительность и начинается легенда?
Для того чтобы осветить вопрос о пределах гибридизации, мы должны 1) познакомиться с процессом естественной гибридизации, 2) рассмотреть препятствия, стоящие на ее пути, 3) оценить перспективы преодоления этих препятствий в направлении расширения пределов гибридизации.
Естественная гибридизация
Естественная гибридизация представляет собой вполне закономерное биологическое явление. Там, где встречаются одновременно способные к оплодотворению друг друга виды в известном, колеблющемся от разных причин проценте, вместо нормального спаривания происходит спаривание
гетерогенное, дающее гибридов. Некоторые из таких гибридов либо встречаются столь закономерно, либо столь своеобразно, что первоначально описывались как самостоятельные виды. Так например была описана утка-нырок (Fuligula Homeyery), и лишь после долгих споров выяснилось, что этот редкий («вид» является гибридом двух нырковых уток
[11].
Наиболее известен систематически встречающийся в наших лесах, хотя изредка, тетерев-межняк, признававшийся то за гибрид то за самостоятельный вид: Tetrao hybridus, Т. medius, Т. intermedius, Т. urogalloides, даже просто Т. maculatus (Brehm, 1831).
Позднее специальными опытами было доказано, что межняк действительно является. гибридом между глухарем и тетеревом-косачом. Оба родительские вида в настоящее время относятся уже к разным родам, фигурируя в качестве Tetrao urogallus и Lyrurus tetrix. Действительно, эти птицы сильно отличаются друг от друга и общими размерами, и морфологией (например различной формой хвостов), и привязанностью к несколько различным стациям: глухарь предпочитает более глухие таежные леса, тетерев — более светлые, перелески. Однако на огромном протяжении ареалы распространения обоих видов перекрывают друг друга, а привязанность к различным стациям оказывается не настолько уж сильной, чтобы исключить возможность встреч. И сходство биологии размножения, в частности знаменитые тока, действительно приводит к этим встречам, в результате чего и возникают гибриды.
Тетерев-межняк во всех отношениях является типичным гибридом (рис. 11): по характеру возникновения — возникая хотя и систематически, но редко и лишь в месте совпадения ареалов обоих видов; по морфологическим свойствам — являясь типичной промежуточной формой, совмещая элементы обоих родственных видов; наконец по своему бесплодию, которое повидимому абсолютно несмотря на отдельные, указания на размножение межняков
[12].
Отмечалось также, что межняки оказываются преимущественно самцами. К сожалению этот факт требует проверки, так как самки-межняки меньше отличаются от чистых видов и могли чаще ускользать от наблюдений.
 Рис. 11. Тетерев межняк (фото Ганноверского музея).
Рис. 11. Тетерев межняк (фото Ганноверского музея).
Соприкасаясь в других местах своего ареала с другими видами, тетерев дает новые формы гибридов. Так, на Британских островах добывали гибридов косача с шотландской куропаткой, в других северных районах — гибридов с белой куропаткой (Lagopus lagopus), в Скандинавии и Сев. России — гибридов с рябчиком. Аналогично с белой куропаткой давал гибридов и глухарь.
В параллель с тетеревами приведем еще несколько примеров из различных групп животных. Вдоль Урала находят друг на друга ареалы распространения куницы (Martes martes) и соболя (М. zibellina). В этом районе (рис. 12) систематически возникают гибриды между ними, так называемые кидасы (рис. 13). Эти гибриды здесь столь часты, что например в 1911 г. в районах верхней Печоры, на Унье, Коле и Вишере было добыто 150 кидасов на 58 соболей и 300 куниц.
Аналогичным образом среди млекопитающих, вообще дающих гибридов значительно реже, возникают гибриды зайца беляка с русаком сравнительно редко, но тем не менее настолько закономерно, что гибрид этот получил у охотников специальное название — тумак. Встречается он естественно лишь в тех областях, где водятся оба родительские вида. Из рыб специальное название у рыбаков получил гибрид плотвы и густеры, так называемая вся-рыба.
В Западной Европе налегают друг на друга ареалы распространения двух родственных видов тритонов — Molge cristatus и Molge marmoratus. Здесь же был описан и третий вид, М. blasii, однако после большого обсуждения и проверочных экспериментов выяснилось его гибридное происхождение. Во Франции этот тритон имеет довольно широкий ареал распространения.
 Рис. 12. Карта распространения куницы, соболя и кидаса (по Огневу).
Рис. 12. Карта распространения куницы, соболя и кидаса (по Огневу).
Из насекомых можно привести пример бражников Deilephila euphorbiae и D. vespertilio, гибриды между которыми (D. epilobii) находятся довольно систематично и расшифрованы в качестве гибридов уже около 100 лет назад. В конце XIX в. были находимы и различные дальнейшие поколения этих гибридов, оказавшихся в мужском поле плодовитыми (Mory, 1901).
 Рис. 13. Кидас (фото Огнева).
Рис. 13. Кидас (фото Огнева).
Общее число гибридных комбинаций, возникающих естественно в природе, несомненно очень значительно. То, что попадает в музеи и вообще регистрируется наукой, столь же несомненно является лишь малой долей не только гибридов вообще, — но и гибридов, попадающих в руки человека, так как для этого необходимо, чтобы гибрид попал в руки людей, могущих распознать гибридность или во всяком случае необычность животного и могущих и желающих передать находку работникам науки или в музеи. Отсюда попятно, что сведения о гибридах идут большей частью из стран с наиболее высоким уровнем образования, а целые огромные континенты, вроде Африки, почти «не дали» естественных гибридов, хотя несомненно, что процесс гибридизации идет и там.
Дать правильную картину естественной гибридизации весьма затруднительно. Ввиду сравнительной редкости гибридов очень большое значение приобретает тот путь, которым поступают сведения о них. Наши сведения о разных группах животных в этом отношении совершенно различны. Животные промысловые, добываемые массами и хорошо известные охотникам и любителям (зайцы, тетерева, промысловые рыбы и пр.), находятся естественно в самых благоприятных условиях. И если мы знаем в этих группах много гибридов, то это еще вовсе может не означать какой-либо особой склонности этих видов к гибридизации (см. впрочем Gloger, 1853, 1854), например по сравнению с какими-либо чайками и мелкими воробьиными, а тем более какими-либо мухами или наездниками с их бесчисленными и мало изученными видами, известными лишь редким специалистам.
Однако с необходимыми оговорками можно отметить, например огромное количество известных гибридов птиц по сравнению хотя бы с млекопитающими. Сюшете дал себе труд свести обширную литературу по гибридам птиц, возникших в природе по отделам куриных, утиных, воробьиных и хищных. Им насчитано всего около 200 гибридных комбинаций. Перечисление их здесь было бы излишним. Это количество не идет ни в какое сравнение с гибридами млекопитающих. Среди последних возникших в природе гибридов известно очень мало. Кроме упомянутых гибридов зайцев (можно добавить гибридов с альпийским зайцем) известны, как сказано, кидасы (Огнев, 1929), гибриды соболей и куницы и гибрид хорька и норки. В Ю. Африке был однажды найден целый череп, сочетавший форму черепа антилопы бубала (Bubalus) с рогами типа Domaliscus и определенный Склетером как гибрид между этими соседними родами. Этим может быть и исчерпывается список природных гибридов млекопитающих. Он может быть пополнен случаями естественной гибридизации домашних собак или овец с их дикими родичами.
Было бы конечно ошибкой думать на основании этого, что это различие между птицами и млекопитающими восходит к каким-либо глубоким физиологическим различиям, резко суживающим перспективы гибридизации млекопитающих по сравнению с птицами. Достаточно поставить вопрос об условиях, необходимых для того, чтобы могла пройти гибридизация, чтобы стало ясно, в какой плоскости надо прежде всего искать объяснение этому различию. Для того чтобы в природе мог возникнуть гибрид, необходимо, чтобы в одной местности встречались виды, достаточно близкие для того, чтобы между ними могла произойти гибридизация, чтобы эти виды встречались в одних стациях, чтобы циклы размножения их совпадали, чтобы они могли возбудить друг друга для полового акта, чтобы создались условия, благоприятствующие скрещиванию с самками чужого вида, а не своего; что бы разница в размерах тела или в строении полового аппарата не представляла механических препятствий, чтобы могло произойти оплодотворение, чтобы гибриды оказались способными выжить в обстановке естественной борьбы за существование и т. д. и т. п. Если сравнить например птиц с млекопитающими с этой точки зрения, то уже значительная разница в числе видов ведет к тому, что для млекопитающих вероятность встречи в одной местности двух родственных видов значительно ниже, а всякого рода препятствий гораздо больше. Поэтому для млекопитающих возникновение естественного гибрида должно являться событием совершенно ничтожной вероятности, так как для этого необходимо исключительно счастливое совпадение ряда редких благоприятных условий. В зоологических садах, облегчающих сближение, возникли уже довольно разнообразные гибриды, например двух родов тюленей (Lönnberg, 1929).
 Рис. 14. Гибрид фазана и тетерева (фото Ганноверского музея).
Рис. 14. Гибрид фазана и тетерева (фото Ганноверского музея).
Мы приходим к важному выводу, что
естественная гибридизация лишь в малой степени исчерпывает гибридизационные возможности и что пределы гибридизации естественной гибридизацией далеко не достигнуты. Имея в виду, что все же мы внаем в природе много межродовых гибридов, этот вывод приобретает серьезный интерес. Правильность его легко подтверждается
экспериментом.
Процесс естественной гибридизации однако идет неравномерно, а местами создаются специальные благоприятные условия для него. Такими благоприятными условиями является например распространение в новую область, связанное первое время с трудностями встреч особей одного вида и может быть с нарушением пропорции полов. Ряд подобных, случаев, правда связанный с человеческой деятельностью, зарегистрирован в научной литературе. Так, в недавнее время с охотничьими целями в Швецию был введен заяц Lepus europaeus. и вскоре там весьма часто стали попадаться гибриды с местным L. timidus (Lönnberg, 1905). При введении в Англию глухаря там в первое время стали часто попадаться межняки. Затем этот процесс ослабел очевидно в связи с тем, что глухарь размножился и для него стали легче нормальные встречи с глухарками. Еще более интересный гибридизационный процесс возник в связи с распространением в Западной Европе охотничьего фазана. Дичая здесь, фазан пришел в соприкосновение с тетеревами и глухарями и стал давать сними замечательных гибридов, замечательных прежде всего по своей значительной отдаленности, так как тетерева и фазаны являются не только разными родами, но относятся систематиками к разным подсемействам или даже (см. напр, каталог Британского музея) к разным семействам. Эти редкие гибриды имеются уже в ряде музеев— в Лондоне, Ганновере, Мюнхене и др. (рис. 14). Здесь мы видим, что даже такое легкое вмешательство человека, как содействие фазану в расселении по Европе привело к резкому повышению достигнутого предела гибридизации тетерева со ступени родового различия до ступени различия семейств или по крайней мере подсемейств..
Препятствия к гибридизации
Многочисленные препятствия, стоящие на пути возникновения гибридов и подлежащие преодолению в работе гибридизаторов, могут быть подвергнуты следующей классификации:
1. Препятствия к встрече двух видов:
а) географические — разобщенность ареалов;
б) экологические — приуроченность к разным стациям и биоценозам.
2. Препятствия к спариванию и осеменению двух видов:
а) экологические — несовпадение циклов размножения;
б) рефлексологические — неспособность одного вида вызвать необходимый половой рефлекс со стороны другого;
в) морфологические — несовпадение размеров строения полового аппарата и пр.;
г) физиологические — гибель спермы в чужих родовых путях и пр.
3. Препятствия к оплодотворению:
а) морфологические;
б) физиологические.
4. Нежизнеспособность или малая жизнеспособность эмбриона.
5. Трудные роды или вылупление из яиц.
Природу этих препятствий и возможные пути их. преодоления мы будем подробно обсуждать в главах V, VI, VII. Здесь мы ограничимся лишь самым беглым обзором этой проблемы для того, чтобы иметь возможность хотя бы приближенно ответить на вопрос о том,
каковы перспективы расширения пределов гибридизации при вмешательстве человека по сравнению с гибридизацией естественной.
Географические препятствия не требуют особых пояснений. Лев и тигр могут давать гибридов, но так как ареалы распространения их не совпадают, то в природе эта гибридизация оказывается невозможной. То же самое — белый медведь и бурый, различные неарктические и палеарктические виды, разбросанные на океанических островах в различных бассейнах и т. д.
Но даже при совпадении ареалов распространения может иметь место экологическая разобщенность. Разные виды могут быть приурочены к разным стациям и биоценозам (лесные, луговые виды, обитатели культурных биоценозов и пр.), особенно если виды связаны с определенными растениями. Разные виды могут вести различный образ жизни: дневной, ночной и т. п. Правда, для большинства свободно передвигающихся животных подобного рода препятствия вряд ли могут иметь абсолютный характер (пример тетерева и куропатки), тем не менее они могут играть свою роль, когда гибридизация и без того встречает различные препятствия. Для форм малоподвижных, как молюски, эти препятствия несомненно играют крупную роль не только при гибридизации, но и при обычном размножении, приводя к сильнейшей диференциации на отдельные местные не смешивающиеся расы (яркий пример того мы имеем у молюсков Cerion, изученных Плате).
Более разнообразны препятствия к гетерогенному спариванию и осеменению встретившихся различных видов. Прежде всего они должны находиться в состоянии готовности к половому сближению. Но, как известно, размножение животных связано со сложной физиологической цикличностью, и эта цикличность может далеко не совпадать. Это выражено особенно сильно у млекопитающих. Если у птиц, у амфибий и др. размножение происходит довольно однообразно в немногие весенние месяцы, то у млекопитающих брачные периоды надают в зависимости от длины беременности и пр. на самые разнообразные месяцы. Течка у самок к тому же часто протекает очень быстро, повторяясь 1–2 раза или даже вовсе не повторяясь, и поэтому разница в сезонах спаривапия на 1–2 месяца может явиться сильным препятствием. Между тем такая разница может иметь место и у сравнительно близких видов (например тар и коза, северный олень и марал).
 Рис. 15. Мужские копулятивные органы у мух рода Sarcophaga.
Рис. 15. Мужские копулятивные органы у мух рода Sarcophaga.
Если встретившиеся различные виды находятся в состоянии половой готовности, сближение их должно быть связано с взаимным возбуждением. Здесь для многих видов лежат чрезвычайно серьезные препятствия к гетерогенному спариванию. В старых терминах говорили, что различные виды испытывают «отвращение» друг к другу или по крайней мере «равнодушие». Как увидим в V главе, проблема эта гораздо интереснее антропоморфического толкования в стиле «отвращения». Половой акт у животных является весьма сложным рефлексологическим (инстинктивным) актом, в котором принимают участие разнообразнейшие раздражители — внешний вид, окраска, форма, позы и различные ухаживания, звуки, запахи и т, д. и различные органы чувств, некоторые из которых, как например у насекомых, не всегда нам понятны. В пределах вида самцы и самки путем длительного взаимного приспособления прилажены друг к другу, как ключ к замку и притом взаимно. При встрече же различных видов самка может просто не реагировать на самца и не «поняты) его ухаживаний.
У множества видов (птицы, пчелы и т. п.) обстановка спаривания носит чрезвычайно специфический характер. У других видов, особенно у млекопитающих и птиц, к этому присоединяется еще резкая агрессия по отношению к чужим видам. Попытка пустить к диким баранам домашних овец или коз приводит сплошь и рядом к немедленному убийству самки самцом или наоборот, и требуется предварительное длительное приручение животных друг к другу, чего конечно в природе не происходит.
Наконец часто самцы и самки разных видов, спущенные вместе, совершенно не реагируют друг на друга в половом отношении, и чем менее родственны виды, тем это равнодушие сильнее выражено.
Если встретившиеся виды все же взаимно возбудят друг друга, то попытка к скрещиванию может столкнуться с серьезными морфологическими препятствиями двух типов: либо оба вида могут оказаться слитком различными по величине либо органы совокупления их могут оказаться устроенными не соответствующим друг другу образом.
Каждый многократно мог наблюдать бесплодные попытки к спариванию собак разных размеров. Маленький самец в положении, необходимом для спаривания, не достает до половых органов самки, а маленькая самка при попытках со стороны большого самца либо садится, не выдержав его тяжести, либо выскальзывает из-под самца.
Особенное значение имеет малый рост самца. У кур например крупный петух, раз в восемь превышающий по весу мелкую курицу-бентамку, все же покрывает и оплодотворяет ее, но петуху-бентамке покрыть крупную курицу уже не удается.
 Рис. 16. Гоноподии у Poecilinae (по Т. Регану).
Рис. 16. Гоноподии у Poecilinae (по Т. Регану).
Большое значение может иметь несоответствие в устройстве совокупительных органов, особенно у. насекомых, где их устройство столь разнообразно и характерно, что часто служит серьезным систематическим признаком, различающим виды (рис. 15). Снабженный различными хитиновыми частями, часто острыми, мужской совокупительный аппарат при введении в половой аппарат самки другой) вида нередко наносит смертельную травму или это введение не удается вовсе. У рыб сем. Cyprinodontidae (зубастые карпы) новейшая систематика основывается тоже в значительной степени на разнообразии в устройстве совокупительного органа — анального плавника самца или так наз. гоноподия (рис. 16). У птиц с их чрезвычайно однообразным и простым устройством органов совокупления эти препятствия меньше, что может быть и способствует гибридизации их.
В тех случаях, когда эякуляция вызывается тоже специальными раздражениями, как у млекопитающих, несоответствие половых органов. самца и самки может вероятно привести к тому, что начавшийся половой акт закончится, не вызвав эякуляции.
Наконец, если эякуляция произошла, сперма, попав в половой аппарат самки, может погибнуть или по крайней мере пострадать.
При этом нужно иметь в виду, что у многих животных, для того чтобы добраться до яйца или до того пункта, где происходит нормальное оплодотворение, сперматозоиду приходится проделать очень длинный путь. Например длина яйцевода домашней курицы около
3/
4 м. Сперматозоид про ходит этот длинный путь в течение нескольких часов. Для этого он должен обладать способностью длительно сохраняться в чуждой обстановке. Совершенно ясно, что сколько-нибудь резкое несоответствие в физиологической обстановке, в которую он попадает, лишает сперматозоид возможности пройти этот длинный путь и дождаться момента оплодотворения. Эта категория трудностей, повторяю, почти совершенно не изучена, но можно предполагать, что она должна иметься и служить крупным препятствием при скрещивании более отдаленных форм.
Эти препятствия незаметно переходят в препятствия для самого оплодотворения, тоже весьма многообразные. Их уже довольно много изучали, но в настоящее время мы очень далеки от того, чтобы иметь полную и ясную картину препятствий, которые могут, встретиться в самом акте оплодотворения, т. е. проникновения сперматозоида в яйцо и слияния его ядра с ядром яйца.
Здесь можно наметить следующую категорию препятствий: во-первых, имеют значение химические выделения яйца в окружающую среду, привлекающие к себе сперматозоиды. По этому поводу имеется ряд наблюдений, которые показывают, что если например в. морскую воду, где находится яйцо какого-нибудь вида морских животных (морского ежа), пустить сперматозоидов разных видов — более близких и более отдаленных форм, то эти сперматозоиды различно реагируют на близость этого чужого яйца. Сперматозоиды собственного вида обыкновенно сильно возбуждаются. Очевидно от яйца исходит какое-то выделение, которое заставляет сперматозоид двигаться.
Наоборот, сперматозоиды более отдаленных форм совершенно не реагируют на яйцо. Их темпы движения остаются теми же самыми, совершенно не усиливаются и для них нахождение яйца даже то соседству не оказывает никакого влияния. Правда, мы не можем сказать во всех случаях, какое значение имеет это возбуждение, помогает оно или нет. Но тем не менее здесь открывается известная категория явлений, которая возможно имеет значение для самого акта оплодотворения. Затем имеет значение и устройство поверхности яйца и его оболочек, как мягких, так и твердых (например насекомых) и наконец снабженных микронпле. Сперматозоиды разных видов и групп животных обладают совершенно различным устройством формы своей головки.
У одних сперматозоидов головка устроена более или менее округло. Сперматозоиды других видов имеют очень острый длинный носик (большинство сперматозоидов птиц). Таким образом механизм самого попадания сперматозоида в яйцо должен быть различным. Пфлюгер (Pfltiger) высказал предположение, что те виды животных (яйца) подвергаются легче гибридизации, у которых сперматозоиды имеют тупую головку, а труднее, наоборот, те, где сперматозоиды имеют острую головку. Например острая головка птичьего сперматозоида позволяет предположить, что птичье яйцо имеет такую оболочку, которую сперматозоид должен пробуравить. И поэтому для сперматозоидов птиц и необходим острый носик. И если мы захотим оплодотворить такое яйцо сперматозоидом тупым, то это оплодотворение будет значительно затруднено, и наоборот. Правда, исследователями Лилли, Лебом и другими эта сторона вопроса оставлена в тени. Они считают, что решающими моментами являются физико-химические, а не морфологические. Однако можно усомниться в полноте постановки вопроса этими авторами.
Наряду с этим необходимо указать, что яйца при помощи выделений, о которых мы упоминали, могут оказывать не только возбуждающее, притягивающее влияние на сперматозоид, но и наоборот, эти выделения могут носить характер защитных приспособлений против проникновения чужого сперматозоида. Ряд исследований показал, что яйца многих видов обладают свойством в отношении сперматозоидов других видов аглютинировать их, т. е. склеивать в комочки. Наблюдения показали, что сперматозоиды при попытке гибридизации ими чужих яиц во многих случаях подвергаются аглютинации, т. е. видимо яйца выделяют специальные вещества, которые склеивают сперматозоиды. Наконец помимо аглютативного действия некоторые яйца оказывают токсическое действие, т. е. просто отравляют сперматозоид, отравляют даже тогда, когда сперматозоид проникает в яйцо, где он может погибнуть и в дальнейшем делении не участвовать.
Возможности преодоления препятствии к гибридизации
Чрезвычайно важно выяснить, насколько перечисленные нами многочисленные категории препятствий, стоящие на пути получения гибридов, являются безнадежными, или, наоборот, их природа такова, что преодоление этих препятствий возможно. Это важно выяснить для того, чтобы оценить дальнейшие перспективы гибридизации и то, насколько исчерпаны естественной гибридизацией пределы гибридизации. Детальному обсуждению проблемы преодоления препятствий гибридизации будут посвящены дальнейшие главы, а здесь мы ограничимся лишь кратким анализом положения, чтобы получить в итоге ответ на вопрос о пределах и перспективах гибридизации.
Не требует пояснения то, что
пространственные препятствия устраняются легко простым приводом животных в живом состоянии в одно место. Более трудным является преодоление
временных (сезонных) препятствий, но здесь мы имеем все же ясные перспективы в возможности задерживать или ускорять наступление охоты соответствующим содержанием, кормлением и прямым физиологическим воздействием (см. гл. V). При гибридизации бабочек время лета подгоняют температурными воздействиями на куколок. Также вполне возможно преодоление, по крайней мере во многих случаях, рефлексологических препятствий к скрещиванию, а разрабатываемая все более методика искусственного осеменения снимает и всякие механические препятствия (см. гл V). Если даже в настоящее время мы еще и не в состоянии в каждом данном гибридном сочетании свободно его осуществлять, то во многих случаях мы эти препятствия уже преодолели и с каждым годом вооружаемся все больше и больше.
Значительно большие трудности встречаем мы на пути преодоления препятствий к гетерогенному оплодотворению (см. гл. VI), но экспериментальная зоология показала уже довольно большой опыт в преодолении этих препятствий — правда, преимущественно на различных «лабораторных» видах (морские ежи), стараясь развернуть рамки возможного оплодотворения до крайних возможных пределов скрещивания классов и типов. И поэтому перспективы преодоления препятствий к гетерогенному оплодотворению в рамках, реально интересующих нас (для получения жизнеспособных гибридов), могут быть оценены также как достаточно благоприятные.
Наименее благоприятными оказываются перспективы преодоления нежизнеспособности гибридного эмбриона. Если в некоторых частных случаях в настоящее время мы с этой задачей уже можем справиться (см. гл. VII), то вообще говоря
нежизнеспособность гибридного эмбриона является тем физиологическим пределом, в который упирается стремление расширить рамки гибридизации как в препятствие, уже непреодолимое (кроме некоторых частных случаев).
Однако и здесь (как и в гетерогенном оплодотворении) в нашем распоряжении остается еще возможность
обхода (при наличии некоторых благоприятных условий) этого препятствия методом
гибридизационных мостов. Если вид А не дает жизнеспособного потомства при прямой гибридизации с видом В, то сохраняется возможность найти третий вид С, который мог бы дать жизнеспособное потомство как с А, так и с В с тем, чтобы, получив гибридов АС и ВС, в дальнейших поколениях попытаться получить гибридов ABC.
Но во всяком случае сейчас можно стать на ту точку зрения, что предел гибридизации кладется не препятствиями к осеменению и даже не препятствиями к оплодотворению, но нежизнеспособностью гибридных эмбрионов, работа над преодолением которой в настоящее время может быть даже признается неочередной. Чтобы обосновать это последнее утверждение, посмотрим, как расширяются перспективы гибридизации в связи с преодолением лишь некоторых препятствий к гибридизации — препятствий к осеменению и оплодотворению.
Расширение перспектив гибридизации при вмешательстве человека уже сейчас огромно. Здесь можно ограничиться рассмотрением трех моментов: 1) помощи во встрече видов, 2) помощи в осеменении и 3) помощи в оплодотворении.
При естественной гибридизации возможно скрещивание сравнительно ограниченного количества видов, встречающихся в одной местности и более или менее в одинаковых биоценозах. Если взять например нашу крякву (Anas platyrhynchus), вид чрезвычайно многочисленный, излюбленный объект охоты, добываемый ежегодно в огромных количествах и к тому же легко гибридизирующий, то список ее естественных гибридов окажется все же чрезвычайно ограниченным. Он сводится к следующему:
Anas platyrhynchus × Dafila acuta (шилохвость)
— × Spatula clypeata (широконоска)
— × Chaulelasmus streperus (серая утка)
— × Nettion crecca (чирок-свистунок)
— × Mareca penelope (свиязь)
— × Merganser merganser (крохаль)
Достаточно однако было человеку начать завоз географически удаленных видов в зоосады, в птичьи хозяйства, чтобы этот список значительно расширился. Так, с начала XIX в. стали добываться охотниками стачала в Западной Европе, а затем и в других местах [Закавказье (Radde, 1871)] гибриды домашних уток (или крякв) с южноамериканской мускусной уткой Cairina moscllata, начавшей распространяться по птичьим хозяйствам Европы. А в зоологических садах и парках стали возникать самые разнообразные гибриды кряквы и домашних уток с представителями чуждых фаун и стаций.
В настоящее время зарегистрированы следующие гибриды кряквы, возникшие следовательно в природе, частью при некотором участии хозяйствующего человека, но тоже естественным спариванием.
1. В пределах подсемейства Anatinae — настоящих уток:
— Polionetta paecillorhyncha
— Chaulelasmus streperus
— Mareca penelope
— Mareca sibilatrix
— Mareca americana
— Nettion crecca
— Dafila acuta
— Poecilonetta boliamensis
— Elasmonetta clilorotis
— ? Querquedula querquedula
— Spatula clypaeata
— Alopochen aegyptiacps
— Cairina moschata
— Tadorna tadorna
— Casarca casarca
— Lampronessa sponsa
— ? Aix galericulata
2. С подсемейством Nyrocinae — нырков:
— Netta rufina
— Aythia ferrina
— Fuligula соllaris
— Hymenolaemus malacorhyncha
3. С подсемейством Merginae — крохалей:
— Merganser merganser
4. С подсемейством Ansorinae — гусей:
— Anser domesticus.
В этот список включены также немногие гибриды не с кряквой, а с ближайшим к ней американским видом. Мы видим здесь гибридов не только в пределах родов, но и с соседними и даже с отдаленными подсемействами. Не зарегистрированы гибриды с представителями еще нескольких остальных подсемейств семейства Anatidae: с лебедями, со шпорцевыми гусями, с куриным гусем и с древесными утками. Легко при этом понять, что например с лебедями гибридизация кряквы невозможна по чисто механическим причинам, а с остальными вероятно Частично по рефлексологическим причинам, частично по их сравнительной редкости [шпорцевый гусь (Plectropterus gambiensi) давал гибридов с египетским гусем и с мускусной уткой, участвующими в приведенном списке].
Этот список иллюстрирует 1) огромные гибридизационные возможности кряквы, далеко не исчерпанные данным списком, в котором имеются представители уже половины подсемейств, и 2) этот список гибридов кряквы, превышающий списки гибридов других видов, стоит несомненно в связи не только с большой способностью кряквы давать жизнеспособных гибридов, но не в меньшей степени с ее многочисленностью в природе, в зоопарках и питомниках и с ее свободной размножаемостью в домашних условиях.
Отсюда легко понять, какие огромные гибридизационные возможности открывает дальнейшее устранение механических и рефлексологических. препятствий методом искусственного осеменения. Приведенный список гибридов кряквы (23 вида) составляет примерно около 10 % всего видового состава сем. Anatidae. Наличие гибридов кряквы с представителями 4 подсемейств, обнимающих подавляющее большинство видов семейства, делает весьма вероятным, что искусственное осеменение не только даст гибридов с большинством остальных видов этих подсемейств, но и с остальными подсемействами. И если не во всех случаях окажется возможным непосредственное скрещивание, то уже при помощи мостов все 250 видов семейства окажется возможным вовлечь в взаимное скрещивание.
Так, гуси дают гибридов, с лебедями, а утки — с гусями, шпорцевые гуси — с мускусной уткой и нильским гусем, а эти последние — с кряквой. Остаются невовлеченными в эту связь лишь древесные утки (Dendrocygninae) и куриный гусь (Cereoposes novae-hollandiae), но в настоящее время это вряд ли можно объяснить чем-либо иным помимо отсутствия опытов в этом направлении.
Таким образом преодоление одних лишь препятствий к осеменению, географическое сближение видов и искусственное осеменение способны будут в десятки раз (а для всего семейства в 200–300 раз) расширить перспективы гибридизации.
Это расширение касается двух моментов: 1) числа возможных комбинаций видов и 2) количества возникающих гибридов. Что касается второго момента, то нужно иметь в виду, что естественная гибридизация, а тем более гибридизация в природных условиях представляет собой хотя и закономерное, но чрезвычайно редкое явление. Подведя итоги всем основным данным до конца XIX в., Сюшете приводит па-пример по тем же утиным следующий цифровой материал.
Число зарегистрированных в научной литературе гибридов утиных найденных в природе (экземпляры)
Mareca penelope × Nettion crecca — 4-5
Dafila acuta × Mareca penelope — 4
Anas platyrhynchus × Daphila acuta — ок. 40
A. plat. × Nettion crecca — 10
A. plat. × Chaulelasmus streperus — ок. 12
Dafila acuta × Nettion crecca — 5
A. plat. × A. obscura — 15-20
A. plat. × Mareca penelope — 6
A. plat. × A. americana — 1
Spatula clypeata × D. acuta — 2
D. acuta × Cli. streperus — 1
A. plat. × Spatnla clypaeata — 3-4
Spatula clypaeata × Cairina moschata — 1
Sp. clypaeata × Cli. streperus — 1
A. plat. × Cairina moschata — ок. 20
A. obscura × Cairina moschata — много
Mareca penelope × Ch. streperus — 7
Mareca penelope × Querquedula circia — 1
Casarca casarca × Querq. (? falcata) — 1
Tadorna tadorna × Anas plat. — 1
Querq. circia × Nettion crecca — 1
Anas discoris × Sp. clypaeata — 1
Anas discoris × Acyanoptera — 1
Ch. strepterus × A. americana — 1
Anas poecilorhyncha × A.plat — 1
_____________________________
Итого 150—200
Иными словами, 1–2–3 гибрида в год во всем мире попадали в научную регистрацию.
Искусственное осеменение даже при низком проценте оплодотворения дает возможность резко повысить это количество. Так например за все время птицеводства известны всего 2 естественно возникших гибрида павлина с курицей. Между тем Тинякову в течение 2–3 месяцев работы из 64 искусственно осемененных яиц кур удалось получить двух таких гибридов, а Подвязной и особенно Огороднему — несколько десятков гибридов петух × цесарка против 15 известных в мировой литературе и возникших естественно. Значение преодоления препятствий к оплодотворению уже в указанных рамках гибридизации несомненно велико, хотя сейчас его и трудно иллюстрировать наглядными цифрами. Несомненно однако, что уже при межродовых скрещиваниях оплодотворение получается нелегко. Если например сейчас искусственное осеменение у домашних уток и кур дает около 55 % оплодотворения, то, как мы только что видели, комбинация курицы с павлином дала лишь около 3 %, а оплодотворение кур индюшиной спермой из нескольких сот яиц не дало еще ни одного гибрида или даже развивающегося эмбриона. Хотя не исключена возможность повреждения сперматозоидов индюка в яйцеводе курицы, тем не менее можно думать, что здесь немалую роль играют уже препятствия к оплодотворению. Указанием на это служит следующий опыт. Три кряквы не несущиеся, были осеменены искусственно спермой: первая — селезня, вторая — гусака и третья — петуха. Через 22–25 часов утки были вскрыты, и у всех в самой верхней части яйцевода (воронке) было найдено значительное количество сперматозоидов. Правда, живых сперматозоидов гусака и петуха было найдено меньше, чем сперматозоидов селезня, но тем не менее они имели еще вполне нормальное поступательное движение, т. е. были повидимому годны для оплодотворения.
Имеем ли мы в этих пределах гибридизации какие-либо указания на препятствия Со стороны жизнеспособности гибридов? Пока почти никаких. Наоборот, такие самые отдаленные (в этих пределах) гибриды, как утка и гусь, фазан и тетерев, являются вполне нормальными птицами в смысле их жизнеспособности и общего строения (отсутствие какой-либо асимметрии, уродств и пр.). Многие из гибридов жили по многу лет, например гибриды павлина и цесарки, цесарки и курицы, гуся и утки. Последний гибрид, найденный в деревне вблизи Москвы, вылупился не позднее 1924/25 г., несколько лет потом прожил на биостанции юных натуралистов в Сокольниках, где охотно спаривался с утками и гусынями, а затем жил в 1933 г. в Московском зоопарке и пал в 1933 г. уже в возрасте 8–9 лет. Нормальным был упоминаемый ниже гибрид ибиса с колпицей и др. Если поэтому мыл не имеем оснований и возможности обобщить эту жизнеспособность для всех гибридов в пределах семейств (см. гл. VII), то во всяком случае можем констатировать, что нет оснований сомневаться в том, что например в пределах утиных может быть получено еще множество жизнеспособных гибридов и что даже при скрещивании менаду представителями подсемейств утиных могут получаться вполне жизнеспособные гибриды, и у нас нет серьезных оснований считать, что невозможны жизнеспособные гибриды и при некоторых степенях еще более отдаленных скрещиваний. Во всяком случае
нередко узким местом при получении гибридов является отнюдь не жизнеспособность гибридов, но препятствия к осеменению и оплодотворению. Это не исключает конечно и того, что в некоторых случаях гибридизация даже близких видов может сопровождаться явлениями нежизнеспособности, как в случае скрещивания Drosophila melanogaster × D. simulans — двух чрезвычайно близких, с трудом различаемых видов (см. гл. VII) и др.
Что касается млекопитающих, то наши сведения здесь еще чрезвычайно скудны. Однако и здесь мы знаем ряд межродовых гибридов (Equus × Hippotigris, Bos × Bison, Bos × Poephagus) с жизнеспособностью не только не сниженной, но даже пожалуй повышенной. Между тем уже здесь разница в морфологии и физиологии видов такова, что выдвигает ряд препятствий в области осеменения и родов, но пока мы не имеем никаких серьезных указаний на то, что нынешним пределом гибридизации является нежизнеспособность. Лишь у бабочек мы имеем ряд случаев слабости развития и малую жизнеспособность гусениц многих гибридных комбинаций.
Таким образом при наших современных знаниях в этой области мы можем считать обоснованным утверждение, что
пределы гибридизации, которые определяются фактом нежизнеспособности гибридов у птиц и млекопитающих, и возможно друг их форм, лежат дальше а может быть и значительно дальше тех пределов, которые кладутся препятствиями к оплодотворению и особенно к естественному осеменению.
Иначе дело обстоит у рыб, где препятствия к гетерогенному осеменению в значительной степени сняты внешним оплодотворением (осеменением). Здесь пределы, полагаемые нежизнеспособностью, поэтому значительно ближе к уже достигнутым сейчас. Что же касается насекомых, то здесь почти невозможно дать какие-либо обобщения. Но и здесь препятствия к гетерогенному осеменению (и может быть проникновению сперматозоида в микропиле) в весьма серьезной степени мешают уже ближайшей межвидовой гибридизации.
Таксономические пределы гибридизации
Если мы признаем, что основным и наиболее трудно преодолимым пределом гибридизации является предел, полагаемый нежизнеспособностью эмбрионов, то для оценки перспектив гибридизации и ее ресурсов важно ответить хотя бы приблизительно на вопрос, где же таксономически лежат эти пределы и как далеко таксономически могут отстоять виды, для которых перспективы их гибридизации утрачивают всякую реальность. Вопрос о таксономических пределах гибридизации важен для гибридагогии в том отношении, что систематика животных изучена значительно детальнее, чем возможности скрещиваний. Поэтому если нам- удастся установить некоторые связи между гибридизационными возможностями и таксономическими отношениями, то это будет иметь на долгие годы значение для ориентации в гибридагогической работе и в учете и использовании ресурсов.
Оценивая известные факты гибридизации как животных, так и растений, мы встречаемся с достаточно ясно-выраженной закономерностью, состоящей в том, что чем менее родственны виды, чем дальше отстоят они в системе друг от друга, тем, вообще говоря, труднее проходит гибридизация. Правда, здесь имеется много фактов, осложняющих эту картину, так как в одних случаях формы, очень близкие, дают гибридов с трудом или не дают вовсе, а в других случаях сравнительно легко гибридизируются виды далекие.
С другой стороны, известны случаи резко различного поведения при гибридизации близких видов, из которых одни дают сравнительно легко разнообразных и довольно отдаленных гибридов, другие же почти не способны к гибридизации. Классическим примером этого являются два вида рыб рода Fundulus — F. heteroclitus и F. majalis (см. гл. VI, VII).
Однако эти факты не разрушают общей приблизительной закономерности, параллелизма между трудностью гибридизации и систематическим отдалением. То, что в одних группах животных гибридизация идет легче, чем в других, не должно нас удивлять, как не должна удивлять и различная легкость гибридизации близких видов. Серьезное нарушение обнаруженной закономерности мы имели бы лишь в том случае, если бы данный вид скрещивался легче с более отдаленными видами, чем с более близкими. Однако указать такие примеры довольно трудно. Факты такого рода например, что домашние куры скрещиваются с
краксами, в то время как они же еще далеко не со всеми более родственными видами дали гибридов, — служить такими примерами конечно не могут, так как далеко не всех фазанов и др. пробовали скрещивать с курами, да и все эти гибриды насчитываются буквально единицами, не позволяющими оценивать
степень легкости гибридизации. Но и здесь все же несомненно, что гибридизация кур с куриными же идет очень интенсивно, а с птицами других отрядов уже не идет.
В порядке возрастающей отдаленности, теоретически говоря, мыслим следующий ряд гибридов: 1) межрасовые или внутривидовые, 2) межвидовые или внутриродовые, т. е. гибриды между разными видами одного рода, 3) межродовые или внутрисемейственные, 4) межсемейственные или внутриотрядные, 5) межотрядные или внутриклассовые и 6) межклассовые или внутритиповые, 7) межтиповые. Какие же однако из этих мыслимых степеней фактически получались?
Межвидовых гибридов (внутриродовых) известно очень много среди самых разнообразных групп животных, млекопитающих, птиц, рептилий, амфибий, рыб, насекомых, иглокожих, и в ряде случаев они возникают без всяких затруднений. Точно так же известно большое число и межродовых (внутрисемейственных) гибридов, примером чего могут служить: як × домашний тур, белый медведь × бурый медведь, курица × фазан, глухарь × тетерев, карп × карась и пр. Конечно при оценке межродовой гибридизации возможны и законны сомнения, насколько правильно таксономически возводят различие между этими видами в степень родовых различий, тем более что различные систематики держатся нередко различных взглядов по этому вопросу. Так например як (Poephagus grunniens) и домашний тур (Bos taurus) до недавнего времени относились к одному роду Bos. Точно так же относимые ныне в разные роды белый медведь (Thalassarctos maritimus) — и бурый медведь (Ursus arctos) долго относились к одному роду, хотя еще в 1825 г. Греем было предложено это разделение. Фазан и петух также в свое время относились Линнеем к одному роду Phasianus, а глухарь и тетерев к одному роду Tetrao.
Со времен Линнея мы безусловно имеем изменения в понимании систематиками родовых различий и огромное количество линнеевских родов раздельно на несколько родов каждый. Обсуждать здесь вопрос о том, насколько целесообразно это снижение уровня родовых различий, было бы неуместным, и поэтому, говори о межродовых гибридах, мы должны иметь в виду более новые современные систематики. Однако даже при наличии сомнений в правильности неизбежно условного разделения видов на роды мы имеем немало форм гибридов между видами, отнесение которых к разным родам ни вызывает никаких сомнений. Так, хотя зубра и домашнего тура раньше и относили к одному роду Bos, однако вряд ли можно сомневаться, что различие между туром и зубром несравненно больше, чем между туром и зебу или даже гаялом и бантенгом. С другой стороны, зубр, бизон и несколько вымерших (ископаемых) видов явно выделяются в особую группу видов, позволяющих объединить их в отдельный род Bison.
Также не может быть сомнения в том, что дающие между собой гибридов фазан и петух, цесарка и павлин, цесарка и индюк принадлежат к разным родам, как относил их уже Линней (кроме отнесенных к одному роду, как указано выше, фазана и петуха).
Наоборот, различия здесь столь многочисленны, что возникает потребность подчеркнуть их отнесением этих птиц не только к разным родам, но и к разным подсемействам семейства фазанов. В настоящее время мы уже имеем довольно значительные данные, говорящие за то, что гибридизация между видами, относимыми к разным подсемействам, возможна в ряде случаев. Так как эти случаи, как увидим сейчас, вплотную приближаются к пределам известной гибридизации, то на их. оценке следует остановиться несколько подробнее.
Среди млекопитающих, где по указанным выше причинам известно немного гибридов, наиболее отдаленные гибриды относятся к таким родам, как як, бизон, зебу (Poephagus, Bison, Bos), или лошадь и зебра (родовых гибридов — собака × лиса и кролик × заяц — мы пока оставляем в стороне как оспариваемых). Правда, имеются указания на еще более отдаленную гибридизацию. В 1932 г. появилось интересное сообщение о получении в Ю. Африке в зоосаду гибрида между коровой и самцом антилопы-канны. Канна относится к другому подсемейству, чем корова, к подсем. Tragelaphinae, и подтверждение этого известия очень важно.
Среди птиц гибриды между разными подсемействами довольно многочисленны. Так, известны гибриды лебедя и гуся, утки и гуся утки и крохаля, различных настоящих уток и нырков, нырков и крохалей, гусей настоящих и шпорцевых, — все это сочетание подсемейств. Среди куриных — гибриды фазана и тетерева, курицы и тетерева, курицы и кракса, о котором будем говорить ниже. Упомянутые выше цесарки, индюки и павлины относятся теперь обычно также к разным подсемействам семейства Phasianidae или даже к разным семействам.
Среди, рыб многочисленные гибриды известны между разными подсемействами семейства карповых, например Abramis (подсем. Abramidini) × Rutilus (подсем. Leuciscini), Leucaspius (подсем. Leuciscini) × Rhodeus — (подсем. Rhodeini), Carassius (подсем. Cyprini) x Barbus (подсем. Barbini) и др.
Чрезвычайно интересен межподсемейственный гибрид самца ибиса (Ibis melanppephala) и самки колпицы (Platalea minor), полученный в Берлинском зоологическом саду и оказавшийся к тому же. по краткому сообщению Гейнрота плодовитым. Хотя оба рода относятся к одному семейству ибисовых, но вместе с другими родами образуют в нем два достаточно резкие подсемейства. Приводимые нами (рис. 17) фотографии черепов ибиса и колпицы (других видов) показывают, какой значительной степени достигают различия между этими двумя видами, хотя и сходными в ряде других отношений. Скелетные различия например фазана и тетерева значительно слабее, чем в данном случае.
Полная жизнеспособность и даже плодовитость этого замечательного гибрида естественно ставит вопрос о том, является ли в межподсемейственных гибридах достигнутым предел гибридизации и не возможна ли гибридизация, еще более отдаленная, между разными семействами.
Однако по вопросу о межсемейственных гибридах наши данные, уже очень скудны. В этом случае ответ зависит оттого, где и. как проводить границы между семействами. Таких гибридов, относительно которых все систематики были бы согласны, что их родители относятся к разным семействам, указать очень трудно.
Для млекопитающих гибридов между различными семействами совершенно не известно (если не считать совершенно недостоверных сведений о якобы получавшихся гибридах между коровой и лосем
[13]). Среди птиц о таких гибридах говорить уже возможно. Так, если согласиться с системой птиц, принятой в каталоге птиц Британского музея (Ogilvie-Grant, Catalogue of the Game Birds in the collection of, the British Museum, 1893), то гибриды между фазаном и тетеревом;, курицей и тетеревом будут уже. межсемейственными гибридами (Phsianidae × Tetraonidae), а гибриды кур с краксами (гокко и Пенелопой) — не только межсемейственными, но даже межподотрядными. По системе куриных Шарпа (Sharpe, 1899), возводящего фазанов в ранг подотряда с подсемействами тетеревиных, фазановых, цесарок и индеек межсемейственными гибридами будут также и гибриды фазанов, кур и павлинов с цесарками и цесарок с индейками, а гибриды кур и фазанов с краксами — межподотрядными гибридами. Наоборот, по системе птиц, привитой Мензбиром («Птицы»), тетерева, фазаны, цесарки и индейки составляют подсемейства фазановых и лишь краксы являются особым семейством. Что касается последних, то хотя прежние систематики и присоединяли их к фазанам в качестве подсемейства, но в настоящее время согласно всем руководящим систематикам гибридов
кур с краксами можно считать межсемейственными гибридами.
 Рис. 17. Черепа ибиса и колпицы сверху и сбоку (фото Аскании-Нова).
Рис. 17. Черепа ибиса и колпицы сверху и сбоку (фото Аскании-Нова).
Совсем недавно обнаружились интересные гибридизационные явления (и притом в природе) в группе райских птиц. Производя их ревизию, Штреземан (Strescmann, 1930, 1931) и Ротшильд (Rotschild, 1930, 1931) привели к заключению, — надо сказать достаточно обоснованному, — что примерно из 6 десятков видов райских птиц 18 видов и даже 8 родов являются лишь гибридами. Этим объясняется как их промежуточный характер, так и редкость нахождения (некоторые — уникумы). При этом интересно, что в образовании гибридов принимали участие далекие роды и подсемейства, а по системе Шарпа, принятой в Британском музее, — даже роды двух различных семейств— Paradisaeidae и Ptilonorhynchidae. Схема (рис. 18) показывает- эти гибридизационные связи с возникающими в результате гибридами. В атласе Гульда, посвященном птицам Новой Гвинеи (1, табл. XXI; этот атлас имеется в библиотеке Зоол. муз. Академии наук), изображен под видовым именем. Diphillodes gulielmi III один из наиболее убедительных таких гибридов, родительские виды которого также изображены на соседних таблицах (Diph. magnificus и Cicinnurus regius). В журналах Ibis (1890, 1911), Journ f. Ornithol. (1902) и в Novit. Zoologicae (1903) можно найти цветные изображения большинства этих гибридов, и стоит потратить время, чтобы познакомиться с этими замечательными птицами.
 Рис. 18. Схема гибридизации райских птиц.
Подчеркнуто сплошной линией сем. Ptilonorhyncliidae; прерывистой — сем. Paradiseidae; подсем. Paraniseinae; точками — сем. Paradiseidae подсем. Epimachinae.
Рис. 18. Схема гибридизации райских птиц.
Подчеркнуто сплошной линией сем. Ptilonorhyncliidae; прерывистой — сем. Paradiseidae; подсем. Paraniseinae; точками — сем. Paradiseidae подсем. Epimachinae.
Так как все эти гибриды найдены в природе и никто еще не пробовал вести с райскими птицами гибридизационную работу, то надо думать, что и здесь еще не достигнуты пределы возможной гибридизации. Нужно впрочем иметь в виду, что разделение Шарпом райских птиц на 2 семейства далеко не признано и представляет такой же увлечение, как и разделение им фазановых на 4 семейства (см. Мензбир, «Птицы»).
Других столь же отдаленных гибридов ни среди птиц ни среди рыб, насекомых и пр. мы больше не знаем. Здесь однако необходимо оговориться. Мы считаем целесообразным, говоря, что такие-то гибриды
получаются, относить это выражение только к тем случаям, когда гибриды оказываются более или менее, жизнеспособными. Ниже мы увидим, что более или менее удачные попытки гибридизации делались и далеко за пределами семейств. Но конечно там, где в результате гетерогенного осеменения получаются лишь начальные стадии эмбриогенеза или уродливые эмбрионы, говорить, что «получен гибриде, можно лишь в совершенно ином смысле, чем например, когда мы говорим, что получен гибрид лошади и осла, фазана и кракса. В таких случаях так и следует говорить: получено оплодотворение, получен нежизнеспособный эмбрион и т. д., а о том, что получен гибрид, говорить лишь тогда, когда гибрид все же более или. менее способен к развитию и сформированию в развитой организм, хотя бы и слабый, выживающий в виде отдельных экземпляров, как например в случае гибридизации некоторых бабочек, когда гибридные гусеницы массами гибнут.
Что касается гибридизации; более отдаленной, чем межсемейственная, то попыток добиться ее было сделано немало. В ряде случаев опыты приводили к оплодотворению, к дроблению и эмбриогенезу. Однако эмбриогенез в таких случаях всегда был ненормальным и приводил к отмиранию (например гибридизация жаб с лягушками). Начало дробления получалось при гибридизации, гораздо более отдаленной, чем семейственная: удавалось оплодотворение при межотрядном (лошадь × кролик), межклассовом (морской еж × морская лилия) и даже межтипном (морской еж × молюск) оплодотворении.
Какова природа этих гибридов? Общей чертой этих гибридов является их нежизнеспособность. Хотя развитие эмбриона и начинается, но оно не протекает нормальным порядком. Эти отдаленные скрещивания сопровождаются очень
большими нарушениями, и ни одного гибрида, который бы дожил не только до взрослого состояния, но даже до более или менее развитого эмбриона, при таком отдаленном скрещивании получено не было.
Правда, неожиданные результаты были получены в самых отдаленных скрещиваниях, когда например скрещивали морских ежей с морскими звездами (разные классы), с молюсками и червями (разные типы). Здесь гибридизация как будто удавалась и даже получалось развитие эмбрионов совсем нормальное или почти нормальное; однако было выяснено, что в этих случаях настоящей гибридизации не происходит. Всегда, когда в результате отдаленной гибридизации получаются нормальные зародыши, они в точности повторяют свойство матерей. При оплодотворении морских ежей спермой молюсков или морских звезд получаются личинки морских ежей, и никаких отклонений в сторону молюсков не обнаруживается. Здесь отсутствует настоящая гибридизация, потому что оплодотворение дает лишь толчок к развитию, но сперматозоид не принимает участия в дальнейшем развитии эмбрионов (ср. рис. 39 и 41). Сперматозоды подвергаются отмиранию, и все дальнейшее развитие идет исключительно за счет материнского ядра. Эти явления очевидно приходится отнести к категории «искусственного партеногенеза»
[14].
Некоторые резкие изменения среды вызывают со стороны яйца реакцию, приводящую к развитию эмбриона без оплодотворения. Возможность такого объяснения далекой гибридизации видна из интересных опытов П. и Г. Гертвигов. Гибридизируя жаб с лягушками, они получили следующие парадоксальные результаты: при оплодотворении икры лягушки нормальной спермой жабы развитие далеко не идет, потому что получаются уродливые эмбрионы (рис. 19). Если же сперматозоиды обработать радием, то развитие идет нормально. Сперматозоиды, обработанные радием, сохраняют способность проникать в яйцо, но утрачивают способность участия в дальнейшем развитии. Таким образом и здесь получается искусственный партеногенез, и этот поврежденный радием сперматозоид служит только толчком для возбуждения развития. Ничего жабьего этот сперматозоид в эмбрион не вносит, потому что ядро его отмирает.
Подводя еще раз итоги, мы можем сказать, что таксономические границы возможного получения жизнеспособных гибридов пока идут (в самой общей форме!) по границам семейств. Гибридизация представителей разных семейств уже не дает жизнеспособных гибридов, что и кладет предел гибридизации, которая при настоящем уровне науки имеет реальные перспективы при работе лишь в пределах семейств.
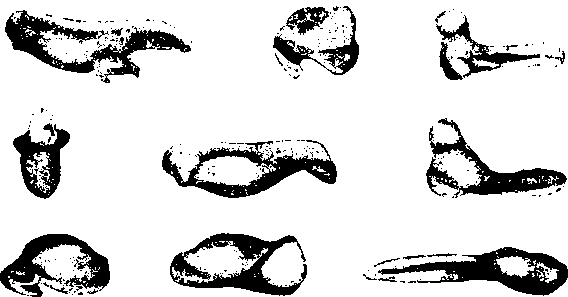 Рис, 19. Уродливые гибриды Bufo × Rana в возрасте 12 дней (по Г. Гертвигу).
Рис, 19. Уродливые гибриды Bufo × Rana в возрасте 12 дней (по Г. Гертвигу).
Устанавливая приблизительные границы возможности гибридизации по границам семейств, мы должны конечно помнить всю относительность понятия семейства, подсемейства, подотряда и т. д. В разных группах животных и растений эти понятия не совпадают, и различие, которое в одних классах принимается за семейственное, в других может оказаться принятым за подсемейственное или, наоборот, за подотрядное. Наши выводы о пределах гибридизации связаны с анализом таких семейств, как фазановые (Phasianidae), утиные (Anatidae), как семейства быков (Bovidae). Для других классов мы имеем однако недостаточно материала, чтобы сказать, насколько и там пределы получения жизнеспособных гибридов совпадают приблизительно с границами семейств. Однако например у насекомых за последние десятилетия объемы семейств подвергались столь резким изменениям, что в иных случаях быть может окажутся возможными и межсемейственные скрещивания.
Если мы поставим как последнюю границу скрещивание между подсемействами, то спрашивается, не слишком ли это сужает наши перспективы как гибридизаторов? Нужно определенно сказать, что это оставляет для нас чрезвычайно широкие пределы работы< Возьмем например семейство Bovidae. Это семейство охватывает и коров, и овец, и всех антилоп, т. е. около двух сотен разных видов, чрезвычайно сильно отличающихся друг от друга, имеющих массу оригинальных свойств. Если обратиться к птицам, то семейство фазановых насчитывает свыше сотни видов, к числу которых относятся и куры, и фазаны, и куропатки, и тетерева, и рябчики, и целый ряд других форм, чрезвычайно, разнообразных по биологическим свойствам и по их экономическому значению. И здесь в пределах семейства имеются огромные перспективы для гибридизационной работы.
Если взять семейство утиных, то здесь мы имеем также почти 250 видов, многие из которых при скрещивании заведомо дают гибридов. Поэтому, даже полагая, что пределы гибридизации лежат в рамках семейства, мы можем иметь огромный материал для гибридизационных работ.
Для того чтобы сделать картину перспектив гибридизационной работы еще более ясной, необходимо познакомиться с мировыми ресурсами гибридизации.
Глава IV. Мировые ресурсы гибридизации
Ресурсы видов
Понятие о мировых ресурсах и их классификация
Под термином мировые ресурсы гибридизации следует понимать совокупность различных форм ныне живущих домашних и диких животных, которые в тех или иных целях используются или могут быть использованы для целей гибридизации и знание которых, возможно более исчерпывающее по охвату и глубине, обязательно для правильной постановки всей работы.
В широком смысле такими ресурсами оказывается весь современный животный мир, рассматриваемый с данной специфической точки зрения. Ввиду необъятности этих ресурсов необходимо подвергнуть их более детальной классификации, в процессе которой понятие о ресурсах приобретет большую ясность и конкретность.
Прежде всего введем понятие о
центрах и
концентрах ресурсов. Под центром мы будем разуметь тот вид животного, который почему-либо является средоточием внимания гибридизатора. Таким центром может явиться прежде всего какой-либо вид домашних животных, над усовершенствованием и реконструкцией которого работает гибридизатор. В другом случае — вид животного, который мы предполагаем акклиматизировать, ввести в нашу фауну, используя для этих целей прием гибридизации, и т. п. Поскольку на данном виде сосредоточиваются наше внимание и работа, он и становится центром, и у нас возникает потребность выделить концентр мировых ресурсов, которые могут быть использованы в работе с данным центральным видом. Поэтому включение того или иного вида в круг ресурсов происходит не абстрактно, а конкретно в форме включения его в тот или иной концентр — ресурсоконцентр. Какая-нибудь дикая бабочка — шелкопряд — не будет ресурсом относительно коровы, но может оказаться в числе ресурсов для шелкопряда, и обратно. Это понятно, и это сразу выделяет из общей массы животного мира уже гораздо меньшую его часть.
Итак, вопрос о гибридизационных ресурсах должен ставиться конкретно относительно каждого центрального вида и уже затем в более общей форме, например гибридоресурсы пушных зверей, гибридоресурсы мясных животных, декоративные ресурсы и пр.
Гиркон и его состав
Пользуясь ставшим привычным приемом сокращения, введем термин
гиркон для обозначения гибридов ресурсоконцентра. Данный вид будет центром его гиркона, и различные виды рода и пр. должны будут расположиться в некоторых отношениях к центру. В самом широком объеме для каждого вида весь остальной животный мир мог бы считаться гирконом, учитывая например опыты ультрагибридизации, при которых удавалось получить начальные стадии развития гибридов даже разных классов и подтипов. Такое расширение рамок гиркона не будет иметь практического значения, и под гирконом мы будем разуметь лишь совокупность тех видов, от которых мы либо можем получать более или менее жизнеспособных гибридов либо можно иметь надежду на получение их в будущем по мере успехов гибридизации.
Но если даже после сделанного ограничения мы зададимся вопросом о составе гиркона данного вида, то увидим, что и здесь необходима дальнейшая классификация. В самом деле, какие виды животных надо включить например в гиркон овцы? Мы имеем, во-первых, группу видов рода Ovis, в том числе родоначальников домашних овец; имеем виды рода Capra, козлов и некоторые другие виды соседних родов, входящих в подсемейство Ovicaprinae, относительно которых либо имеются отрывочные гибридологические данные либо их не имеется вовсе. Далее идут овцебыки, быки и антилопы, входящие в качестве других подсемейств в семейство Bovidae, вопрос о гибридизации которых либо вовсе не ставился либо ставился в порядке научного или хозяйственного фантазирования. Затем идут другие семейства парнокопытных, копытных и другие отряды млекопитающих, затем классы типа позвоночных и наконец различные другие типы животного царства.
Какие из них можно и должно включать в гиркон вида овцы?. Вопрос о проведении рациональных границ гиркона далеко не прост. Прежде всего необходимо подчеркнуть, что включение в гиркон есть динамический процесс, и для каждого данного уровня науки и техники содержание гирконов будет иным. В предыдущей главе мы подробно обсуждали препятствия, стоящие на пути гибридизации, и возможность преодоления этих препятствий, — Так как при каждом успехе гибридагогии будут преодолеваться те или другие препятствия, то в круг ресурсов будут включаться все новые и новые виды. Прекрасным примером этого является разработка искусственного осеменения, которое сразу включает в ресурсы гибридагогии все те формы, с. которыми ранее скрещивание было невозможным по рефлексологическим и механическим причинам.
С другой стороны, каких-либо пределов гибридизации мы указать сейчас не можем, так как нам еще не известны те причины, которые должны сделать такую гибридизацию абсолютно невозможной, а препятствия непреодолимыми, и поэтому сейчас эти пределы мы вынуждены определять эмпирически, пользуясь постепенно совершенствуемой техникой. В предыдущей главе мы пришли к выводу о том, что рационально — в настоящее время видеть такие пределы приблизительно в пределах семейств.
Все видовые ресурсы каждого гиркона на. каждом данном уровне развития техники гибридизации можно делить на отделы или «круги»: α (альфа) «плодовитый круг» — виды, дающие прямо или черев посредство мостов плодовитых гибридов с центральным видом; β (бета) «бесплодный круг» — виды, дающие бесплодных гибридов; — γ (гамма) «сопредельный круг» — виды соседних подсемейств или семейств, с которыми пока гибридизация не удается. Изучение этого сопредельного круга также важно для гибридизатора, освещая перспективы его работы и помогая выбрать наиболее подходящие объекты для опытов по расширению возможностей гибридизации.
Все остальные более отдаленные виды можно назвать «внешним кругом», интересным только для ультрагибридизации и связанных с ней теоретических проблем.
Изучение каждого гиркона должно вестись всеми доступными методами. В первую очередь, как сказано, должен применяться прямой эксперимент. Однако, имея в виду, что добывание для экспериментальной работы диких видов, особенно видов редких
[15], сопряжено часто с большими трудностями и затратами, важно использовать все имеющиеся данные для предварительного отнесения вида (предположительно) к тому или иному кругу. Здесь должны быть использованы под новым углом зрения данные систематики, сравнительной анатомии и палеонтологии, а если возможно, то и сравнительной; эмбриологии и физиологии, выяснена степень родства, наличие особенностей в половом аппарате и других важных органах, особенности эмбриогенеза с точки зрения того, нет ли здесь указаний на возможность серьезного нарушения развития гибридного зародыша и т. п. По подобной программе и должна быть проведена первая часть, изучения гирконов — изучение ресурсов видов.
Задачей изучения ресурсов видов, входящих в гиркон, должно явиться:
1. Определение степени систематической удаленности крайних видов гиркона для определения внешних границ гиркона, в пределах которых гибридизация может рассчитывать на удачу.
2. Изучение родственных взаимоотношений между видами, родами и подсемействами и расположение их относительно центра гиркона для того, чтобы гибридизатор мог планировать очередность, своих работ, выбирать мосты к более отдаленным членам гиркона.
3. Определение степени бесплодия, жизнеспособности и природы препятствий к гибридизации между центром гиркона и его членами-и членов друг с другом, т. е. перевод сравнительно-анатомического-масштаба взаимных расстояний между видами на масштаб гибридизационный, или иначе — проведение границ между кругами, перечисленными выше. В первую очередь отыскание всех комбинаций, дающих плодовитых гибридов.
Проблема «формы» гиркона
Ориентируя гирконные виды относительно центра, как более близкие и более далекие, и беря, хотя бы и крайне приблизительно, как это делается при построении «родословных деревьев», масштаб, отражающий это расстояние, мы можем поставить себе задачу некоторого графического изображения строения гиркона, схематического взаимного расположения гирконных видов как относительно центра, так и относительно друг друга. Такая графика гиркона была бы очень полезной для гибридизатора при разработке планов его работы. Выбрав термин «круг», «концентр», мы тем самым как бы приняли концентрическую схему, строения гиркона (рис. 20).
Надо однако мотивировать ту степень правомочности, с которой мы это приняли.
 Рис. 20. Схема строения гиркона.
Рис. 20. Схема строения гиркона.
Обычно родословные дерева рассматриваются как бы сбоку, когда они именно и похожи на дерево. Для целей построения схемы гиркона не представляет трудности так перечертить родословное древо, чтобы его ветви были ориентированы относительно интересующего нас центра и преемственный ряд видов от корня древа до «центра» представлял бы собой прямую линию, ось. Эго и будет тауроцентрическим, овицентрическим и т. д. изображением родословного древа, на котором сейчас соответствующие ветви изображаются в виде «боковых ветвей» и прямой ствол идет более или менее прямо к человеку.
Для целей гибридизации является заманчивым взглянуть на родословное древо гиркона не сбоку, а сверху, чтобы уяснить себе взаимное расположение концов его ветвей. Впервые чрезвычайно интересную попытку такого рода сделал Фюрбрингер, давший три горизонтальных среза родословного древа птиц (рис. 21). Его систематика ныне живущих видов таким образом оказалась схематически построенной на плоскости и даже в Пространстве.
 Рис. 21. Схема классификации куриных Фюрбрингера; числа в кружках — число видов.
Рис. 21. Схема классификации куриных Фюрбрингера; числа в кружках — число видов.
В новейшее время, изучая фазанов, Биби (Beeby 1918–1922) пришел также к заключению о невозможности какого-либо линейного расположения современных видов этого семейства и дал его в циклическом изображении (рис. 22). Правда, его схема по отношению к большинству видов остается все же линейной с той лишь разницей, что, удаляясь от петуха в одном направлении, мы через кольцевой путь снова приходим к петуху, но уже с другой стороны, через другие виды.
 Рис. 22. Схема классификации фазанов Биби.
Рис. 22. Схема классификации фазанов Биби.
Совершенно ясно конечно, что реальные взаимоотношения между видами не могут быть изображены ни прямой, ни кругом, ни кубом, ни какой-либо другой геометрической формой. Однако проблема формы гиркона вовсе не является надуманной и связана с вопросом, для гибридизации чрезвычайно важным, о том,
может ли быть несколько видов или групп видов (родов, семейств и пр.),
более или менее равно удаленных от центра гиркона, или такая группа (вид) лишь одна и именно на ней должно быть сосредоточено в первую очередь внимание гибридизатора.
Обозначая скорость эволюционных изменений разных ветвей родословного древа через v
1, v
2, v
3, а расстояние в поколениях через n
1, n
2, n
3 …, будем иметь в качестве мерила «расстояния» между видами величины n
1v
1, n
2v
2, n
3v
3 и т. д. Разные свойства видов могут эволюировать конечно с разными скоростями, но для нас сейчас это особого значения не имеет, так как v
1, v
2, v
3 могут обозначать либо средние скорости многих признаков либо скорости эволюции одного или нескольких решающих признаков.
При таком измерении расстояний гирконных видов от центра мы можем допустить очевидно уже неопределенно большое число видов, равно или приблизительно равно удаленных от центра. Несомненно, что таково именно положение и в объективной природе. Это позволяет нам схематически изображать строение гиркона в виде концентров; и искать равно удаленные от центра виды не в одной какой-нибудь, родственной, группе видов, но в различных, друг от друга далеко отстоящих. Эти равно удаленные от центра, но далеко отстоящие друг-от друга гирконные виды должны будут наноситься на схеме гиркона в различных местах окружности. Схема гиркона в таком изображении будет наиболее близкой к схеме, данной Фюрбрингером, — рассмотрение родословного древа «сверху».
 Рис. 23. Схема взаимных отношений четырех родов куриных (по Полю).
Цесарка стоит на вершине косой трехгранной пирамиды.
Рис. 23. Схема взаимных отношений четырех родов куриных (по Полю).
Цесарка стоит на вершине косой трехгранной пирамиды.
Однако нужно помнить, что эта схема дефектна в том отношении, что, позволяя расположить гирконные виды относительно центра, она не позволяет все же правильно расположить их друг относительно друга. Происходит это от того, что направлений эволюции: много. Точно на плоскостной схеме могут быть расположены лишь, три вида, образуя треугольник. Четыре вида образуют тетраэдр (см. схему Поля для курицы, фазана, цесарки и павлина, рис. 23). Расположение более чем четырех видов требует уже многомерной схемы. Принимаемая же нами плоскостная схема гиркона является; не более, как проекцией этой многомерной схемы на. плоскость. Так она и должна пониматься.
Ресурсы признаков
Феноресурсы
Изучение ресурсов, признаков или фенов, наследуемых различий и свойств разных видов — членов гиркона — должно наполнить конкретным содержанием все наше знание гиркона. Именно ресурсы признаков, накопленных эволюционным процессом в данном гирконе, представляют ближайший интерес для гибридагога. Видовые ресурсы интересны для пас лишь как средство к использованию феноресурсов, а геноресурсы, хотя и представляют еще более глубокий интерес, но теснейшим образом связаны с феноресурсами, а частично познаются уже через феноресурсы. Если мы находим у яка своеобразную оброслость, находим оригинальную оброслость у бизона и т. д., то тем самым мы уже получаем предварительное представление и о геноресурсах, и дальнейшее изучение может лишь (правда, чрезвычайно сильно) дополнить наши знания о возможности получения еще новых признаков путем перекомбинирования генов.
В настоящее время приходится констатировать чрезвычайно ничтожную изученность феноресурсов даже в главнейших гирконах. Все, что мы знаем о диких видах — членах этих гирконов, сводится к: 1) более или менее детальному внешнему описанию, обслуживающему однако лишь запросы систематики; 2) сравнительно-анатомическим данным, чрезвычайно отрывочным и опять-таки обслуживающим лишь запросы систематики и эволюционного учения; 3) достаточно элементарным экологическим сведениям — зоогеографическим и фенологическим и 4) лишь для немногих животных — к охотничье-промысловой оценке, имеющей некоторую полноту уже лишь для совсем единичных видов.
Особенно ощутительным пробелом является, можно сказать, не рискуя ошибиться, полное отсутствие сравнительно-физиологических данных. Что например мы можем сказать об особенностях обмена веществ у различных видов — членов гиркона коровы? А между тем несомненно, что у яка, бантенга, у различных антилоп, живущих в самых различных условиях и климатах, должны быть здесь крупные различия, большое разнообразие
[16], которое могло бы выдвинуть боевые задания по освоению их в интересах соцстроительства. А пока мы вынуждены оценивать интерес того или иного гибридного скрещивания лишь по самым элементарным внешним различиям между видами — по величине, по общему характеру оброслости или оперения и т. п.
Изучение феноресурсов должно выразиться прежде всего в составлении полного списка отличий каждого вида от других по всем категориям признаков экстерьера, интерьера — анатомии и гистологии, физиологии, рефлексологии, биохимии как взрослых, так и эмбриональных стадий, определении разнообразных параметров для каждого признака.
Выяснение крайних вариантов должно составить следующую стадию обработки собранных параметров. Надо иметь в виду, что задача выяснить, у какого вида мы имеем крайнее развитие какого-либо признака, достаточно сложна и требует изучения закономерностей коррелятивной изменчивости. Пусть нас интересует например вопрос, у какого вида в гирконе коровы самая маленькая голова, в расчете на то, чтобы использовать соответствующую часть генотипа этого наиболее мелкоголового вида. Если мы просто измерим длину головы, всего тела, то конечно самая маленькая голова окажется у самой маленькой антилопы. Но этот ответ очевидно не тот, какой нам нужен. Если мы возьмем относительный нес головы, то самым маленьким он, наоборот, окажется у одного из самых крупных видов, может быть у бизона, являющегося тем не менее несомненно самым головастым во всем гирконе. Из физиологии роста и из сопоставления всех млекопитающих видно, что при всяком увеличении размеров тела голова увеличивается гораздо слабее других частей тела. Следовательно и такая примитивная поправка на величину тела для нас не годится, так как нам важно найти такую «маленькую голову», которая позволила бы надеяться выделить именно особую часть генотипа, характерную для данного вида, с тем чтобы перенесение его на корову дало уменьшение ее головы, не меняя общих размеров тела. Очевидно, что для того, чтобы эту задачу разрешить, нужно найти закон регрессии, связывающий величину головы с величиной тела, и судить о величине головы того или иного вида, основываясь уже на атом законе.
Следующий аналогичный пример покажет нам ход решения этой типовой задачи. В гирконе утки форма яиц весьма разнообразна: есть яйца очень круглые и очень удлиненные. Какие же виды имеют самые круглые яйца и какие — самые удлиненные? Если взять отношение ширины яйца к его длине у разных видов, то мы получим примерно следующий ряд:
0,750 — Lampronessa sponsa — 23 (номер на графике)
0,750 — Mergus allbolus — 2
0,745 — Erismatura leucocephala — 3
0,732 — Anas platyrhynchus — 9
0,728 — Tadorna tadorna — 15
0,725 — Alopochon aegyptiacus — 16
0,716 — Fuligulafuligula — 6
0,710 — Mareca penelope — 10
0,702 — Histrionicus histrionicus — 8
0,700 — Clangula clangula — 17
0,695 — Querquedula querquedula — 11
0,683 — Anser albilrons — 18
0,670 — Brenta canadensis — 22
0,653 — Brenta bernicla — 21
0,665 — Ansererythropus — 19
0,638 — Chenopsis atratus — 25
0,691 — Cygnus cygnus — 24
Самые круглые яйца оказываются у каролинской утки — 0,750, а самые длинные у лебедя. Вместе с тем самые круглые яйца оказываются у самого мелкого вида, самые удлиненные — у самого крупного. Что это не случайность, видно и по другим видам: вверху списка стоят все мелкие виды, внизу — крупные. Очевидно в этой группе птиц существует связь между величиной тела и формой яиц, и длина яйца растет быстрее, чем ширина.
Следующая диаграмма (рис. 24) показывает графически эту связь для утиных между длиной яйца и его шириной. На графике (на рассматриваемом участке) мы отчетливо видим простую линейную связь этих величин от самых мелких яиц Lampronessa до самых крупных Cygnus. Эта прямая регрессия выражается уравнением
y = 0,542
x + 10,3.
 Рис. 24. Корреляция длины и ширины яиц у утиных.
Слева контуры широкого, среднего и узкого яйца. Нижний ряд цифр — величина индекса, отвечающая линии регрессии. Точки 22 и 17 следует сдвинуть на одну клетку вниз.
Рис. 24. Корреляция длины и ширины яиц у утиных.
Слева контуры широкого, среднего и узкого яйца. Нижний ряд цифр — величина индекса, отвечающая линии регрессии. Точки 22 и 17 следует сдвинуть на одну клетку вниз.
На графике вместе с тем видно, что в то время как большинство видов расположено очень близко к линии регрессии, некоторые довольно сильно уклоняются от нее, доходя до 10 % в ту или иную сторону. И здесь мы обнаруживаем, что в минус-сторону почти на 10 % отклоняются Oedemia nigra и Querquedula querquedula, а в плюс-сторону (более круглые яйца) Erismatura leucocephala
[17].
Таким методом мы устанавливаем, что у савки (Erismatura) яйца наиболее отличаются в сторону большей ширины от имеющей место в группе утиных закономерной регрессии ширины яйца по длине яйца/ По сравнению с другими видами, имеющими яйца такого же размера, савка является исключительно круглояичной, что несомненно отражает особенности ее генотипа. При сравнении же савки с еще более круглояичной каролинской уткой возникает очень важный для гибридагогии вопрос о природе этих различий. Для того чтобы ответить на этот вопрос, нужно вскрыть природу той корреляции, которая связывает форму яйца с величиной птицы. Необходимо различать два типа корреляции — корреляцию физиологическую, или онтогенетическую, и корреляцию историческую, или, вернее, филогенетическую. Обе они имеют для гибридагогии совершенно различный интерес и значение, будучи не отличимы по форме. Физиологической корреляцией мы называем ту, которая возникает на базе процессов индивидуального развития. Когда например животное растет, то у него увеличиваются все части тела, однако с разными коефициентами. Трубчатые кости увеличиваются быстрее, плоские кости, в том числе кости черепа, примерно в квадратный корень медленнее и т. д. На этой базе путем ослабления или усиления общего роста можно получить изменение пропорции тела, получить более головастых или менее головастых животных и т. д. Совершенно иначе связываются признаки филогенетической корреляцией, создаваемой естественным отбором, примером чего служит относительное утолщение и укрепление скелета конечностей при увеличении размеров вида. Известно, что у крупных животных скелет относительно во много раз массивнее, чем у мелких, что легко объясняется учением о сопротивлении материалов: пропорциональное увеличение всех частей скелета мыши до размеров быка привело, бы к тому, что такое животное разломилось бы на части под действием собственной тяжести. Поэтому в процессе эволюции увеличение размеров должно сопровождаться параллельным отбором генов, вызывающих добавочное утолщение скелета (хотя вероятно к этому примешивается и физиологическая корреляция).
В анализируемом примере формы яйца возможно участие обеих корреляций. Так как по законам роста продольные диаметры тела увеличиваются сильнее поперечных, то более крупные птицы (в среднем) окажутся несколько более удлиненными, как и их органы, почему и яйцо окажется несколько удлиненным. Но, с другой стороны, в пределах одной систематической группы у мелких птиц менее выгодно отношение поверхности к массе, и естественный отбор имеет основание приближать их к шарообразной форме более интенсивно, чем крупных птиц. Мелкие утки поэтому вероятно и исторически могут оказаться более шарообразными, укороченными, чем крупные, что может привести и к укорочению яиц и к приобретению ими более округлой формы.
Для оценки наших феноресурсов очень важно подвергнуть анализу этот вопрос, так как если в установленной нами связи основную роль играет физиологическая корреляция, то достаточно будет установить. что каролинка и лебедь различаются величиной, чтобы в этот фен (наследственное различие) оказалось бы включенным и различие в форме яиц — никаких более или менее специфических частей генотипов, влияющих на расширение яиц у каролинки, искать не следовало бы. В реестр феноресурсов достаточно было бы внести лишь разницу в росте. Наоборот, если эта корреляция в основном — филогенетическая, то у каролинки наряду с признаком. «малого тела» на до будет внести и признак «широкие яйца», так как одной величиной тела физиологически объяснить ширину яйца будет уже невозможно. И если бы понадобилось для создания новой формы птиц включить в ее генотип гены, вызывающие расширение яйца, то наши исследования феноресурсов должны были бы указать как на. возможный источник этих генов уже не на савку (Erismatura), а на каролинку (Iampronessa). Ясно, что в зависимости от удельного веса физиологической и. исторической корреляции этим источником могут оказаться либо савка либо каролинка. И если мы хотим эксплоатировать наши феноресурсы не вслепую, а наверняка, го мы должны соответствующим образом подвергнуть анализу феноресурсы и вскрыть закономерности, определяющие многообразие признаков, и их коррелятивные связи;
Более детальное изложение этих вопросов должно быть отнесено уже к частной гибридагогии, к описанию отдельных гирконов.
Геноресурсы
Ресурсы генов, имеющихся в данном гирконе, представляют собой конечно наиболее интересный и важный элемент ресурсов, однако и наиболее трудно изучаемый, ибо их изучение возможно в полной мере лишь в самом процессе межвидовых скрещиваний, и притом в F
2 или F
b по крайней мере. Первое поколение, давая некоторые указания на доминантный или рецессивный характер тех или иных генов, ничего не говорит о числе генов и более или менее полном объеме действия каждого из генов. Впрочем в недавнее время Биддлем (Biddle, 1932) сделана интересная попытка анализа роли хромосом на бесплодных гибридах дрозофил.
Без применения гибридного скрещивания, т. е. в пределах вида, могут быть изучены лишь относительно немногие гены, давшие мутационные изменения. Так, в пределах группы кур мы без гибридного скрещивания знаем уже несколько десятков генов у домашних кур, пару генов у павлина, 3–4 у цесарки, столько же у индюка. Возможно некоторое дальнейшее расширение этого списка. Но несомненно, что эти гены представляют собой совершенно ничтожную часть всего числа генов, обусловливающих различия только между этими четырьмя видами — курицей, павлином, цесаркой и индюком.
С каким именно числом генов в данном примере (т. е. числом генов, связанных с различиями между 6 парами видов А — В, А — С, А — D, В — С, В — D, С — D) мы имеем дело сейчас, даже приблизительно сказать невозможно. Минимальной гипотезой будет 4, т. е. что у каждого вида имеется лишь по 1 специфическому гену, плейотропно определяющему все отличия этого вида от других. Такое расширенное допущение возможного объема плейотропии конечно ни на чем не основано. Более обоснованной будет максимальная гипотеза, допускающая, что у всех названных видов все гены отличны от генов других видов, причем часть генов гомологична, образуя серии аллеломорфов (А
g А
p A
n A
m, B
g B
p B
n B
m и т. д., где g = gallus, p = pavo, n = numida, m = meleagris), а часть может быть даже не имеет гомологов у других видов, являясь
неоморфами (Меллер).
В свете. открытия малой вероятности точной повторимости мутаций (Серебровский и др.) и открытия Тимофеева-Рессовского о различии нормальных аллеломорфов у двух географических рас дрозофилы эта максимальная гипотеза имеет за себя многое. С.этой точки зрения геноресурсы четырех видов будут иметь (помимо мутаций в пределах вида) следующий схематичный вид:
Gallus bankiva … A
g B
g C
g D
g … K
g
Pavo cristatus … A
p B
p C
p D
p … K
p
Numida numida … A
n B
n C
n D
n … K
n
Meleagris gallo pavo … A
m B
m G
m D
m … N
m
— … Гомологичная часть … Негомологичная часть
Сколько генов имеется в генотипе каждого вида, мы не знаем, но по мнению большинства генетиков число генов каждого вида должно измеряться несколькими тысячами. Тогда по максимальной гипотезе число генов, имеющихся в гирконе и составляющих его геноресурсы, определится произведением Числа генов вида на число видов. В пределах например гиркона коровы или особенно гиркона утки с 250 видами это выразится в числе, близком к миллиону.
Истина вероятно лежит где-то между обеими крайними гипотезами. Обсуждать несостоятельность первой гипотезы много не стоит; она опровергается к тому же уже биологическим опытом, обнаруживающим как правило, что в любом межвидовом скрещивании в F
2 получается трудно обозримое многообразие, сводимое к расщеплению многих генов. Что касается максимальной гипотезы, то она нами может быть принята, если составить список видов, лишь достаточно различающихся друг от друга. Но систематика иногда описывает в качестве отдельных видов (как может быть в роде Anas, — в группе Phasianus colchicus-mongolicus, во многих родах антилоп и др.) столь близкие формы, частично различающиеся может быть даже ненаследственно, что считать для каждого из этих видов все гены хотя и гомологичными, но отличными мы пока веских оснований не имеем. Вероятнее, что у этих видов значительная часть генотипа практически одинакова, а лишь по части генов имеются различия.
Впрочем, с другой стороны, совершенно несомненно, что во многих видах мы имеем наследственный полиморфизм, наследственное разнообразие, т. е. что у них некоторые гены, может быть даже многие, имеются по крайней мере в двух аллеломорфных изменениях. Многочисленные географические формы описаны у многих антилоп, оленей, лис и др., а по Гейнке даже каждый косяк сельди имеет свой собственный средний генотип, отличный от генотипов других косяков.
Таким образом, даже не имея сейчас возможности сколько-нибудь точно определить число генов в геноресурсах каждого гиркона, мы однако должны стать на ту точку зрения, что это число должно быть огромно. Овладение. этим огромным многообразием тем более заманчиво, что число комбинаций, которые возможно получить из этого многообразия, уже совершенно неисчислимо, выражаясь формулой, — близкой к М
n, где М — число видов в гирконе, а n — число генов (мест) у одного вида. Даже число 100
1000 (сто видов в гирконе с 1000 генов у одного вида), имеющее 2 000 знаков, не поддается воображению.
Метод изучения геноресурсов есть метод
генетического анализа в той однако его осложненной форме, которая связана с особенностями наследования в межвидовых скрещиваниях. Нужно иметь в виду, что изучение списка генов каждого вида будет достигнуто лишь в процессе скрещивания данного вида со многими другими видами вплоть до самых отдаленных, ибо скрещивание только двух видов, особенно более близких друг к другу, способно будет дать нам сведения лишь о части их генотипов, именно о той части, которая обусловливает
разницу между двумя скрещенными видами. При скрещивании близких видов, имеющих значительную часть генотипа общей, анализу окажется подвергнутой, лишь остальная часть генотипа, различная у обоих скрещенных видов. При скрещивании данного вида с третьим видом-анализу подвергается уже некоторая другая часть генотипа, и т. д. Чем больше будет получено скрещиваний с разными видами, тем полнее будут сведения о генотипе вида, тем полнее будет список генов.
Краткий обзор главнейших гирконов
Сколько-нибудь подробный обзор хотя бы главнейших гирконов, представляющих интерес для народного хозяйства, занял бы слишком много места и в этой части нашей книги не может быть приведен. Поэтому здесь мы дадим лишь самые краткие обзоры. В соответствии с доводами главы III мы принимаем гипотезу, что внешняя граница всего гиркона будет проходить примерно по границе семейства.
Лошади
Гиркон лошади принадлежит к числу наиболее бедных видами и многообразием. В таксономическом отношении он четко ограничивается рамками семейств лошадей (Equidae), так как ближайшие другие семейства тапиров (Tapiridae) и носорогов (Rhinocerotidae) являются уже настолько далекими и несвязанными с лошадьми никакими ныне живущими переходами, что включать их в гиркон лошади. нет никаких оснований. При этих условиях в гирконе лошади оказывается дюжина видов — лошадь, осел, полуосел, несколько видов зебр, — достаточно близких друг другу, чтобы давать гибридов. И действительно в настоящее время известны гибриды почти между всеми видами гиркона, почти во всех комбинациях, из которых лошадь × осел является самой древней гибридной комбинацией, полученной человеком и получаемой в широком масштабе и теперь. Этот гиркон состоит повидимому лишь из β-гиркона,
Несмотря на видовую бедность гиркон лошадей представляет выдающееся значение, так как качество, лошади как тягового и транспортного животного зависит от столь тонких деталей строения, что даже незначительные морфологические и анатомические отличия могут играть здесь большую роль. Основной задачей по этому гиркону является преодоление бесплодия, так как с разрешением этой задачи даже обычное скрещивание лошадь × осел, проведенное до F
2, сможет открыть огромные перспективы для улучшения как. лошади, так и осла. Однако и в пределах F
1 возможности этого гиркона еще далеко не исчерпаны, так как проводившаяся здесь гибридизация в большинстве случаев — кроме мулоразведения — еще не носила промышленного характера и вводившиеся в скрещивания лошади были обычно малоценными. Поэтому очередной задачей здесь является сравнительное изучение всевозможных F
1 с введением в него лучших (и различных — скаковых, рысистых, тяжеловозов) представителей лошадей и ослов, достоинства которых, были бы способны компенсировать недостатки диких форм, ассимилируя однако их достоинства.,
Коровы
Гиркон коровы в противоположность предыдущему представляется, наоборот, неисчерпаемо богатым, если провести его границы, по границам семейства Bovidae. Насколько мы вправе поступить так, сейчас сказать, правда, трудно, и возможно, что некоторые группы антилоп не удастся включить в гиркон коровы. Однако это семейство при всем своем многообразии столь однородно в основных признаках и столь богато различными переходными формами, что если и не прямым скрещиванием, то по крайней мере при помощи мостов можно надеяться связать весьма отдаленные подсемейства. Всего в семейство Bovidae входит около 200 видов, разделяемых на 13 подсемейств, охватывая быков, быкообразных антилоп (канна, нильгау, куду — подсемейство Tragelaphinae), конеобразных (чалая, черная антилопа), коровьих антилоп (бубал, блесток, гну), два-три подсемейства мелких антилоп (дуйкеров и др.), настоящих антилоп — газелей, сайгу, подсемейство серн, наконец коз, овец и овцебыка. Что касается подсемейства собственно быков (Bovinae): тур, зебу, як, бизон, зубр, гаур, гаял, бантенг, аноа и буйволы, — то за исключением буйвола и неисследованного аноа все остальные связываются друг с другом гибридизацией и при этом дают плодовитых самок. Это позволяет уже смело ставить гибридизацию быков подсем. Bovinae с соседним подсем. Tragelaphinae (канна, нильгау, куду и др.), в котором насчитывается еще около 15 видов. Недавно сообщалось, что получен гибрид коровы с канной и канны с куду, и таким образом создается группа уже в 25 чрезвычайно разнообразных видов, перспективы гибридизации с которыми трудно исчерпаемы: достаточно сопоставить бизона и нильгау, яка и канну и т. п. Но если подсемейство Tragelaphinae способно связаться с Bovinae, то оно будет вероятно способно связаться и с другими, уже более близкими ему подсемействами антилоп. Таким образом перспективы включения всех 200 видов в гиркон коровы нам представляются вполне реальными, особенно по мере дальнейших успехов в области искусственного осеменения и оплодотворения. Конечно это не будет простым делом. Неудачи попыток, уже многократных, получения гибридов козы с овцой кладут до некоторой степени предел. Однако уже и имеющиеся сейчас налицо плодовитые самки зубр × бизон, зубро-бизоно-тур, як × тур, як × зебу, бантенг × тур, гаял × тур и в ряде случаев и плодовитые самцы, хотя бы и не F
1, — дают такие широкие перспективы формотворчества, которые могут поспорить с ботаническими примерами.
Перечислить богатства морфологических признаков в гирконе коровы — задача, трудно выполнимая. Уже по размерам тела мы имеем диапазон от карликовых антилоп, по размерам мало отличающихся от зайца, до колоссов — дикого яка и буйвола. Такие же крайности мы имеем и по общему телосложению, сравнивая буйвола или бизона c Ammodorcas и Lithocranium (рис. 98), но эти крайности. связаны бесконечными переходами. Чрезвычайно разнообразны окраски: бизон, як, бантенг, канна, нильгау, Tragelaphus scriptus, газель, блесбок и т. д. Чрезвычайно разнообразна оброслость: зубр, як, почти голый буйвол, замечательная по оброслости антилопа Tragelaphus angasi, черная антилопа, гну, овцебык, овцы и козы. Исключительно высокое качество мяса констатировано у канны. Бесконечно разнообразные формы и величины рогов, своеобразные изменения копыт (Limnotragus), разнообразные железы и т. д. дают лишь поверхностное представление об исключительном богатстве этого гиркона, на котором должно быть сосредоточено самое напряженное внимание.
Овцы и козы
Гиркон овцы и гиркон козы, относящихся к тому же семейству Bovidae, в своих широких пределах совпадают друг с другом и с гирконом коровы, и к ним следовательно относится почти все сказанное выше. Возможно однако, что овца принадлежит к трудно гибридизируемым формам, так как известные для нее гибриды с другими видами рода Ovis являются скорее субгибридами, так как их кровь частично уже течет в домашних овцах. К сожалению до сего времени не решен вопрос о возможности получения (по крайней мере в обычных условиях) гибридов с козой. В сведениях о подобных гибридах слишком много неверного, — чтобы можно было выделить верное, необходимы дальнейшие настойчивые усилия. С козой дело обстоит не лучше, так как вряд ли верны сведения, приводимые в большой статье Гиртаннера о гибридах козы с серной, описанные, правда, достаточно живо (родятся голыми, при рождении очень, слабы и т. д.). Более ранняя заметка Манна (Mann, 1867) также сопоставляет недоразвитие волос якобы с гибридизацией. Однако гипотрихоз сохранился у этих гибридов до взрослого состояния и очевидно никакого отношения к разной продолжительности беременности у обоих видов не имеет. Никаких других признаков серны кроме «живых глаз» ни Манн ни Гиртаннер не могут указать.
Марала
Гиркон марала (пантовых оленей) также весьма интересен. Он не так богат
видами, как гиркон коровы, но уже в настоящее время известно много гибридных сочетаний оленей, позволяющих думать, что все семейство может быть действительно включено в этот гиркон. До сих пор нет сведений о гибридизации лишь с такими уклоняющимися формами, как лось, лань с ее особенным эмбриогенезом и северный олень. Но при естественном осеменении возникновение таких гибридов было либо невозможным либо маловероятным, и поэтому можно думать, что гибридизацией, как прямой, так и при помощи мостов, может быть связано в единый гиркон все семейство. В таком объеме несмотря на сравнительно небольшое число видов гиркон оленя оказывается достаточно богатым. Так, мы имеем здесь формы начиная от тропических и кончая заполярными, имеем очень большой диапазон размеров начиная от карликовых оленей Южной Америки, немногим превышающих размерами зайцев, и кончая огромным лосем. В экологическом разрезе мы имеем болотных оленей, степных, лесных, идущих как к тундровому северу, так и довольно высоко в горы; имеем ряд отдельных ценных признаков, как цепное мясо, отличные кожи и наконец получающие ныне повидимому большое значение эндокрины (пантокрин) как мощное лечебное и укрепляющее средство.
Северного оленя
Что касается гиркона северного оленя, то его особенностью на данный момент является то, что пока не известно ни одного гибрида между северным оленем и другими. Является ли это лишь чисто поверхностным результатом северного распространения этого вида, мало встречающегося с другими, или это связано с особенным Положением северного оленя в системе, — сегодня сказать невозможно, и для уяснения этого важного вопроса необходимо как можно скорее произвести пробную гибридизацию в первую очередь повидимому с наиболее близким (по Флерову) родичем — виргинским оленем.
Кролика
Гиркон кролика. Кролики и их ближайшие родичи зайцы являются основной массой, образующей один из. двух подотрядов грызунов, именно Duplicidentata, противопоставленного второму подотряду Simplicidentata, обнимающему все остальное множество семейств и родов грызунов.
В свою очередь подотряд Duplicidentata распадается на два семейства: Ochotonidae (или Lagomyidae, — пищухи, сеноставцы) и Leporidae — зайцы (см: подробный обзор остеологии, Lyon, 1904).
Зайцев также, но уже более искусственно можно разделить на две группы (Major, 1898, Weber, стр. 270, 1928): зайцев собственно и кроликов, в которую и попадает центр гиркона — домашний (и дикий) кролик (Oryctolagys clmiculus). Эта группа является повидимому более примитивной (Weber, 270), сохранившей больше сходства с упомянутым семейством сеноставцев в связи со сходным образом жизни (более короткие и почти одинаковые конечности, более короткие уши, меньшие глаза). Противоположные признаки зайцев являются признаками специализации, связанными с выработавшейся у них быстротой бега и лучшими органами чувств в связи с этим (см. Major, 1. с.).
К этой группе ближайших родственников кролика относятся Сарrolagus hispidus с южных подножий Гималаев, житель лесов Суматры Nesolagus netscheri, не роющий нор; далее бесхвостый маленький мексиканский Romerolagus nelsoni, имеющий вполне развитую ключицу, редуцированную у других членов семейств (но развитую в семействе Ochotonidae). Далее сюда же относится приспособившийся к водной жизни Limnolagus (L. aquaticus, L. palustri) восточной части Северной Америки
[18] и также американский Sylvilagus (много видов), являющихся местами бичом земледелия.
Таким образом мы имеем довольно многочисленную группу ближайших родственников кролика, к сожалению почти неизвестных советской науке и не встречающихся никогда в наших зоологических садах. Как видно из вышеприведенного, мы имеем здесь довольно разнообразные формы вплоть до «водяной», гибридизация с которыми могла бы представлять величайший интерес и возможно иметь крупное народнохозяйственное значение.
Вторая группа собственно зайцев объединяется в род Lepus с 3 подродами (Lepus, Poecilolagus, Macrotolagus). В этот род входит около 100 описанных видов, распространенных за исключением Австралии и Мадагаскара по всей земле. Мы имеем здесь и тропические формы, и полярные, и низменные, пустынные и формы альпийские (L. timidus varronis), различающиеся между собой порой достаточно сильно (Lyon, 1904; Rhoads, 1896). И здесь также нужно отметить почти полное незнакомство наших зоологов и кролиководов с этими обширнейшими мировыми ресурсами форм, которые могут быть использованы для гибридизации с кроликом и тем более с нашими зайцами.
Что касается возможностей гибридизации различных членов этого гиркона между собой, то мы имеем несомненные и нередкие случаи гибридизации наших обычных зайцев, русака (L. europaeus) и беляка (L. timidus), происходящей в природе («тумаки»). Указывали также на гибридизацию русака с альпийским зайцем. Леннберг (Lönnberg Е., 1905) считает почти несомненным, что эти гибриды плодовиты как с русаками, так и с беляками. По крайней мере после того как в Ю. Швецию были ввезены из Дании и Германии русаки, там стали часто попадаться гибриды, причем наряду с гибридами, четко промежуточными, нередки были гибриды и с сильным преобладанием беляка или русака, что Леннберг трактует как признак скрещивания F
1 × беляк и F
1 × русак. Эти выводы, основанные, правда, на тщательном изучении черепов, требуют конечно экспериментального подкрепления.
Для нас важен сейчас вопрос, насколько вообще зайцы могут быть уже сейчас включены в гиркон кролика. По этому поводу, как известно, идет спор о так называемых лепоридах. В свое время существование лепоридов, т. е. гибридов кроликов с зайцем, не вызывало сомнений. Однако затем эти лепориды были подвергнуты заслуженному сомнению. Ряд специальных опытов искусственного осеменения (так как естественное очень затруднено различием в длине задних ног) Кэстля, Ямане и Эгашпра, Геммонда и др., давших отрицательный ответ, окончательно укрепил эти сомнения (см. историю вопроса у Филипченко, 1928, стр. 77, 79).
В недавнее время Гагедорн вновь выступил с сообщением о лепоридах, где он приводит достоверные по его мнению случаи такой гибридизации; в частности в одном случае крольчиха, покрытая зайцем, дала потомство, родившееся с открытыми глазами, как у зайцев. Гагедорн подчеркнул, что более легко удается гибридизация зайца ♀ × ♂ кроликом. В письме к нам Гагедорн настаивает на несомненности этих фактов. Необходимо очевидно испробовать как различные виды кроликов, так и различные виды зайцев и добиться этих гибридов хотя бы при помощи «мостов» при точном соблюдении чистоты опыта и детальном изучении скелетных и других признаков гибридов. Охотники и любители, имеющие «ручных» зайцев, должны помочь в этом деле
[19].
Курицы
Гиркон курицы принадлежит к числу богатейших. По объему он почти совпадает с отрядом куриных. Хотя систематика этого отряда и продолжает переделываться, но по двум примерам видна его основная конструкция. Оджильви-Грант в каталоге птиц Британского музея принимает такое строение отряда:
Отряд Gallinae
Подотряд Alecteropodes:
— Сем. 1. Tetraonidae: 11 родов с 25 видами — тетерева, рябчики, северные куропатки.
— Сем. 2. Phasianidae: 59 родов со 190 видами — куры, фазаны, павлины, индейки, цесарки, перепел, куропатки, франколины и пр.
Подотряд Peristeropodes:
— Сем, 3. Megapodiidae: 7 родов с 25 видами — сорные или большеногие куры.
— Сем. 4. Cracidae: 11 родов с 55 видами — краксы, шуаны, пенелопы.
В настоящее время известны многочисленные гибриды не только в пределах семейств, не только в пределах подотряда (Tetraonidae × Phasiariidae), но между подотрядами (в данном смысле), между фазианидами и краксами. Из всей массы куриных птиц таким образом лишь небольшая груша сорных кур не дала гибридов. Но, принимая во внимание их своеобразные инстинкты, редкость в зоосадах и также то, что гибриды кур с тетеревами и кур с краксами происходили либо безо всякого участия человека либо с минимальным (завоз фазанов в районы тетеревов, воспитание краксов в зоосаду под курицей-наседкой) и что при этом полученные гибриды оказываются птицами вполне нормальной жизнеспособности — надо думать, что пределы гиркона кур окажутся еще. более широкими, чем мы можем считать сейчас. В сопоставлении с этим следует думать, что таксономическое значение названных выше групп слишком преувеличено и например более скромная оценка, принятая Мензбиром, более правильна
[20].
Отряд Galliformes
Подотряд 1. Mesites — пастушковые куропатки
Подотряд 2. Turnices — кустарные перепела
Подотряд 3. Galli — куриные
— Сем. 1. Megapodiidae — сорные куры
— Сем. 2. Cracidae — краксы
— Сем. 3. Phasianidae — фазановые
—.— Подсем. 1. Numidinae — цесарки
—.— Подсем. 2. Meleagrinae — индейки
—.— Подсем. 3. Phasianinae — фазаны
—.— Подсем. 4. Odontophorinae — американские перепела
—.— Подсем. 5. Tetraoninae — тетерева
Подотряд 4. Opisthocomi — гоацин
Во Всяком случае гиркон курицы чрезвычайно богат. По величине мы имеем, диапазон от павлина и индюка до крохотного перепела. Разнообразие форм оперения и окрасок не поддается описанию. По линии экологии имеется диапазон от тропических форм — краксы, цесарки, фазаны — до полярных, как полярная куропатка. От низменностей куры доходят до вечных снегов (горные индейки, улары), а в Гималаях поднимаются до очень большой высоты. В отношении образа жизни мы имеем лесные, кустарные, тундряные и скалистые формы и не имеем. лишь водяных и болотных. Огромен диапазон в свойствах и абсолютных и относительных размерах яиц вплоть до крупных яиц сорных кур, «искусственно инкубируемых» птицей в кучах листьев без насиживания. Яйца одного африканского франколина имеют столь прочную скорлупу, что ветер катает их по щебню, не разбивая.
В связи с различным образом и местом жизни большого разнообразия достигает характер питания вплоть до поедания глухарями в большом количестве хвои повидимому в качестве противоглистного средства (по Мантейфелю). Отсюда вероятно разнообразен и характер пищеварения, хотя по этому поводу сравнительно физиологических данных и нет.
Что касается более близких перспектив гибридизации кур, то в настоящее время может считаться окончательно установленным, что в пределах рода Gallus гибриды плодовиты, и следовательно ресурсы генов G. bankiva, G. varius, G. Sonnerati u G. Lafaieti уже могут быть соединены в единый комплекс. Что же касается дальнейших гибридных скрещиваний, в частности с богатейшей группой фазанов, то здесь прямые гибриды с курами оказываются уже бесплодными и их освоение требует преодоления бесплодия. Что касается получения гибридов, то в настоящее время работами нашей группы работников этот вопрос почти разрешен, так как искусственное осеменение позволяет уже получать гибридов с павлином, цесаркой, и несомненно мы стоим у открытия возможности и дальнейшего расширения списка гибридов, которых можно будет получать искусственно. Вопрос тут стоит лишь в таком приручении диких видов, чтобы от них можно было брать сперму, а, большинство куриных, к счастью, приручается легко и даже богатый ряд видов уже полуприручен. Таким образом перспективы гибридизации в гирконе кур можно оцепить как чрезвычайно благоприятные.
Фазана
Гиркон фазана следует упомянуть отдельно, так как хотя он целиком совпадает с гирконом курицы и Лишь слегка отличается от него положением центра, по это отличие идет в самом благоприятном направлении. В то время как центр гиркона курицы — Gallus bankiva лежит в роде Gallus всего с 4 видами и следующие виды дают уже пока бесплодных гибридов, центр гиркона фазанов, например одомашненный охотничий фазан, лежит среди обширной группы фазанов, со многими из которых он дает токонотов (см. главу VIII), и не только с многочисленными фазанами типа colchicus-mongolicus, но даже с резко отличным серебристым фазаном (Gennaeus). Поэтому перспективы гибридизации уже теперь, без преодоления бесплодия гибридов в гирконе фазанов являются очень благоприятными. На базе широкой и планомерной работы здесь могут быть Достигнуты крупные успехи и в направлении дальнейшей доместикации, акклиматизации фазанов и создания новых форм их с повышенной продуктивностью, которые сделали бы фазанов способными стать хозяйственно полезными птицами наряду с курицей, тем более что для социалистического государства исключительная красота этих птиц представляется тоже совершенно реальной ценностью для украшения общественных парков, окрестностей городов, районов домов отдыха и курортов и т. п.
О
гирконе уток мы говорить не будем, так как в разных местах многократно приводили относящиеся к нему данные.
Карпа и карася
Гирконы карпа и карася в видовом отношении являются самыми обширными, так как в семействе Cyprinidae насчитывается до 200 родов, а число видов до 800
[21], хотя в некоторых группах и имеет место увлечение систематиками возведением незначительных различий в видовое достоинство
[22]. К сожалению сейчас еще трудно сказать, насколько границы гиркона совпадают с границами семейства, однако имеющиеся данные по гибридизации карповых рыб более или менее подтверждают такое совпадение. В настоящее время в сем. Cyprinidae выделено несколько подсемейств: Abramidinae, Barbini, Rhodeini и др., и сейчас известен уже ряд гибридов не только между родами, но и между подсемействами.
Таковы гибриды:
Carassius × Barbus
Cyprinus × Tinca
Abramis × Rutilus
Leucaspius × Rhodeus и др.
Правда, некоторые подсемейства этими гибридами не связаны с другими, например подсемейства Gobiini и Schizothoracini, но здесь нам вообще ничего неизвестно о гибридах. Таким образом без риска большой ошибки можно сказать, что и здесь границы гиркона более или менее совпадают с границами семейства или по крайней мере с его основной частью. Это не значит конечно, что любой вид гиркона непосредственно можно скрещивать с карпом.
В пределах этого огромного комплекса видов внешнее многообразие, правда, очень мало. Карп, карась, золотая рыбка, линь, плотва, лещь, уклейка, голавль, усач, пескарь, горчак очерчивают рамки разнообразия в этом семействе. Тем не менее и среди перечисленных форм такие противопоставления, как карп — пескарь, карась — усач, линь — горчак, дают уже достаточно несходные пары. По величине мы имеем диапазон от карпа и леща до Leucaspius. Однако более интересны для нас физиологические различия, о которых, правда, мы можем судить лишь косвенно. Так, географически сем. Cyprinidae распространено по всему свету кроме Южной Америки и Австралии, занимая таким образом и тропические и (в основном) умеренные страны и заходя до довольно высоких широт. Будучи в основном пресноводной рыбой, карповые заходят и в море и в вертикальном направлении поднимаются в горных речках очень высоко (до 4 км). По образу жизни в этом гирконе имеются и вялые растительноядные жители тинистых прудов и жители чистых озер и каменистых речных быстрин. У некоторых мы встречаем такие поразительные биологические особенности, как откладку яиц горчаком в полость мантии Anodonta и Unio. Карась способен до 3 суток жить вне воды во влажном мху и вместе с илом подсыхать до оцепенения.
Наконец к плюсам этого гиркона надо отнести и то, что здесь в виде исключения для рыб, с двумя видами (Cyprinus carpio и Carassius с. auratus) проведена уже значительная селекционная работа, в результате которой получено многообразие как в общем строении— форме тела, так и в кожном покрове (зеркальные карпы, голые), строении плавников (вуалехвосты) и глаз (телескопы), окраске, повторяющей весьма близко весь ряд окрасок домашних млекопитающих: дикий — черный — рыжий (красный), пегий — черный по красному и пегий — цветной по белому. Уже одно соединение генофондов домашних карпов и разных рас золотой рыбки имело, бы для рыбоводства очень крупное значение, так как несомненно позволило бы выделить из расщеплений новые более хозяйственноценные линии, о частности имеющие высокую выносливость карася.
Шелкопряда
Гиркон тутового шелкопряда в настоящее время не может быть очерчен сколько-нибудь определенно. Существует всего до 80 видов шелкопрядов, шелком которых пользуется человек, и помимо того огромное количество остальных видов. Однако объединяемые русским термином «шелкопряд» бабочки эти принадлежат к разнообразным, порой далеко не родственным группам и отнюдь не все могут быть включены даже провизорно в гиркон тутового шелкопряда. Из 7 рядов семейств, на которые распадается (см. Кузнецов в книге Шарп, «Насекомые») вся масса бабочек (Hepialodea, Tineodea, Pyralidodea, Papilonodea, Lasiocampodea, Sphingodea и Noctuodea), «шелкопряды» имеются в трех последних сериях семейств. Положение семейств Bombycidae, к которому относится тутовый шелкопряд, среди этих групп следующее:
Серия Lasiocampodea 5 семейств, в том числе:
— Сем. 1. Lasiocampidae — более 100 родов и 500 видов
— Сем. 2. Dimorphidae — 1 вид
Серия Sphingodea 11 семейств, в том числе:
— Сем. 2. Eupterotidae — походный шелкопряд
— Сем. 3. Ceruridae — гарпии
— Сем. 1. Bombycidae —15–20 родов и 70 видов
— Сем. 5. Sphingidae — бражники; без кокона
— Сем. 7. Saturnidae — большое и разнообразное семейство шелкопрядов. Около 70 родов, несколько сотен видов
— Сем. 10. Geometridae — пяденицы
Серия Noctuodea 7 семейств, в том числе:
— Сем. 1. Arctiidae — медведицы, 500 родов и 3000 видов
— Сем. 2. Lymantriidae — до 200 родов и более 800 видов
Мы имеем таким образом три группы «шелкопрядов»: небольшое семейство Bombycidae, собственно шелкопрядов с 15–20 родами, каждый с небольшим числом видов, и два другие обширные семейства: Saturnidae, расположенное ближе к Bombycidae, но надо думать уже совершенно безнадежное для гибридизации с Bombyx, и еще более удаленное богатейшее семейство Lasiocampidae, содержащее до 500 видов. Последнее семейство впрочем не имеет промышленного значения. Наоборот, Saturnidae имеют сравнительно большое. значение.
Таким образом гиркон Bombyx, если его приравнять к семейству Bombycidae, будет, охватывать около 70 видов, относящихся к 15–20 родам. Однако в настоящее время у нас нет никаких оснований считать, что эти 70 видов действительно могут быть включены в один гиркон, так как никаких скрещиваний в этом семействе мы не имеем. С другой стороны, остальные виды рода Bombyx (Sericaria), а именно В. textor, В. sinensis, В. croesis, В. fortunatus и В. аггасаnensis очень близки друг к другу и повидимому уже использованы человеком при образовании различных рас домашнего шелкопряда. Таким образом приходится констатировать, что перспективы гибридизации в гирконе В. morj столь же неясны, как и объем самого гиркона.
Гиркон чесучевых шелкопрядов, принадлежащих к сем. Saturnidae, наоборот, заслуживает самого большого внимания. Род Antheraea, к которому относится центр этого гиркона, китайский дубовый шелкопряд (A. Pernyi), довольно богатый видами, также уже имеющими промышленное значение (A. yama-may, A. mylitta, A. assama и др.) и частью полуодомашненными (рис. 25). К этому роду далее примыкают достаточно близко другие (например Telea). В настоящее время уже получены гибриды между некоторыми видами рода Antheraea, так же как и между видами Saturnia в известном исследовании Штандфусса (рис. 26).
 Рис. 25. Китайский дубовый шелкопряд (Antherca Pernyi) 2/3 нат. велич.
Рис. 25. Китайский дубовый шелкопряд (Antherca Pernyi) 2/3 нат. велич.
Уоллесу (Wallace, 1869, 1870) удалось получить гибридов Antheraea Pernyi ♂ × ♀ A. yama-may (см. Bond, 1870, 1871 по данным Wallace), которые оказываются плодовитыми (Wallace, 1880). Ему же удалось получить гибридов с A. roylei. Были получены гибриды также в роде Philosamia (Ph. cinthia × Ph. arrindia) (Guerin-Meneville, 1858, 1859, и Wailly, 1893) и в роде Platysamia (виды P. gloveri, cecropia, caenothi и Columbia) с рядом интересных наблюдений над гетерозисом и исследованием видовых различий (Morton, 1895; Wailly, 1886, 1893; Meyer, 1898 и 1899). Наконец удалось получить вылупление гусеничек и при межродовом скрещивании Callosamia prometea × Philosamia cinthia (Harrison, 1908), а Штандфуссу — полувоспитать гусениц Saturnia × Actias isabella.
 Рис. 26. Тройной гибрид Saturnia (pavonia ♂ × ♀ spini) ♂ × ♀ pyri (по Штандфуссу) 2/3 нат. велич.
Рис. 26. Тройной гибрид Saturnia (pavonia ♂ × ♀ spini) ♂ × ♀ pyri (по Штандфуссу) 2/3 нат. велич.
Таким образом несмотря на значительные технические трудности, подчеркиваемые в частности Штандфуссом, дальнейшая гибридизационная работа в этом гирконе может представить значительный интерес и позволит в результате гибридизации получить более одомашненные породы и более высокого народнохозяйственного значения.
По Ватсону (Watson, 1893) гибриды Philosamia вдвое увеличивают количество шелка по сравнению с чистыми видами. С другой стороны, гибридизация в атом гирконе должна будет дать большой теоретический материал по ряду вопросов гибридагогии в целом (см. например главу о поле гибридов).
Пчелы
Гиркон пчелы по своему объему и перспективам работы в нем пока тоже очень неясен. Центр гиркона состоит из двух очень близких видов: Apis mellifera — европейской пчелы и Apis remipes — кавказской пчелы, различающихся как биометрическими константами, так и биологическими признаками — например малой «злобностью» кавказинки, позволяющей работать с ней без сетки и дыма. Эти различия впрочем трансгрессивны. Оба вида имеют локальные формы, описываемые часто как виды. Для Ленкорани и Сев. Персии Скориков устанавливает третий вид A. meda. Эти три вида объединяются и в секцию III подрода Apis. Вторая секция из двух видов (adansoni и unicolor) занимает Африку, а первая секция — Ю.-В. Азию (виды johni, cerana или sinensis indica и japonica). Эти 9 видов ближайших родичей домашних пчел составляют. подрод Apis того же рода, а весь род обнимает и еще два индийские вида (florea и dorsata). Если некоторые виды очень близки к домашним, то уже cerana отличается ясно и по некоторым указаниям не скрещивается с mellifera («Пчелов. дело»,1928, № 7, 370). Наконец florea и dorsata отличаются очень сильно, особенно dorsata, выделявшаяся Ашмидом в особый род.
Далее в сем. Apidae имеется еще целое подсемейство Meliponini с большим, недостаточно изученным числом видов мелких нежалящих пчелок-мелипон (роды Melipona, Trigona, Tetragona). В одной лишь долине Амазонки их насчитывают не менее 45 видов.
Возможно ли скрещивание домашних пчел с видами других секций, подродов и с подсемейством мелипон — сейчас совершенно неизвестно за. отсутствием экспериментов. Между тем мощное оружие искусственного осеменения проникло уже и сюда, и отсутствие данных об естественной гибридизации этих видов ориентировать нас уже не может. Несомненно однако, что богатейшие ресурсы этого семейства еще совершенно не достаточно использованы человеком.
Глава V. Проблема гетерогенного осеменения
Первой задачей, которую приходится решать гибридизатору в его работе по получению гибридов, является задача гетерогенного осеменения. Под этим термином мы будем разуметь
совокупность всех тех процессов и манипуляций, которые необходимы для доведения сперматозоидов одного вида до яйца другого вида, относя тем самым все вопросы, связанные с активным приближением сперматозоида к яйцу, внедрением сперматозоида в яйцо и возбуждением яйца к развитию, к следующей проблеме — проблеме гетерогенного оплодотворения, которое мы будем рассматривать в главе VI.
По отношению к некоторым группам животных, например к рыбам и другим формам с внешним оплодотворением, проблема гетерогенного осеменения оказывается достаточно простой, сводясь лишь к одновременному получению зрелых яиц и сперматозоидов и смешению их. Наоборот, у других видов — млекопитающих, птиц, насекомых — эта задача значительно труднее, и разрешение ее сталкивается с рядом препятствий механического, физиологического и рефлексологического порядка. В настоящее время, правда, благодаря успехам искусственного осеменения смогли найти способы преодоления некоторых из этих препятствий. Но там, где искусственное осеменение еще не разработано, например у насекомых (кроме пчел), эти препятствия остаются еще очень значительными. Поэтому проблема гетерогенного осеменения требует еще значительной теоретической и технической разработки и, как увидим ниже, оказывается связанной с широким комплексом самых разнообразных проблем экологии, рефлексологии, физиологии, биохимии и т. д.
Гетерогенное осеменение может быть достигнуто путем или естественного полового акта или искусственного осеменения. В обоих случаях явления, связанные с прохождением сперматозоида до яйца, остаются одинаковыми. Однако искусственное осеменение открывает значительные перспективы в смысле возможности сокращения для сперматозоида длинного «естественного» пути от влагалища до яйца. Наоборот, начальные стадии осеменения в обоих случаях достаточно различны. Используя путь естественного полового акта, мы должны заставить спариться самца и самку, принадлежащих к разным видам. Во многих случаях это может оказаться чрезвычайно трудным или даже невозможным. Но даже и в тех случаях, когда самец идет на самку, естественный половой акт может встретиться с механическими препятствиями, о которых говорилось в III главе, и введение семени во влагалище окажется невозможным.
Искусственное осеменение устраняет эти препятствия, так как оно может пользоваться естественной садкой самца на самку его же вида и подменяет эту самку самкой другого вида или чучелом лишь тогда, когда это оказывается удобнее. Наконец в некоторых случаях искусственное осеменение обходится вовсе без нормального полового акта, заменяя его механическими или хирургическими манипуляциями.
С другой стороны, искусственное осеменение встречается со своими специфическими задачами и трудностями в виде получения спермы в таком виде, чтобы с ней можно было дальше манипулировать, с задачами введения спермы в организм самки, с хранением спермы и т. д.
Не может быть однако сомнения в том, что серьезный расцвет гибридизации может произойти лишь на базе искусственного осеменения, в котором гибридизатор перестает быть кустарем и становится полным хозяином положения. Поэтому в основу дальнейшего изложения мы должны положить именно эту ориентацию, и с этой точки зрения стоящие перед нами проблемы могут быть разделены на три группы:
1) проблема получения спермы от самца;
2) проблема своевременного введения спермы в самку или вообще доставления к яйцу;
3) проблема доведения самки до размножения (овуляции).
Получение спермы
Получение спермы возможно тремя способами:
а) хирургическим методом,
б) провоцированием эякуляции,
в) нормальным половым путем.
Хирургический метод
Хирургический метод добывания спермы сводится к проникновению в организм самца и изъятию гонад, эпидидимиса, семяпровода и других вместилищ спермы, к устройству фистул и введению канюль и пр. Так, при гибридизационных опытах с морскими ежами и другими иглокожими вскрывают зрелых самцов по экватору и вырезают обширные семенники. Также получают сперму у трутней пчел, у мышей, которые слишком мелки для того, чтобы можно было применять другие методы. Геммонд получал так сперму у зайцев для гибридизации с кроликом, Домм — у петухов. Румянцеву отлично удалось осеменение стада овец спермой дикого барана, застреленного в горах за несколько десятков километров. У млекопитающих достаточно зрелые сперматозоиды находим в эпидидимисе; расщипывая его в физиологическом растворе или других более сложных средах, можно приготовить искусственную сперму, годную для осеменения ею самок.
Этот метод однако является в большинстве случаев слишком расточительным, так как позволяет использовать самца либо однократно либо самое большее — двукратно (сначала один семенник, затем другой). Там, где — самцы очень многочисленны и дешевы (например у пчел), эта расточительность может быть допущена. Но по отношению к более крупным и дорогим животным на хирургический метод в подобной форме можно итти лишь в крайнем случае, если иные пути оказываются бессильными.
Однако нужно иметь в виду, что сказанное относится лишь к тому, самому примитивному хирургическому приему, который применялся до сих пор, — простое вырезывание эпидидимиса и других частей полового аппарата. Эта примитивность конечно вовсе не обязательна. Наоборот, можно считать вполне реальными перспективы разработки таких операций, которые позволили бы получать сперматозоиды постоянно и без вреда для животного. В качестве такой задачи должно быть поставлено образование фистул эпидидимиса или семяпроводов с образованием резервуара для выделяемых сперматозоидов и с возможностью периодической пункции их. Нежность и небольшие диаметры семяпроводов делают эту задачу конечно нелегкой, но разработанные методы фистул и выведение наружу желез слюнных и других позволяют считать эту задачу не безнадежной, особенно но отношению к более крупным животным.
Разработка этого направления в хирургическом методе дала бы наиболее простые и удачные возможности систематического получения спермы, снимающие целый ряд трудностей. Особое значение этот метод может иметь именно для гибридагога, так как проблема получения спермы у самцов диких видов животных встречается нередко с особыми трудностями.
Провоцирование эякуляции
Провоцирование эякуляции возможно двумя методами; 1) механическим раздражением пениса (мастурбацией) и 2) электрическим током.
 Рис. 27. Получение спермы мастурбацией у лиса (фото Аскании-Нова).
Мастурбация
Рис. 27. Получение спермы мастурбацией у лиса (фото Аскании-Нова).
Мастурбация является одним из самых простых способов получения спермы, но к сожалению пока она удается лишь у немногих видов и в настоящее время применяется исключительно к видам сем. Canidae — собаке, лисице, корсаку, енотовидной собаке (Дубровский). Захватив сбоку рукой препуциум, несколькими движениями заставляют самца выпустить головку пениса, после чего самец переступает ногой через руку экспериментатора и пенис оказывается повернутым назад, как ясно видно на прилагаемом рис. 27. Ритмическими надавливаниями на пенис проксимальнее вздувшейся бульбы получают дальнейшее усиление эрекции и наконец постепенную эякуляцию, причем сначала вытекает собственно сперма, а затем секреты добавочных желез. В целях уменьшения раздражения и может быть болезненных ощущений, вызываемых непосредственным соприкосновением руки с нежными частями пениса, Дубровским разработан «закрытый способ». Для этой цели сконструирован простой приборчик, приближающийся к «искусственной вагине» (см. ниже) и состоящий из резиновой достаточно широкой и мягкой трубки, в один конец которой вставлена делительная воронка со стеклянным краном. Резиновая трубка надевается на головку пениса и его бульбы, когда пенис повернут назад. Вздувшиеся бульбы натягивают стенки трубки, герметически закупоривая ее. Экспериментатор, надавливая ритмически на резину, вызывает эякуляцию, предварительно выпустив через кран сдавленный в аппарате воздух. Этим способом Дубровскому удавалось получить от лиса до 15–20 см
3 спермы за сеанс, причем сперма получается совершенно чистой, чего трудно достичь при «открытом способе».
Применение мастурбации требует предварительного приручения животного, и в этой предварительной работе заключается основная трудность, метода. Однако после того как первые опыты удались, приручение прогрессирует. Лисы в Аскании-Нова, с которыми работает Дубровский, по его зову сами вспрыгивают на стол и неподвижно стоят во время манипуляции, лишь слегка придерживаемые за корень хвоста. Однако все нее для удачи необходима известная обстановка, особенно в начале работ, — отсутствие посторонних лиц, предпочтительно ночь, тишина. Стук двери способен моментально прекратить эрекцию.
У енотовидной собаки пенис не отгибается назад, и техника работы соответственно несколько изменяется.
Электроэякуляция удавалась у морских свинок и крыс. Фиксированному животному один полюс прикладывается к нижней челюсти (в рот) в форме металлической пластинки. Другой полюс в форме иглы вкалывается под кожу в затылочной области. Ток напряжением около 30 вольт пропускается до наступления эякуляции, для чего необходимо около 20 секунд. Электрический ток достаточно тяжел для животного, хотя Кабаку удавалось неоднократно повторять его на одном и. том же самце (морских свинок) с промежутком в 10 дней. Механизм электроэякуляции еще мало изучен, и вопрос о возможном расширении объектов его применения и облегчения шока подлежит дальнейшей разработке. В частности получить электроэякуляцию у мышей и быков нам не удалось — слабые удары не давали эякуляции, а более сильные убивали животное. Груздеву не удались опыты с кроликами.
Наоборот, с птицами нам удалось недавно получить достаточно благоприятные результаты. Использовался постоянный ток (очень удобно — от «анодной батареи», употребляющейся для радио) силой около 30–45 вольт. Один полюс в. виде хирургической иглы вкалывался под кожу крестца или при повторных опытах в виде мокрой подушечки прижимался к освобожденной от перьев коже крестца. Второй полюс опускался в миску с водой. Замыкание производилось погружением клюва птицы в миску. Вариированием расстояния, клюва от провода (опущенного в миску) и глубины погружения легко изменять силу удара. Обычно после 1–2 минут слабого тока (включением 45 вольт) давалось до 4 более сильных ударов (включением всех 80 вольт батареи). При хорошем состоянии птицы уже при слабом токе, а обычно при ударах, выделяется небольшое количество спермы, которое может быть собрано пипеткой (Серебровский и Соколовская, 1934). Таким способом нам удалось получить сперму у селезня, гусака, цесарок и различных фазанов. При этом у группы цесарей получение спермы производилось регулярно в течение почти 3 месяцев (у одних — почти ежедневно, у других — через 1–2 дня). Сперма получается очень чистой и нормального качества, но к сожалению в небольшом количестве (не больше нескольких кубических миллиметров), что является пока недостатком метода. Однако перспективы этого метода несомненно чрезвычайно велики. Метод электроэякуляции быстр и в принципе прост, не требует приручения, что особенно важно при работе с дикими животными. В этом отношении он мог бы быть столь же универсален, как и метод кастрации, допуская однако многократное использование самца, что является конечно чрезвычайно ценным. Малая изученность этого приема не должна нас смущать. То, что в этой области очень мало известно, указывает лишь на то, что она стояла вне поля внимания исследователей. Развитие гибридизации эту проблему выдвигает на очередь, и она должна быть разрешена в ближайшее время.
К провоцированию эякуляции должно отнести и простой прием получения спермы у рыб путем надавливания на брюшную область зрелых самцов. Самца берут под жабры одной рукой так, что он висит хвостом вниз, и другой рукой охватывают пальцами туловище и, слегка надавливая, двигают сверху вниз. Зрелые молоки легко выливаются в подставленный сосуд.
Получение спермы нормальным половым актом
Получение спермы нормальным половым актом пока остается основным методом. В понятие «нормальный» мы вкладываем однако только то, что наиболее важный для нас момент эякуляции вызывается рефлексом без посторонних механических или электрических раздражений. Детали же полового акта могут быть разнообразны и в значительной степени искусственны, проходя например при ближайшем участии человека с применением специальной аппаратуры для собирания спермы, с заменой живой самки чучелом с искусственной вагиной и тому подобными техническими ухищрениями. В настоящее время в этой области имеется уже довольно богатый опыт, который интересно суммировать. В этом вопросе необходимо различать две стороны: 1) получение садки и эякуляции и 2) сбор эякулята — спермы.
Рефлексология садки
Нормальный половой акт (будем называть его садкой) является результатом действия сложного безусловного рефлекса (инстинкта), а при повторных садках — и условных рефлексов. Для того чтобы этот врожденный инстинкт начал действовать, необходимо воздействие на рефлекторный аппарат двух категорий раздражений — внутренних, связанных с физиологическим состоянием самца и создающих необходимый тонус, и внешних, поступающих на головной мозг через органы чувств — зрение, обоняние, слух и пр. Хотя эта общая картина и ясна, но множество важнейших вопросов остается еще неизученным, а между тем без ясного понимания этих деталей мы не будем в состоянии овладеть этим инстинктом так, чтобы в руках гибридизатора он работал без отказа.
Категория внутренних раздражений чрезвычайно важна, и при их отсутствии инстинкт не проявляется. Эти раздражители в основном повидимому гормонального, а также зависящего от питания характера, если говорить о млекопитающих и птицах. Как правило взрослого самца можно посадить на самку лишь в том случае, если у него нормально работает эндокринная система- или во всяком случае некоторые звенья ее. Удаление семенников, кастрация, устраняет эти раздражения почти нацело. Об условных рефлексах скажем ниже. У большинства млекопитающих и птиц, особенно у диких, эта эндокринная система работает циклически и сезонно, и необходимый для работы полового инстинкта тонус создается лишь в известное, иногда очень короткое время года. Вне этих сроков антагонизирующие эндокринные железы подавляют инстинкт, и по крайней мере у некоторых видов садка становится невозможной. Точно так же наличие этих раздражителей ограничено определенным возрастным отрезком. Они отсутствуют в достаточно развитой степени до наступления половой зрелости и угасают с окончанием или ослаблением деятельности некоторых эндокринных желез.
Для целей гибридизации важна конечно садка только в соответствующий сезон, так как при бездействии половых желез получение садки теряет свой смысл. В некоторых случаях поэтому гибридизатор может быть крайне заинтересован не только в получении самой садки, но и в возбуждении деятельности половых желез. Этому важнейшему вопросу будет посвящена глава VIII. Сейчас же мы предполагаем, что в нашем распоряжении находятся самцы уже с наличием этих внутренних раздражителей и для получения садки необходимы лишь соответствующие внешние раздражения.
Каковы те внешние раздражения, которые необходимы для того, чтобы пустить в ход инстинкт садки, — вот тот чрезвычайно важный вопрос, который необходимо решать гибридизатору и притом решать не только вообще, но и конкретно по отношению к каждому виду животных, с которым ему приходится работать. Эта конкретность необходима уже потому, что у кап? до го вида животных эти внешние раздражения настолько специфичны, что в естественных условиях в подавляющем большинстве случаев самец спаривается только с самкой своего вида. Отсюда можно было бы заключить, что эти раздражения связаны всецело с самкой того же вида. Однако, что это не так, показывает уже весь богатый материал естественной гибридизации и еще более — гибридизации в условиях зоосадов и животноводческих хозяйств, где часто происходит гибридизация в самых неожиданных комбинациях и в еще более неожиданных комбинациях происходят попытки к спариванию. Так, нередки случаи попыток селезней спариваться с курами или петухов с утками, нередки попытки двух самцов спариваться друг с другом, быков — с волами-буйволами, и т. д.
Эти случаи показывают, что не все в самке необходимо для вызывания садки со стороны самца.
Еще большее недоумение вызывает на первый взгляд разработанная ЛИО ВИЖ
[23] методика садки самцов домашних животных на чучело. Такое чучело может весьма отдаленно напоминать живую самку (неподвижно, иного запаха), а тем не менее например быки, бараны и хряки охотно идут в садку на чучело и даже более — на чучело, имитирующее корову, может охотно итти и жеребец (рис. 28), а индюк может делать садку на положенную перед ним шапку или другой предмет, ничего общего с птицей не имеющий.
 Рис. 28. Садка жеребца на чучело короны (фото ЛИО ВИЖ).
Рис. 28. Садка жеребца на чучело короны (фото ЛИО ВИЖ).
Если мы будем искать нечего общее во всех этих раздражениях, то нам будет совершенно непонятно, почему же в естественных условиях животные не спариваются в самых фантастических комбинациях уже не в виде исключения, а как правило.
Для того чтобы разобраться в этих вопросах, необходимо прежде всего различить роль безусловных рефлексов и рефлексов условных. Под первыми подразумеваются рефлексы врожденные, связанные с врожденными нервными структурами, благодаря чему раздражение
А влечет за собой со стороны животного реакцию
В. Под условными рефлексами, открытыми и изученными школой ак. И. П. Павлова, разумеются рефлексы, устанавливающиеся уже в течение индивидуальной жизни организма и как таковые не передающиеся но наследству через яйцо и сперматозоид. В общей форме механизм установления условного рефлекса таков: если раздражение
А безусловно вызывает реакцию
В и если раздражение
С не вызывает реакции
В, то при многократном совпадении во времени раздражения
С с раздражением
А первое включается в рефлекторную цепь — и в конце концов это включение оказывается настолько сильным, что раздражение
С получает возможность вызывать реакцию
В даже в отсутствии раздражения
А, требуя лишь последующего подкрепления раздражителем
А, без чего условный рефлекс подвергнется «торможению».
В интересующем нас явлении спаривания картина установления условного рефлекса может приобрести например такой вид. В порядке безусловного рефлекса селезень кроет утку. Но эта утка сидит в отдельной клетке. Ежедневно приходит экспериментатор и берет утку из клетки, пересаживает ее к селезню, после чего и происходит спаривание, и утка запирается обратно в клетку. Многократное совпадение этих событий приводит к тому, что одно появление экспериментатора приводит селезня в состояние полового возбуждения, и когда экспериментатор опускает к селезню руки с находящейся в них уткой, селезень тут же в руках кроет утку. Явление
С (появление экспериментатора) приобрело характер условного раздражителя, и селезень продолжает совершать половой акт и тогда, когда в руках экспериментатора оказывается уже не утка, а ее чучело, и вместо клоаки его пенис попадает в стеклянную баночку. Если бы мы упустили из виду, что чучело стало объектом садки, лишь включившись в безусловную рефлекторную цепь: утка — мозг селезня — садка, то мы бы ошибочно приписали чучелу вообще способность вызывать работу инстинкта садки.
Описывая опыты посадки жеребцов на
чучело, авторы ЛИО отмечают (стр. 346): «Особенно интересно в этих опытах то, что чучело переносилось и ставилось в разных местах, в том числе и в открытой степи, в силу чего отпадают соображения об условнорефлекторной связи полового акта с определенной обстановкой и местом».
Наблюдения свои авторы резюмируют следующим образом:
«1. Для полового возбуждения самца не является обязательным специфический раздражитель в виде наличия охоты у самки и даже вообще наличия самки. Возбуждение самца получается и в присутствии
чучела самки, а также самца или кастрата, поставленных в соответствующую обстановку.
2. Обычное представление о том, что для возбуждения самца. необходим
запах находящейся в охоте самки, не подтверждается нашими опытами. Садка производителей на чучела из выделанных шкур, на чучела, обтянутые искусственными материалами, а также на самцов и кастратов подтверждает это достаточно определенно. Тем не менее самцы и при садках на чучело предварительно долго обнюхивают их и даже воспроизводят характерную мимику (поднятие головы и вздергивание верхней губы). Мы считаем этот положительный рефлекс, утративший свое физиологическое значение, пережитком. Возможно, что в условиях вольной стадной случки обонятельное ощущение помогает самцу отыскивать приходящих в: охоту самок.
3. Повидимому основным фактором, возбуждающим производителя, является
зрительное впечатление, которое может быть связано с некоторыми другими условнорефлекторными раздражителями (место, обстановка). Это зрительное впечатление не является особенно точным, как об этом свидетельствуют опыты садки жеребца на чучело коровы. Не ясно еще значение цветовых восприятий».
С этой точкой зрения невозможно согласиться. Нужно иметь в виду, что все эти наблюдения делались над животными, которые были предварительно приучены к ручной садке и вследствие этого имели целую установившуюся систему условных рефлексов. Опыты садки жеребцов на чучело коровы в открытой степи вовсе не опровергают этого: и там обстановка случки была вполне типичной. Жеребца на двух длинных поводах конюха подводили сзади к чучелу совсем так, как это делается при ручной случке: перед жеребцом стояло нечто сходное с кобылой. Правда, ряд элементов здесь отпадал: запах и некоторые детали зрительных; впечатлений. Поэтому эти опыты показывают лишь, что условные рефлексы могут приобрести столь большую силу, что могут подменить некоторые безусловные раздражители.
Анализ же того, какие именно безусловные раздражители вызывают
первую садку и какие могут быть опущены, может быть, исследован лишь на самцах, впервые идущих в ручную случку и садку вообще.
Для уяснения вопроса о различии между первым и дальнейшими-спариваниями большой интерес представляют следующие факты, приводимые Л. Морганом (Морган, 1899, стр. 200): в уединенной сосновой роще Востер нашел пару кардиналов (Piranga rubra) и застрелил самца. Самка уже через 2 дня имела другого самца, который снова был убит, и таким образом в течение недели было убито 4–5 самцов, причем «эти самцы, расположенные в том порядке, в каком они были убиты, составляли постепенный ряд, в котором первая птица была необыкновенно ярко окрашена, последняя удивительно невзрачна». Другой наблюдатель добыл трех самцов краснокрылых дроздов, летавших последовательно над одним и тем же гнездом. Первый был очень ярко окрашен, второй. менее ярко, третий был очень молод, а с самкой остался четвертый, вылупившийся лишь в прошлом году, судя по отсутствию красного цвета на крыльях.
Эти факты, которые конечно совершенно недостаточны и недостаточно строги, рассматриваются как доказательство в пользу того, что яркость окрасок способствует выбору самкой самца (хотя в данных примерах возможно, что более молодые самцы просто прогонялись более сильными и взрослыми). Но они могут быть использованы и как примеры того, что для первого спаривания необходимо большее напряжение раздражителя (в данном случае со стороны самца), а после того как у самки установились уже и условные рефлексы на всю обстановку спаривания, отдельные элементы самца — яркость его окраски — уже утрачивают значение. Было бы очень важно собрать более серьезный наблюдательный и экспериментальный материал по этому вопросу.
Под этим углом зрения становится понятным и парадоксальный факт обнюхивания, наблюдавшийся в ЛИО. Известно, что быки перед садкой, обнюхивают половые органы самки с характерным вздергиванием верхней губы. Ясно, что запах играет определенную роль в возбуждении инстинкта садки, но при работе с чучелом совершенно такая же реакция обнюхивания была получена на струю чистой воды. Так как вода не пахнет, то получилась странная картина, ставящая как будто под полное сомнение роль запаха. Но если имеется в виду, что здесь уже произошла подстановка безусловных раздражителей условными, можно понять, что эти условные раздражители вызвали садку несмотря на то, что безусловные уже отсутствовали.
Таким образом мы должны притти к выводу, что гибридизатор должен четко различить проблему условных раздражителей от проблемы безусловных раздражителей, вызывающих садку. Сейчас уже совершенно ясно, что в работе с животными для получения садки мы можем в чрезвычайно широкой степени произвести подмену безусловных раздражителей условными, заставив самца делать садку на тот предмет, на который нам надо его посадить по техническим соображениям. Но нужно помнить, что это возможно лишь в порядке включения в безусловную рефлекторную цепь условных раздражителей. Этими условными раздражителями можно легко и удобно пользоваться при дальнейших садках. Но они ни в какой мере не снимают
проблемы безусловных раздражителей, необходимых для получения первой садки, начиная с которой мы уже можем устанавливать систему условных рефлексов.
Чучельный метод. ЛИО разработан метод садки самца млекопитающих на чучело. После приучения самца (жеребец, бык, хряк, баран) к ручной садке на самку можно без особого труда посадить его на чучело (рис. 28). Чучело устраивается из прочного каркаса, обтянутого мягким и обернутого шкурой или еще лучше — сукном, так как сухая шкура издает барабанный звук, пугающий самцов при садке. Сходство чучела с самкой может быть весьма относительное. Как упоминалось, жеребец идет и на чучело коровы. В опытах Веревкиной и Скаткина «быки симменталы с первого же раза охотно пошли на чучело несмотря на то, что оно было еще не доделано: сзади ниже вымени торчала деревянная основа, голова не была зашита, из-под кожи торчали стружки, кроме того кожа после выделки имела специфический запах квашения».
В вопросе
о безусловных рефлексах садки основным является вопрос о том, в какой степени они связаны 1) с комплексом свойств и признаков самки и 2) с комплексом элементов естественной экологии этого вида. Что не все свойства самки и не все элементы экологии являются важными, в этом можно не сомневаться. Поэтому подлежит выяснению чрезвычайно интересный и широчайшего значения (для эволюционной теории) вопрос, какие именно свойства самки необходимы и достаточны для провоцирования самца на садку и какие элементы окружающей обстановки необходимы и достаточны для этого. Ясно, что этот вопрос должен решаться особенно конкретно для каждого вида путем специальных наблюдений и экспериментов.
При этом однако важно и необходимо рассматривать не только изолированно взятую самку, но и систему самка — самец, ибо поло-вой акт происходит именно в этой системе (оставляя пока в стороне окружающую обстановку). Если например внимательно присматриваться к процессу спаривания индюка и индюшки, то можно видеть следующий ряд явлений в этой системе.
1. Одиноко гуляющий индюк кричит.
2. Самка отвечает, индюк кричит в ответ, и т. д. Началом может явиться голос самки или вид ее вдали.
3. Самка идет к самцу, самец — к самке.
4. Самец, видя самку вблизи, начинает токовать — распускает оперение, надувает зоб, наливает кровью головные украшения, начинает характерные движения.
5. Видя и слыша токование индюка, индюшка приседает.
6. Индюк садится на самку, начинает топтать ее.
7. Самка поднимает хвост и повидимому выворачивает отверстие яйцевода.
8. Самец изливает сперму в отверстие ватины.
 Рис. 29. Садка индюка (фото Аскании-Нова).
Рис. 29. Садка индюка (фото Аскании-Нова).
В этом ряду, на самом деле еще более сложном, имеется цепь безусловных рефлексов, вернее фаз единого полового рефлекса (инстинкта): фаза подачи голоса, фаза, приближения на голос другого пола своего вида, фаза токования при виде самки, фаза приседания при виде токования самца, фаза садки на присевшую самку и топтания ее (рис. 29), фаза ответа на топтание и т. д. В естественной обстановке вся эта цепь рефлексов необходима для получения coitus’a. Однако будет ошибкой думать, что вся эта цепь необходима всегда, и при желании заставить индюка дать сперму в первый раз необходимы все элементы индюшки — ее голос, ее вид, окраска, ее размеры, приседание и пр. Например, что будет делать индюк, если на его голос приблизится к нему не индюшка, а другая птица (случайно или по желанию экспериментатора)? В какой степени должна быть эта птица похожей на индюшку, чтобы вызвать токование? В чем она должна быть похожа”—в окраске, голосе, «манерах»? Каково будет поведение индюка, если мы заставим сесть перед ним другую птицу, и в какой степени и в каких элементах должно быть это сходство? и т. д. Каждый из этих вопросов требует анализа, и, проделав его, мы получим ответ, какие именно элементы необходимы и достаточны для приведения в действие инстинкта садки индюка.
Наблюдения показывают например, что столь резкое изменение вида индюшки или индюка, как замена их нормальной окраски белой или желтой, не оказывает никакого (повидимому) влияния на ход цепи рефлексов, перечисленных выше
[24].
Точно так же нередко токование начинается при приближении курицы или даже в полном одиночестве (возможно однако, что при реакции на голос другого индюка или индюшки). Наоборот, большое значение несомненно имеет голос
[25]. По нашим наблюдениям в вызывании садки решающее значение имеет приседание самки, которое может быть легко вызвано например при помощи веревки, привязанной к ноге самки.
При потягивании за веревку индюшка присаживается, и индюк немедленно садится на нее. Учтя это наблюдение, мы допускали к индюку курицу и веревкой заставляли ее присаживаться или даже падать на бок, вызывая со стороны индюка реакцию садки. (Приводя эти примеры, мы оговариваемся, что имели дело не с первой садкой.) Таким образом из множества свойств индюшки одним из наиболее решающих является ее свойство реагировать на токование индюка приседанием. Выяснив этот решающий элемент, — мы оказываемся в значительной мере хозяевами положения, будучи в состоянии нажимать необходимую пружину, приводящую в действие инстинкт садки у индюка.
Не менее поучительна игра половых рефлексов при спаривании павлинов (рис. 30). Она состоит из следующих фаз:
1. Увидев самку-паву, павлин распускает хвост, поворачивается к самке задом и начинает «стрекотать» распущенными до земли крыльями, попеременно двигая ими; иногда трясет веером «хвоста».
2. Через некоторое время самка приближается к самцу, затем
бежит, обегает его по широкой дуге и на, некотором расстоянии перед ним, хвостом к нему, приседает.
3. Самец, издав характерный крик и постепенно опуская, но не складывая веер, бросается на самку, хватает ее за перья затылка (но не за хохол) и вскакивает на спину, опуская веер на землю. Топчет (проверить!) самку.
4. Самка в ответ на топтание поднимает хвост (? и выворачивает яйцевод).
5. Самец изгибается, прикладывает клоаку и извергает семя.
6. Самец вскакивает и, снопа распустив веер и обернувшись к самке задом, некоторое время снова токует («нахшпиль» немцев, см. Heinroth об утках
[26]).
Первое звено довольно расплывчато, и павлин легко токует перед курами, индюшками и т. д. Наиболее ответственной и обычно решающей является вторая фаза, оканчивающаяся приседанием павы. Физиологический механизм этой фазы настолько специфичен, что конечно вызвать на него самку другого вида (кроме видов p. Pavo) павлин уже не может. Повидимому однако в некоторых случаях игра идет несколько проще, а иногда павлин бросается на самку без первой и второй фазы, почему изредка садится и на кур.
 Рис. 30. Токование павлина.
Пава бежит, чтобы присесть перед павлином (фото Аскании-Нова).
Рис. 30. Токование павлина.
Пава бежит, чтобы присесть перед павлином (фото Аскании-Нова).
Совершенно иначе обстоит дело у петуха. Здесь, так же как и у индюка, внешний вид самки имеет ничтожное. значение — петух готов крыть курицу любой породы вплоть до белых, голошеек, курчавых и пр., даже в первую садку. Также как и у индюка, огромное значение имеет голос — от закричавшей цесарки петух может испуганно отскочить (возможно однако, что это уже условный рефлекс; необходимо исследование воспитанных вместе петухов и цесарок-самок в отсутствии другого пола своего вида). Но очень большое значение имеет движение. Если курицу
бросить петуху с некоторой высоты, наступает обычно немедленная садка. Легко посадить петуха таким образом и на цесарку и на чучело, хотя потом петух может и соскочить, например услышав крик цесарки. Поэтому же индюк не может покрыть курицу, так как не может заставить ее присесть.
Утверждение, что индюк не может покрыть курицу, впрочем не абсолютно. Мы наблюдали следующее явление. В маленькой вольере, где индюк ходил вместе с курами (а за решоткой помещались индюшки), индюку иногда удавалось заставить курицу сесть. Мы видели, как однажды индюк, токовавший среди кур и напиравший на одну из них, вдруг клюнул ее в голову, и курица моментально присела в той типичной позе (со слегка приподнятыми крыльями), в какой она присаживается перед догнавшим ее петухом. На такую присевшую курицу индюк уже довольно спокойно взобрался и начал ее топтать ногами (спаривание не доходило до контакта клоак благодаря большой разнице в размерах, хотя эякуляция и наступала). Это явление нами наблюдалось неоднократно и над несколькими курами. Также иногда удается и павлину посадить курицу. Интересно отметить, что рефлекторное приседание происходит у многих кур и тогда, когда их настигает ловящий их человек. Некоторые из кур приседают очень легко, — стоит лишь создать ей, ловя ее, более или менее «безвыходное» положение.
Вопрос о взаимном возбуждении самца и самки на самом деле значительно сложнее, так как нередко полы могут сильно изменять свое нормальное поведение трудно поддающимся объяснению образом. Гейнрот (Heinroth, 1910) сообщает следующие наблюдения над Lampronessa sponsa: «так как этой весной мы имели (в Берлинском зоосаду) значительно больше самцов, чем самок, то оставшиеся холостыми самцы образовали пары и держались неразлучно вместе, как настоящие пары. Желая спариться, оба начинали с типичных самцовых приемов (попивание — Antrinken), но наконец один принимал на себя женскую роль — подобно самке ложился плашмя на воду, а второй самец взбирался на него и спаривался самым нормальным образом». В 1933 г. в Московском зоопарке широконоски (Sp. clypaeta) спарился с самцом же серой утки (Ghaul. streperus) и неотлучно плавал с ним. Мантейфель наблюдал и типичную садку.
Если, наоборот, в вольере имеются в избытке самки или если самцы не активны, то спариваться начинают две самки (мы наблюдали то же у индюшек, уток и одна взбирается на другую, начинает топтать ее и ведет себя совершено по-селезиному. Замечательно, что она производит и соответствующий «нахшпиль», какой проделывает самец после садки
[27]. Конечно по анатомическим причинам отсутствует лишь «повисание» (Hangen).
Совершенно то же наблюдали и мы на домашних и на мускусных утках.
Как отмечает Гейнрот, наиболее интересно, что здесь самец принимает на себя совершенно женскую роль и наоборот, вопреки нашему представлению о том, что особенности поведения зависят от физиологии пола. Никакого гомосексуального изменения физиологии здесь нет, так как описанные явления происходят лишь при недостатке другого пола и немедленно исчезают, когда восполняется этот недостаток.
Надо думать, что в нормальном самце (и самке) имеются рефлекторные механизмы для поведения как в качестве самца, так и в качестве самки, но поведение противоположного пола подавляется «своей» категорией рефлексов. Тем не менее остается очень темным вопрос о том, что именно заставляет самца принимать на себя роль самки.
Анализируя подобным образом рефлексологию животных, мы можем выработать целый арсенал теоретически обоснованных приемов, позволяющих добиваться садки самцов и у иных видов. Так например посадкой самки за веревку, привязанную к ноге, нам удавалось ускорить садку у индюков, павлинов, селезней. Основываясь на этом, мы применили ее к фазану (Ph. colchicus) и через несколько минут получали садку и у этой птицы, уже гораздо более дикой, чем домашние куры и индюки, у которых вероятно в результате отбора в обстановке пестроты и многопородности могла уже образоваться несколько большая неразборчивость и расплывчатость инстинкта — могли например выключиться из числа безусловных раздражителей окраска и другие внешние признаки. Однако выяснение решающих раздражителей представляет несомненно чрезвычайно благодарную и важную задачу. С какими интересными явлениями мы можем здесь встретиться, показывает например следующий случай в зоопарке Аскания Нова. Здесь в 1932 и 1933 гг. образовалась пара — первый шаг к спариванию — из двух>столь разнообразных птиц, как белощекая казарка и серый журавль. В 1932 г. журавль пал, и на следующий год та же казарка образовала неразлучную пару с другим журавлем. Ни внешний вид ни походка этих двух птиц не имеют ничего общего, тем не менее какой-то элемент в журавле оказался тем решающим раздражителем (возможно впрочем, что условным), который пустил в ход инстинкт казарки. Это тем более удивительно, что в том я?е парке имелись и другие белощекие казарки, с которыми эта могла встречаться и действительно встречалась. Гейнрот (1910, стр. 636) описывает аналогичный случай спаривания магелланской гусыни с павлином, причем гусыня гнала от себя самцов своего вида.
Экологические раздражители. Говоря об условиях, провоцирующих садку, нельзя упускать из виду и важного значения тех раздражений, которые идут со стороны окружающей среды. Здесь наше знание и чрезвычайно обширно и чрезвычайно недостаточно обработано аналитически. Для большинства видов довольно хорошо известны те условия, та обстановка, в которой проходит их естественное размножение, а значит и спаривание. Но какие именно из этих элементов являются решающими, детально исследовалось лишь по отношению к немногим видам. Несомненно, что ряд видов не размножается без наличия некоторых специфических условий. Так, птицы, гнездящиеся в дуплах, не начинают размножаться, если в их вольерах нет дупляков (или домиков), и устройство дупляков побуждает их к гнездованию, кладке яиц и спариванию. Также известно, что для ряда видов не удалось найти тех условий, которые заставили бы их гнездиться (казарка краснозобая и др.). Однако трудно сказать, какова здесь причинная связь — является ли тормозом инстинкта размножения отсутствие подходящих условий для гнездовании, или, наоборот, неправильное кормление и содержание не приводит физиологию организма в такое состояние, чтобы возникли те внутренние раздражители, без которых не могут действовать и внешние. На примере дупляков можно предполагать, что несмотря на наличие внутренних раздражителей не хватало внешних и размножение не вызывалось. Однако насколько можно эти отрывочные наблюдения обобщать— сказать трудно. Можно и должно лишь подчеркнуть, что аналитической экологии размножения диких животных еще не существует и она находится еще на элементарном описательно-наблюдательном уровне
[28].
Выводы и практические замечания. В настоящее время проблема получения спермы у самцов диких видов является одной из центральных проблем гибридизации, на пути разрешения которой мы встречаемся с рядом серьезных трудностей и пока в большинстве случаев терпим неудачи.
В некоторых случаях трудности получения спермы диких видов могут быть обойдены использованием для целей гибридизации диких самок под сперму от домашних видов. Например, имея возможность легко получать сперму домашнего петуха, селезня, гуся, можно ею осеменять чрезвычайно широкий круг диких видов. Однако здесь мы встречаемся с новым затруднением в виде процесса размножения самок; дикие самки кладут мало яиц, с трудом гнездятся, и иметь несущихся:, самок, годных для осеменения, еще труднее.
Таким образом наиболее удобным приемом должно явиться использование домашних самок под сперму диких самцов. Домашних самок можно иметь неограниченное количество; самки домашних видов птиц как правило дают гораздо больше яиц и с ними легко оперировать, самцов даже редких и ценных видов легче добыть хотя бы в одном экземпляре, что вообще говоря совершенно достаточно для искусственного осеменения большого количества самок. Все это вместе взятое и выдвигает на первое место задачу разработки методов для получения спермы у самцов диких видов (поскольку для самцов домашних видов эта проблема в основном уже разрешена).
К сожалению на сегодня вопрос о практическом методе получения спермы у живых диких самцов, имея в виду в первую очередь млекопитающих и птиц, остается еще неразрешенным. Основное затруднение, с которым мы здесь встречаемся, — это затруднение рефлексологического порядка. Основные задачи состоят в том, чтобы добиться садки, самца на самку своего вида или родственного домашнего вида и притом в такой обстановке, которая допускала бы немедленное взятие выброшенной им спермы. Это значит, что садка должна произойти в присутствии человека (открытом или скрытом) и притом в более или менее короткий срок после припуска самки к самцу.
К счастью в некоторых случаях это удается без особого труда, и тогда задача оказывается сразу решенной. Так например зубры, бизоны, бантенги довольно легко в таких условиях идут на домашних коров, мускусный селезень — на домашнюю утку и пр. Анализируя эти случаи, приходится подчеркнуть, что здесь мы имеем дело с домашними или полудомашними видами, во всяком случае с животными, достаточно прирученными. И это сразу подчеркивает решающую роль
приручения в этой проблеме. Можно думать поэтому, что именно на пути максимального приручения, приручения. к рукам, и лежит основной ключ к разрешению проблемы получения спермы. Пока самец дик, пуглив, получить от него садку в короткий срок после припуска самки достаточно трудно. Поэтому гибридизатор должен в совершенстве овладеть всеми теми приемами, которые выработаны практикой дрессировки и приручения, для того чтобы-иметь возможность в кратчайшие сроки приучать к себе диких животных. Такими приемами является прежде всего кормление из рук или в присутствии людей, постоянный уход и т. д.
План такого приручения может быть намечен в следующем виде.
ЭТАПЫ
0. Предоставление возможности самцам и самкам образовать пары (для моногамов).
1. Изоляция спаривавшихся самцов и самок решоткой.
2. Ежедневный повторный кратковременный припуск самки к самцу одним и тем же экспериментатором с предоставлением возможности вначале Свободных садок.
3. Приучение к присутствию экспериментатора при встречах с самкой и присадках.
4. Приучение к «ручной садке».
5. Приучение самок к спермособирателю перед пуском к самцу.
6. Получение садки на спермособиратель.
7. При желании получения естественного осеменения после 4-го (в некоторых случаях даже 3-го) этапа замена самки своего вида самкой другого вида.
Собирание спермы
Собирание спермы, эякулируемой при садке, составляет необходимый технический элемент этого способа получения спермы. В настоящее время эта сторона дела достаточно разработана применительно к млекопитающим и птицам (у рыб этих приемов не требуется; так как сперма получается простым выдавливанием ее в сосуд). Наиболее примитивным (оставляя старый губочный метод) является вагинальный способ, при котором после садки во влагалище вводится влагалищное зеркало специальной конструкции, по желобку которого вытекает наружу в подставленный сосуд сперма, разбавленная иногда в несколько раз влагалищными выделениями. Сперма благодаря этому получается загрязненная и при дальнейшем использовании для других самок может явиться источником передачи инфекций. Более совершенным является метод спермособирателя (рис. 31) — резинового чулка; вводимого во влагалище самки перед садкой. Здесь пенис самца не соприкасается со стенками влагалища, что предохраняет случаемых животных от взаимной инфекций, и сперма получается, совершенно чистой, без примеси влагалищных выделений. После садки спермособиратель осторожно извлекают, и сперма выливается в сосуд.
Наконец при садке на чучело или на самца (а в настоящее время с большим успехом и при садке на самку) употребляется искусственная вагина — резиновая трубка с двойными стенками, между которыми наливается теплая вода. Один конец трубки оставляется открытым для введения пениса, а в другой вставляется пробирка отверстием в сторону пениса. В момент садки находящийся рядом человек подставляет искусственную вагину так, чтобы пенис попал в ее отверстие, Ритмическим надавливанием на резиновую грушу ритмически изменяется давление между стенками вагины, что облегчает эякуляцию. Сперма стекает в пробирку, замыкающую вагину с другого конца; прилагаемый рисунок 31 изображает эти инструменты.
 Рис. 31. Инструменты для искусственного осеменения млекопитающих (фото Ландауэра).
А — искусственная вагина, В — влагалищное зеркало, С — спермособиратель, D — корнцанг, Е — ампула с разбавителем, F — шприц.
Рис. 31. Инструменты для искусственного осеменения млекопитающих (фото Ландауэра).
А — искусственная вагина, В — влагалищное зеркало, С — спермособиратель, D — корнцанг, Е — ампула с разбавителем, F — шприц.
Губчатый метод, ныне выходящий из употребления, состоял в том, что перед случкой во влагалище вводилась губка, которая пропитывалась эякулятом и по извлечении ее из вагины отжималась под особым прессом. Сперма при этом получалась тоже разбавленной влагалищными выделениями, и, кроме того масса сперматозоидов адсорбировалась на перекладинах губки и не отжималась обратно даже при сильном давлении.
1. Эта аппаратура применяется для сбора спермы у млекопитающих. Для птиц, естественно, аппаратура существенно иная. У куриных, у которых отсутствует пенис, а кроме того, как и у всех птиц, сперма эякулируется в ничтожном объеме (0,2–0,5 см
3), извлечение спермы наподобие вагинального, метода невозможно, а метод спермособирателя и искусственная вагина сливаются в единый метод.
Спермособиратель (рис, 32) устраивается следующим образом: рамка из проволоки с 4 ушками по углам обшивается резиной с загнутым нижним краем, образующим кармашек. Из резиновых шнурков де-дается шлейка из двух петель, скрепленных двумя пряжками
А и
В, благодаря чему образуются три. окна. I, II и III. В окно I продевается хвост, и пряжка
А передвигается назад, закрепляет нужный размер зияния. В окно II продеваются левое крыло и левая нога, в окно III — соответственно правое крыло и нога. Пряжка
В (отсутствует на рис.) продвигается вперед для натягивания резинки. Спермособиратель укрепляется так, чтобы он закрывал собой клоаку. Петух во время садки эякулирует сперму на смазанную вазелином резину спермособирателя, по которому сперма стекает в кармашек и оттуда выбирается пипеткой.
 Рис. 32. Спермособиратель куриный (по Соколовской).
Рис. 33. Спермособиратель гусиный (по Соколовской).
Рис. 32. Спермособиратель куриный (по Соколовской).
Рис. 33. Спермособиратель гусиный (по Соколовской).
Для водоплавающей птицы, имеющей пенис, форму спермособирателя приходится изменять. Соколовской сконструирован спермособиратель (рис. 33) в виде алюминиевой пластинки, к которой пришита плотная резина, оттопыривающаяся от пластинки, так что между ней и пластинкой образуется воронка. Внизу воронки — алюминиевый стаканчик. Такой спермособиратель удобно делается из алюминиевой конусовидной пробирки от центрифуги. У пробирки сделан разрез вдоль одной стенки с небольшим поперечным разрезом внизу. Края пробирки от разреза развернуты и образуют пластинку, о которой сказано выше.
Однако приручение самцов птиц приводит к возможности значительного упрощения работы. Так, токующему индюку можно просто подставить часовое стекло или стаканчик, куда он выбрызгивает сперму. Также на стаканчик приучил давать сперму павлина Тиняков. Кузьмина получала сперму селезня просто в чашку Петри, а затем в стаканчик. Лучшие результаты получались, если брался стаканчик, обтянутый сверху резиной с отверстием. Селезень, вводя пенис в отверстие резины, долго удерживался в таком положении, почему вся сперма успевала стечь и ее получалось больше, чем при других способах. Эти стаканчики являются уже скорее искусственными вагинами, чем спермособирателями, так как при работе с ними контакта между клоаками самца и самки уже не допускается, и самец может сидеть на самке или на чучеле в любом положении, а например индюк может, эякулируя, просто стоять рядом с самкой, положенной на землю. Наконец Тиняков получал большие порции спермы, приклеивая петуху к клоаке неразвернутый презерватив и повторно пуская петуха в садку.
Введение спермы в самку
Введение спермы может быть естественным или искусственным. В первом случае мы поручаем это самому самцу путем нормального полового акта (рис. 34), во втором случае делаем это при помощи инструментов, вводя сперму, предварительно собранную в сосуд.
 Рис. 34. Садка зубробизона на корову (фото Аскании-Нова).
Рис. 34. Садка зубробизона на корову (фото Аскании-Нова).
До разработки метода искусственного осеменения естественный метод был единственным, и почти все гибриды, полученные до последнего времени в природе и в зоосадах (кроме старых асканийских зеброидов и гибридов в СССР наших дней), получались естественным методом введения спермы, совпадающим с ее получением.
В настоящее время разработка метода искусственного осеменения переносит основные перспективы именно на этот метод. Тем не менее естественный метод может сохранить свое применение в некоторых случаях. Так например, если птицы не размножаются, будучи изолированными от самцов, к ним приходится подпускать самцов другого рода; для возбуждения инстинкта гнездования, иначе мы не получим гибридов, сколько бы мы ни осеменяли такую самку искусственно. Или если самка принадлежит к трудно приручаемому виду, например антилопы, и чрезвычайно пуглива, попытки искусственного осеменения могут привести к тяжелой травматизации и даже гибели животного. В этих случаях подпуск самца близкого вида может легче привести к получению гибридного приплода. Этот естественный метод гибридизации сталкивается с часто трудно преодолимыми препятствиями, о которых мы упоминали в главе III и при анализе проблемы получения спермы и которые в данном случае распадаются на два вида:
1) рефлексологические препятствия,
2) механические препятствия.
Рефлексологические препятствия для гибридного спаривания сводятся к тому, что самка чужого вида не возбуждает самца вовсе. или в достаточной степени не реагирует на половые сигналы, идущие от. самца. Об этом мы говорили выше.
Эти препятствия однако во многих случаях устраняются или отпадают, в результате чего мы и имеем многочисленные, хотя относительно и редкие, случаи гибридизации естественной или в зоосадах и хозяйствах. Но так как базироваться в работе на этих редких исключениях невозможно, то следует выяснить методы и пути, которые могут привести к систематическому преодолению этих препятствий. Таких методов может быть много. Некоторые из этих приемов имеют довольно общее значение, хотя здесь и нужно подчеркнуть, что в каждом гибридизационном сочетании видов эти препятствия специфичны и подлежат конкретному выяснению.
1. Совместного содержания и воспитания во многих случаях оказывается уже достаточно для получения гетерогенного спаривания. Многие виды например птиц при этом спариваются в весьма широких пределах. Например селезень мускусной утки (Cairina moschata), уединенный с домашними утками или гусынями, очень легко переходит в наступления, а так как он является сильным самцом, хватающим самок, стаскивающим их за крыло в воду и т. д., то самки тех видов, где самец имеет сходные инстинкты, реагируют на его ухаживания. Точно, так же самцы разных быков, баранов, козлов и т. д. идут довольно легко на самок других видов. Совместное воспитание в Аскании-Нова (Шулер) отнятых от матерей телят антилоп, канн, бантенга и зебу привело к попыткам спаривания самца канны с самкой бантенг, как только у нее наступила первая течка. Совместное раннее воспитание, говорят, облегчает гибридизацию рыбок-живородок.
2. Помощь самцу в том случае, если самка неохотно подпускает чужого самца, может быть оказана разными способами. Заключение в тесный выгон, привязывание (рис. 34), завязывание головы, чтобы самка не видела самца, подвязывание ног, чтобы она не могла ударить самца, и тому подобные приемы могут помочь делу. Впрочем подрезание ножек и крыльев не облегчало спаривание видов дрозофил в опытах Стертеванта.
3. Метод подмены самки. На базе установления условных рефлексов у самца на обстановку ручной садки может быть развит прием подмены самки, примененный нами и Соколовской при гибридизации гуся с уткой. Гусак «Дружок» был хорошо приучен к ручной садке, при которой ему приносили гусыню. Экспериментатор деру жал ее в руках, клал ее на землю и сажал на гусыню гусака. После месяца такой работы ему была принесена белая пекинская утка.[Гусыня находилась. в соседнем выгуле, отделенном сеткой. На зов экспериментатора гусак начал проявлять признаки возбуждения, стал гоготать, но направился не к утке, а к сетке навстречу гусыне, проявлявшей тоже большое возбуждение. Итти к утке гусак отказывался. Тогда гусыня была поймана, принесена в отделение гусака и положена рядом с уткой.
Когда гусак направился к гусыне и схватил ее за голову, была произведена подмена, гусь оказался посаженным на утку и правильно покрыл ее. В следующие дни достаточно было экспериментатору держать на земле утку, а помощнику принести кричащую гусыню и стоять с ней, держа ее вблизи, чтобы гусак уже без отказа садился на утку. (рис. 35). Замечательно, что к этому приему приходилось прибегать даже для того, чтобы посадить гусака на другую гусыню — пока не принесут «его» гусыню, гусак возбужденно стремился к ней через сетку и на зов экспериментатора не шел. Принос «его» гусыни успокаивал одну часть его возбуждения, и гусак уже нормально спаривался с подменяющей самкой.
 Рис. 35. Подстановка утки гусю (фото Аскании-Нова).
Рис. 35. Подстановка утки гусю (фото Аскании-Нова).
Таким же приемом мы заставляли прирученного петуха естественно осеменять индюшку.
4. Большое значение имеет поднятие возбуждения самца, для чего могут быть использованы разные приемы — нажимы как со стороны внутренних стимулов, так и со стороны внешних. К таким внутренним стимулам могут быть отнесены например введение пролана, гравидана и других эндокринов, усиленная фосфористая диета, моцион и пр. К внешним — содержание в виду самок.
Мы упоминали уже о практиковавшемся приеме раскраски кобыл и ослиц (Hunter; см. Дарвин, «Происхождение человека» (белыми полосками или одевания полосатых шкур и попон при случке с зебрами (Wohltmann, 1911). Петух, не идущий нормально на цесарку, идет на нее, если ему бросить ее внезапно с некоторой высоты.
Подробные указания об искусственном возбуждении самцов для спаривания их с самками другого вида дает для бабочек Штандфусс (1896). Его указания частично можно распространить и на другие объекты.
В ряде случаев прямые попытки естественного осеменения бабочек чужими самцами, помещенными в общий садок, не приводили к цели. Самцы либо вовсе не возбуждались либо возбуждались слабо. Для того чтобы поднять возбуждение самцов вида
а и заставить их итти на самок вида
b, Штандфусс помещал в соседний, садок самок вида
а. Самки вида
а выделяли запах, приводящий самца их вида в сильное возбуждение. Но так как свои самки оказывались недоступными, то самцы бросались на находящихся с ними самок чужого вида. Этот прием был разработан и удачно привел к получению первых экспериментальных гибридов бабочек в 1837 г. Хузом (Huose, 1842, Smerinthus populi × ocellatus).
Несколько иначе этот метод применялся в другом случае. В один и тот же садок к самцам. вида
а сажались самки двух видов: и
а и b. Но самцов было значительно больше, чем самок их вида. Придя в возбуждение от запаха своих самок, самцы бросались на них, но так как для всех самцов «своих» самок нехватало, то самцы начинали спариваться и с чужими самками.
Аналогичное возбуждение может конечно применяться и в других видах. Однако этот метод требует понимания экспериментатором тех основных возбуждений, которых нехватает самцам. В одних случаях это может быть запах, как в опытах Штандфусса, в других — зрительные образы и пр. Если это запах, то возможно применить обмазывание чужих самок пахучими выделениями «своих» самок — мочой и потом, выделениями при течке и пр. Если нехватает зрительных, образов, необходимо дополнить их раскраской самок и пр.
Тем не менее рефлексологические препятствия к гетерогенному спариванию могут оказаться чрезвычайно большими и непреодолимыми по той причине, что мы этой областью овладели еще не достаточно, и если самец не выказывает желания итти к самке чужого вида, то экспериментатору часто чрезвычайно трудно решить, чего именно нехватает самцу для возбуждения. В птичнике Аскании-Нова например имелся самец аргус в полном здоровье и красоте. Мы пускали к нему индюшку, цесарку, фазанку, применяли метод посадки самки перед ним при помощи веревки, привязанной к ноге. Если бы самец обнаружил хотя слабую попытку к спариванию, можно было бы, ухватясь за нее, развить возбуждение больше. Однако ни в одном случае аргус не обнаружил ни малейшей половой реакции на самок и продолжал шагать вдоль сетки вольеры, оставляя экспериментатора в полном неведении того, в какой области лежит та пружина, на которую необходимо нажать — в области ли внешнего вида самки, ее голоса, в области ли окружающей обстановки и т. п. Конечно здесь еще в большей степени, чем в случае гомогенного спаривания, необходимы изучение естественной экологии вида и ее детальный экспериментальный анализ.
Далее немаловажными могут быть и
механические препятствия к гетерогенному половому акту, о чем мы уже упоминали раньше. В некоторых случаях они могут оказаться непреодолимыми, если например размер полового аппарата самки слишком мал, или, наоборот, самец слишком мал, чтобы суметь овладеть самкой, или если, как у насекомых, твердые хитиновые части полового аппарата, весьма специфические у разных видов, окажутся несоответствующими друг другу, и т. д.
Однако в ряде случаев умелым вмешательством гибридизатор может устранить эти препятствия или ослабить их. Так, если мы скрещиваем дикий вид с домашними формами, то у нас всегда есть выбор пород различных величин. Для гибридизации с фазанами например удобно брать кур мелких пород или бентамок, для гибридизации с дикими утками — некоторые мелкие породы домашних. При получении мулов выбирают для скрещивания с ослом невысоких кобыл, а под жеребцов — наоборот, самых рослых ослиц.
Если этого недостаточно, то можно прибегнуть к еще большим уловкам — например поставить кобылу, идущую под осла, в ямку или под горку и т. п. Необходимо только путем тщательного наблюдения установить природу механического препятствия. Так например особая форма наследственно куцых кур дает очень часто, неоплодотворенные яйца. Наблюдение показывает, что благодаря отсутствию рулевых перьев верхние кроющие хвоста свисают вниз и закрывают отверстие клоаки. Простое постригание их открывает клоаку и позволяет петуху оплодотворить курицу. Точно так же у курдючных овец их тяжелые висячие курдюки сильна мешают спариванию, и чабаны систематически помогают баранам других пород, отводя хвост овец в сторону.
Однако такого порядка помощь возможна лишь для случая с самцами, дающими садку в присутствии человека, и в этих случаях надобность в ней в значительной степени отпадает, так как мы получаем возможность применить искусственное осеменение, снимающее сразу все механические препятствия.
Искусственное введение спермы сейчас находится уже в значительной мере в наших руках и требует лишь дальнейшего усовершенствования. Собранную сперму можно вводить двумя основными путями: 1) через вагину, естественное половое отверстие или 2) непосредственно в полость тела, к яичнику через стенку брюшной полости. Первый способ уже достаточно разработан и с успехом применяется у млекопитающих, птиц и пчел. Попытки интраперитонеального, «брюшного» осеменения пока дали в одном случае результат (у кролика). Возможно однако, что в дальнейшем этот метод получит развитие, особенно в тех случаях, когда сперматозоиды окажутся неспособными совершить нормальный путь в чужих половых путях от вагины до встречи с яйцом.
У млекопитающих собранная сперма при помощи шприца вводится в шейку матки самки, находящейся в охоте. У большинства млекопитающих этого достаточно для осеменения. Недостаточно этого лишь у животных с овуляцией, провоцируемой коитусом (кролик). У таких видов после введения спермы необходим естественный коитус с вазэктомированным самцом, который должен быть предварительно приготовлен недели за две (или инъекция пролана).
У птиц сперма вводится в отверстие яйцевода (рис. 36 и 37), в вагину, открывающуюся в клоаку, которую приходится несколько растянуть каким-либо инструментом, например но со расширителем или просто пальцами.
Искусственного введения спермы у насекомых (кроме пчел) в целях гибридизации еще не производилось, почему описывать подробно довольно кропотливую методику введения спермы пчелиным маткам мы здесь не будем — принципиальных отличий она не имеет и особенности связаны лишь с необходимостью работать под бинокулярной лупой. Музалевский и Лысенко дали
более простой способ (Музалевский, и Козлов 1933).
 Рис. 36. Сбор спермы петуха пипеткой (по Никитиной).
Рис. 36. Сбор спермы петуха пипеткой (по Никитиной).
Разбавление спермы
Тесным образом с проблемой введения спермы связан вопрос
о разбавлении и хранении спермы. Необходимость разбавления спермы часто диктуется двумя соображениями.
 Рис. 37. Впрыскивание спермы курице (по Никитиной).
Рис. 37. Впрыскивание спермы курице (по Никитиной).
1. Получение спермы у самцов обычно сопряжено со значительной затратой сил и времени, почему возникает настойчивая потребность полученную сперму использовать для осеменения максимально возможного количества самок. Это сокращает затрату труда и времени на осеменение одной самки и позволяет вести работу в более широком масштабе, что для многих задач гибридизации чрезвычайно важно.
2. Во многих случаях сперма получается в столь ничтожном количестве, что с ней технически трудно манипулировать. Так, птицы дают вообще эякулят, не превышающий 0,5–1,0 см
3. При добывании спермы хирургическим путем из эпидидимиса количество спермы (масса сперматозоидов) может быть еще меньше. Наконец при гибридизации с редкими видами в распоряжении гибридизатора может находиться всего один самец, каждым эякулятом которого приходится крайне дорожить.
Проблема разбавления спермы в настоящее время в значительной степени разрешена, в особенности работами Милованова и др., и быстро совершенствуется. Выяснено в первую очередь для млекопитающих, что изотонические растворы фосфатов, сульфатов и тартратов в смеси с изотоническим раствором глюкозы являются средой, при помощи которой можно во много раз разбавить сперму либо вовсе без вреда для ее оплодотворяющей способности либо лишь со слабым ее понижением.
Так как у каждого вида свое осмотическое давление спермы, которое иногда довольно заметно отличается от других видов, то для получения наилучших результатов для каждого вида должен быть подобран специфический разбавитель.
Приводимая таблица показывает состав разбавителей для домашних млекопитающих.
I. Глюкозо-фосфатные (в двух растворах)
Для барана (ГФО-2)
1. Глюкоза безводная — 64,0
— Вода дестилированная — 1000,0
2. Натрий фосфорнокислый однозамещ. — 41,0
— Калий фосфорнокислый однозамещ. — 1,7
— Кальций фосфорнокислый двузамещ. — 0,1
— Магний фосфорнокислый двузамещ. — 0,1
— Вода дестилированная — 1 000,0
Для быка (ГФК-3)
1. Глюкоза безводная — 57,0
— Вода дестилированная — 1000,0
2. Натрий фосфорнокислый двузамещ. — 36,7
— Калий фосфорнокислый однозамещ. — 1,4
— Натрий сернокислый безводный — 1,7
— Кальций фосфорнокислый двузамещ. — 0,1
— Магний фосфорнокислый двузамещ. — 0,1
— Вода дестилированная — 100,0
II. Глюкозо-сульфатные (в одном растворе)
Для жеребца (СГЛ-2)
Глюкоза безводная — 57,6
Натрий сернокислый безводный — 3,4
Пептон (без солей) — 2,0
Вода дестилированная — 1000,0
Для быка (СГК- 2)
Глюкоза безводная — 12,0
Натрий сернокислый безводный — 13,6
Пептон (без солей) — 5,0
Вода дестилированная — 1000,0
Оба раствора хранятся отдельно и перед употреблением смешиваются в отношении 1:1.
Для птиц разбавителем могут быть жидкие фракции яичного белка, отсасываемые пипеткой
[29].
Хранение спермы
Проблема хранения спермы представляет для гибридизации огромное значение, почему разрешению этой проблемы, еще далекой от разрешения, должно быть уделено самое серьезное внимание. Значение этой проблемы обусловлено следующим.
1. Даже небольшой эякулят спермы содержит в нормальных случаях огромное количество сперматозоидов, число которых в 1 см
3 достигает сотен миллионов (у утки до
1/
2 млрд.). Поэтому одного такого эякулята при возможности его хранения могло бы хватить для большого количества самок, даже полагая на каждое осеменение хотя бы по 10 000 000 сперматозоидов против одного необходимого для оплодотворения. Длительное храпение спермы дало бы возможность наиболее рационального и полного использования эякулятов. Между тем благодаря нашему неумению хранить сперму многие эякуляты получают ничтожное использование, так как оказывается невозможным иметь одновременно достаточно большое количество самок, годных для осеменения по своему физиологическому состоянию (наличию течки и т. п.).
2. Гибридные сочетания многих видов невозможны или затруднены вследствие полного или частичного несовпадения фаз половых циклов., Так, у большинства диких баранов течка падает на зиму, когда у домашних овец она уже окончена. У лисиц сезон спермообразования оканчивается тогда, когда у собак течка лишь начинается. Лебеди гнездятся с начала марта, утки — в апреле, крохали — даже в конце мая или в июне, и т. п. Возможность соответственно долгого хранения спермы сняла бы и эти затруднения.
Сформировавшиеся зрелые сперматозоиды начинают энергично двигаться. После эякуляции в связи со смещением их с секретами добавочных желез полового аппарата, имеющими щелочную реакцию, это движение приобретает необыкновенно бурный характер, и густая сперма под микроскопом производит впечатление «кипения». Это бурное движение продолжается, постепенно ослабевая, от нескольких часов до нескольких суток, и наконец сперматозоиды замирают, теряя вообще говоря свою оплодотворяющую способность. Так как на движение сперматозоиды расходуют энергию, то проблема хранения их может разрешаться двумя путями:
1) прекращением или ослаблением расхода энергии,
2) пополнением запасов энергии в сперматозоиде.
Оба эти пути в настоящее время начинают разрабатываться с некоторыми успехами. Здесь же следует отметить, что проблема сохранения спермы связана и с еще одним условием, а именно — с борьбой с микрофлорой спермы. Эякуляты всегда оказываются загрязненными флорой мочеиспускательного канала, клоаки и пр. и, представляя отличную питательную среду, быстро загнивают. Более или менее снимается эта проблема лишь в том случае, если сперма берется из эпидидимиса или семяпровода стерильно.
Разрешение проблемы длительного хранения спермы должно итти комбинированным путем по всем трем указанным направлениям и должно предусмотреть по Милованову следующее:
1) добиться возможно большего замедления расхода энергии сперматозоидами, снизить их активность однако лишь на время хранения;
2) найти приемы питания сперматозоидов;
3) найти среду, наиболее благоприятную для сперматозоидов;
4) предупредить изменение свойств этой среды, в особенности в результате жизнедеятельности сперматозоидов (интоксикацию),
5) предупредить развитие микробов и плесени.
Нужно иметь в виду, что продолжительность жизни сперматозоидов сильно зависит от степени их зрелости. Как молодые недозрелые сперматозоиды, эякулируемые в результате часто следующих друг за другом садок (до 20 садок в день в опытах Ллойд-Джонса и Хея с кроликом), так и старые сперматозоиды, долго находящиеся в самце, погибают скорее, чем находящиеся в оптимально зрелом состоянии.
Все предложенные методы хранения можно вслед за Миловановым разбить на следующие категории:
1) методы, основанные на понижении температуры;
2) методы, основанные на ограничении газообмена;
3) методы, основанные на применении консервирующих веществ;
4) методы, основанные на применении питательных сред.
Понижением температуры возможно значительное ослабление деятельности сперматозоидов вплоть до остановки их движения, причем даже сперматозоиды теплокровных выдерживают охлаждение ниже нуля, лишь постепенно теряя способность к возобновлению движения. Кржишковский и Павлов получали оживление, однако не далее 95 часов, при охлаждении в капилярах, предохраняющих от образования льда. Хэммонд (Hammond, 1930) показал на кролике, что для охлаждения существует некоторый оптимум, и наиболее долго — и при этом значительно дольше, чем при температуре тела, — сперматозоиды живут при 10°.
Температура хранения — Наибольший срок сохранения сперматозоидами способности к оплодотворению (в часах)
35° — 14
10° — 96
0° — 15
Ослабление газообмена путем прекращения доступа кислорода способствует более длительному переживанию. Однако Очи (Ochi) нашел, что оптимум лежит не при полном отсутствии кислорода, а при 10 % кислорода. Без кислорода и при 30 % и выше переживание сокращается почти вдвое. Кржишковский и Павлов сохраняли сперматозоиды в капилярах, закрытых с обоих концов, а Хэммонд и др. предложили сохранять сперму под слоем парафинового масла.
Из консервирующих средств большое значение играет углекислота, — наркотизирующая сперматозоиды. Для аэробных сперматозоидов морских ежей хорошие результаты дают слабые дозы цианистого натрия, прекращающего движение, позже восстанавливающееся.
Наконец предоставление сперматозоидам глюкозы, предложенное Ямане и Като, обеспечивает их питание, несомненно возможное несмотря на отрицание его некоторыми авторами.
Так как в методы разрешения проблемы хранения спермы задачи гибридизации не вносят чего-либо нового, то подробнее на этом важном разделе мы останавливаться не будем, отсылая к имеющимся на русском языке изданиям ЛИО ВИЖ и позднейшим работам («Проблемы животноводства»), тем более что пока успехи в этой области еще скромны и не превышают немногих дней. Нам необходимо было лишь подчеркнуть, какие элементы в этой главе гибридагогии должны быть разработаны более, чем они разработаны по отношению к задачам работы лишь с домашними животными.
В качестве такой проблемы, оставшейся пока мало разработанной и специфически важной для гибридагогии, являются проблема жизни и движения сперматозоида в половом тракте самки чужого вида и проблема помощи сперматозоиду в его скорейшем и безболезненном достижении яйца в этих чуждых для него условиях.
Проблема достижения сперматозоидом яйца в половых путях самок чужого вида и помощь ему в этом
Сперматозоиды, попадая в половой тракт самки чужого вида, оказываются вообще говоря в обстановке, для них. необычной. Каждый вид животных помимо морфологических особенностей имеет конечно и свою собственную физико-химическую характеристику. Указания на это мы получаем хотя бы при сравнительном изучении осмотического давления спермы разных видов животных. Следующие таблицы показывают степень этого различия как в абсолютных величинах, так и в процентах, если взять за 100 сперму человека с наименьшим осмотическим давлением из числа видов, приведенных в таблице.

Уже в пределах этого списка различие достигает солидной цифры в 20 %. Между тем рядом исследований показано, что сперматозоиды могут быть весьма чувствительны к колебаниям осмотического давления. Такими стеносмотическими формами являются млекопитающие и птицы (?) в противоположность формам, у которых оплодотворение происходит в воде (рыбы, амфибии, иглокожие), сперматозоиды которых способны переносить более сильные колебания.
Изменение осмотического давления на 50 % (особенно в сторону повышения) способно убить сперматозоиды млекопитающих почти моментально. Если вспомнить по этому поводу, что для прохождения полового тракта самки сперматозоиду требуется несколько часов, то станет понятным, что даже незначительные отклонения в осмотическом давлении могут явиться сильнейшим препятствием для гибридизации.
Прилагаемая таблица показывает, как резко снижается переживание сперматозоидов барана при незначительных отклонениях от оптимального осмотического давления.

Если признать, что вообще говоря осмотическая характеристика сперматозоида соответствует условиям в женском половом аппарате, то легко видеть из этой таблицы и рис. 38, что уже в половых путях коровы сперма барана будет себя чувствовать (в отношении осмотического давления) в два-три раза хуже, чем в половых путях овцы (переживание вместо 3,4 лишь 1,5).
Осмотическое давление мы взяли лишь в качестве примера, правда, одного из самых важных факторов. Но помимо того и другие физические, химические и биологические (например фагоциты) факторы в чужих родовых путях могут оказаться губительными для сперматозоидов.
 Рис. 38. Кривая падения жизнеспособности спермы при изменении осмотического давления (по Милованову).
Рис. 38. Кривая падения жизнеспособности спермы при изменении осмотического давления (по Милованову).
Прямые наблюдения Дональда Иочема (Yochem, 1927) показали резкое снижение срока жизни сперматозоидов морской свинки в шейке матки крысы и обратно по сравнению со сроком переживания в шейке матки своего вида (при одинаковой технике искусственного осеменения).
Продолжительность жизни в часах

При гетерогенном осеменении переживание снижалось таким образом на 75 %.
Более удивительные результаты были получены Соколовской и нами у птиц. Ненесущимся кряквам была искусственно введена сперма: одной — селезня, другой — гусака, третьей — петуха. Вскрытие через 22–25 часов показало, что сперматозоиды в большом количестве проникли в самые верхние отделы яйцевода. Сперматозоиды селезня сохраняли оживленное движение; сперматозоиды гуся (22 час.) и петуха (25 час.) в большинстве были неподвижны, но некоторые имели хорошее поступательное движение.
Широкое проведение подобных опытов на разных сочетаниях должно будет нарисовать конкретную картину того, с какой степенью препятствий мы встречаемся в этой области. Если вернуться к осмотическому давлению, то возможно конечно, что у некоторых близких видов оно может оказаться более различным, чем у далеких видов. Так, и в приведенной на стр. 125 таблицы видно, что жеребец в этом отношении стоит очень близко к человеку и собаке — видам систематически очень далеким, и, наоборот, далеко стоит от гораздо более родственного ему барана. Мало того разбавитель для спермы лошади оказывается подходящим для спермы гуся (если судить по движению сперматозоидов).
В настоящее время мы еще слишком мало знакомы с этой областью, чтобы извлечь из нее достаточно конкретные указания для гибридизации. Однако некоторые указания для нас достаточно важны.
Так например Гельхорн для лягушки и Поярков для собаки нашли, что сперматозоиды их выдерживают гораздо лучше гипотонию, чем гипертонию. Если поэтому мы имеем два вида — гипотонический и гипертонический (по сравнению друг с другом), то осеменение гипотонического вида гипертоническим должно будет дать ceteris paribus лучшие результаты, чем реципрокное осеменение. С этой точки зрения например осеменение коровы спермой барана (в осмотическом отношении) более перспективно, чем осеменение овцы спермой быка.
В качестве более сложного метода помощи сперматозоидам может быть поставлен вопрос о попытках изменения осмотического давления в родовых путях путем введения в них соответствующих веществ. Для этого могут быть попользованы введения этих веществ сверх-него конца яйцевода через брюшную полость. Правда, сильные буферные способности организмов почти не позволяют считать эти перспективы реальными, однако упомянуть их следует. Эти приемы будут аналогичны приемам помощи гетерогенному оплодотворению, и поэтому мы остановимся на них в следующей главе.
Более перспективным путем мы считаем путь брюшного (интраперитонеального) осеменения, имеющего целью внести сперматозоиды сразу в непосредственную близость к яичнику и верхним отделам яйцевода. Так как яичник у большинства млекопитающих (кроме хищных и некоторых других) лежит свободно в полости тела, то, проникнув в эту полость шприцем, мы можем вылить сперму непосредственно к яйцам. Опыты подобного рода были поставлены с кроликами, и в одном случае дали положительные результаты.
Еще с 1880 г. Pinner инъицировал тушь в полость тела около верхних частей труб кролика, и уже через 2
1/
2 часа при вскрытии тушь была обнаружена в матке и во влагалище. Лоде (Lode) повторил опыт на 2 кроликах с яйцами Ascaris и нашел их в средней части труб через 20–36 часов после инъекции в полость тела.
Для разработки этого метода чрезвычайно важно изучить действие жидкостей тела на сперматозоиды, в частности брюшной жидкости. Мы произвели опыт прибавлением жидкости из брюшной полости только что убитой овцы к сперме барана и козла. Небольшая прибавка (1:1) при кратком наблюдении не оказала заметного действия на движение сперматозоидов обоих видов. Наоборот, прибавление брюшной жидкости примерно в 8—10-кратном размере убило оба вида сперматозоидов моментально. Эти наблюдения однако слишком отрывочны, чтобы не требовать повторения.
Брюшное осеменение у птиц затруднено расположением мезентериальных листков. Воронка яйцевода поднимается на яичник с вентрально-латеральной стороны, и устье его отгорожено от брюшной стенки тонкой складкой брюшины. Поэтому шприц, вводимый через стенку тела, даже минуя omentum, попадает концом в полость между этой складкой брюшины и париетальным листком, и сперма остается в этой полости, не попадая к яичнику. Проколоть и эту тонкую пленку трудно, так как она слишком податлива.
Поэтому нами была предложена следующая операция (на курах): между VI и VII ребрами выше уровня process uncinatus(на 2
1/
2 пальца от средней линии спины влево и на 2
1/
2 пальца кпереди от начала легко прощупываемого бедра) делается разрез около 1
1/
2 см в коже и стенке тела. Щипцами или сильно отсасывающей пипеткой, введенной в разрез, притягивается упомянутая складка брюшины, вытягивается в разрез, и в ней между кровеносными сосудами скалпелем прорезается несколько отверстий. После этого брюшина вправляется и рана зашивается. Операция переносится очень легко. Впрочем поставленные опыты осеменения по этому способу положительных результатов еще не дали, хотя сперма попадала на яичник и в устье яйцевода.
Обсуждая вопрос об отношении сперматозоидов к жидкостям самок другого вида, уместно кратко упомянуть о вопросе об естественных гетероспермотоксинах, т. е. об убивающих сперматозоиды токсинах из сывороток крови чуждого вида. По этому вопросу имеется ряд очень противоречивых опытов. В одних случаях сперма погибала в гетеросыворотке очень быстро, в других случаях жила долго. Так как сперма легко погибает от разных причин, то случаи длительной жизни более показательны. Так, в опытах Шороховой (1933) сперматозоиды человека жили в сыворотке морской свинки (другой отряд!) 36 часов, так что говорить о токсичности нельзя, хотя в изосыворотке жизнеспособность сохранялась еще более. Интересно, что свежая сыворотка содержит какие-то, вещества, подавляющие подвижность спермы, а при инактивации сыворотки эти вещества исчезают. В инактивированной сыворотке морской свинки сперматозоиды человека жили значительно лучше, чем в нормальной женской сыворотке (до 92 часов). Шорохова считает, что эти термолабильные вещества являются не токсинами, а ферментами крови. Несомненно, что-дальнейшее изучение этого вопроса очень важно как для проблемы гетерогенного осеменения, так и оплодотворения.
Проблема своевременности гибридного осеменения
Проблема своевременности гибридного осеменения должна быть подчеркнута в заключение, так как по этой линии, как упомянуто, перед гибридизацией стоят свои препятствия, подлежащие устранению на базе их изучения.
В общей форме можно сказать, что сперма должна быть введена с таким расчетом, чтобы к моменту появления яйца в трубе (или в верхнем отделе яйцевода) сперматозоиды уже находились
в этом месте, причем, учитывая сказанное выше, важно, чтобы им пришлось как можно меньше дожидаться яйца.
Это возможно без труда в том случае, если периоды размножения и половой охоты у видов, вводимых в скрещивание, совпадают. Этого однако может и не быть. Гибридизатору приходится оперировать с животными, привезенными из разных климатов, от полярного до тропического, и даже из разных полушарий, южного и северного, причем половые циклы могут расходиться до шести месяцев.
Преодолеть эти препятствия можно двумя путями. Первый, о котором мы упоминали, наиболее радикальный, — это путь длительного хранения спермы. Однако хранение спермы в течение 6 месяцев пока не осуществимо. Между тем, если сперма должна быть взята у вида, который дает ее позднее, чем приходят в охоту самки второго вида (например самец тар и самка овца), то возникает задача хранить сперму уже не шесть месяцев., а 10–11, т. е. в 50—100 раз дольше, чем это возможно в настоящее время.
Поэтому в настоящее время внимание должно быть уделено второму методу — методу передвигания половых циклов. Искать разрешение этой проблемы возможно различными методами, в частности
1) по линии изменения экологии вида и 2) по линии прямого физиологического-влияния на процесс овуляции и спермогенеза.
В качестве первого метода приведем пример передвигания полового цикла птиц, привезенных из южного полушария в северное. В настоящее время в северном полушарии разводится уже значительное количество видов, передвинувших свою фенологию на 6 месяцев, — таковы например мускусная утка, Магелланов гусь, чилийская свиязь и др. Некоторые виды однако совершают этот переход с большими трудностями и долго продолжают сохранять календарь южного полушария.
Какими приемами можно добиться этого передвигания, показывает пример работы в СССР со страусами-эму.
В 1923 и 1924 гг. работая, в Московском зоопарке, Фортунатов попытался передвинуть ближе к весне кладку имевшихся там нескольких пар эму. Нормально последние несли свои великолепные чернозеленые яйца в декабре, и за редкими исключениями все яйца оказывались неоплодотворенными. Исходя из предположения, что содержание на воздухе и в холоде задержит кладку, он пристроил просторный открытый загон, где птица содержалась до наступления больших холодов. По переводе в помещение температура держалась и нем от 0 до 5°. В результате кладка передвинулась на конец января.
В следующем году таким же путем удалось передвинуть кладку на февраль, а в 1925 г. кладка сдвинулась еще дальше к весне, и удалось получить довольно удачный вывод птенцов.
Большое значение в этом направлении имеет для нас свойство птиц продолжать кладку и задерживать переход к насиживанию в случае потери первой кладки. Гейнрот в работе о каролинской утке (Lampronessa sponsa) показал, что если отнять у птицы первую кладку, то это не только заставляет птицу через некоторое время приступить ко вторичной кладке, но оказывает на всю ее физиологию столь сильное воздействие, что у нее передвигается (отодвигается) на месяц время наступления линьки. Поскольку линька, как в настоящее время хорошо выяснено, определяется гормонными воздействиями эндокринных желез, антагонизирующих с половой железой и имеющих свою цикличность, причинно увязанную с работой половой железы, постольку в данном случае воздействием на инстинктивный аппарат птицы мы оказываемся в состоянии повлиять на работу всей ее эндокринной системы, передвинув ее циклические явления на некоторый срок. Было бы чрезвычайно интересно выяснить, как далеко простирается это воздействие, оказывает ли влияние это отставание линьки на месяц на начало яйцекладки следующей весной и какой силы должен быть нажим на нервную систему, чтобы это влияние осуществить.
Прямое физиологическое воздействие и вызывание овуляции недавно начаты разработкой и обещают интересные перспективы. В наших руках находится уже целый ряд чистых гормонов — тиреоидин, адреналин, пролан, фолликулин, ряд других животных препаратов — гравидан, сушеная плацента, свежие железы и пр.; использование их в указанных целях крайне интересно. На млекопитающих проблема искусственного возбуждения охоты несомненно близка к разрешению. Правда, выяснено, что течка представляет сложное явление: необходимо получить овуляцию, внешние проявления охоты; необходимо подготовить матку к тому, чтобы яйцо могло закрепиться в ней, и т. д. Отдельные моменты — например вызывание овуляции, вызывание внешних признаков охоты — в основном уже разрешены, и остается лишь привести их в более стройный вид.
Однако эти опыты (М. и Б. Завадовские и др.) показали, что каждое животное имеет свою, весьма резко выраженную специфику. Отсюда возникает обязанность овладения этим методом и разработки его применительно и к разнообразным видам диких животных, входящих в гибридизационные ресурсы.
Глава VI. Проблема гетерогенного оплодотворения
Проблема овладения гетерогенным осеменением, как мы видели, в основном разрешается приемами искусственного осеменения. Эти достижения снимают большой ряд трудностей, стоящих перед гибридизатором, большие количества видов переводят в круг возможных ресурсов и тем расширяют поле деятельности гибридизатора. Вместе с тем эта проблема вплотную подводит его к следующим проблемам — к проблеме гетерогенного оплодотворения и к проблеме жизнеспособности гибридного эмбриона.
Гетерогенное оплодотворение
Гетерогенным оплодотворением будем называть такое оплодотворение, такое внедрение сперматозоида в яйцо и возбуждение его к развитию, при котором яйцо и сперматозоид, участвующие в оплодотворении, относятся к разным видам, родам, семействами пр. Всегда, когда возникает гибрид, имеет место гетерогенное оплодотворение, но не обратно, ибо не всегда в результате гетерогенного оплодотворения возникает гибрид. Нередко гибридный эмбрион возникает лишь на самый краткий срок и затем погибает, или нее хотя эмбрион и получается, но он оказывается негибридным.
Об этом мы упоминали уже при обсуждении проблемы предела гибридизации. Впервые такое явление наблюдал Леб (1910) при гибридизации иглокожих. Он обратил внимание на Парадоксальный, казалось, факт, что при очень далеких скрещиваниях, которые мы называем ультрагибридизацией, возникающие эмбрионы обнаруживают почти в чистоте материнские признаки и совершенно не обнаруживают отцовских признаков. И это в то время, когда более близкие скрещивания дают гибридов явно промежуточных то своим признакам между признаками родителей. Так, при оплодотворении яиц морского ежа Strongylocentrotus purpuratus близко родственным ему видом St. franciscanus плютеусы (личинки) обнаруживали весьма ясно свой гибридный характер (рис. 85 и 86, глава IX), о чем можно было легко судить по строению первичного скелета. Наоборот, при оплодотворении яиц того же вида спермой морских звезд (относящихся уже к другому классу типа иглокожих, т. е. имеющих отличие от ежей примерно такого порядка, как млекопитающие от птиц или амфибий), что происходило лишь при помощи специальных приемов (щелочью или кальцием), получающиеся плютеусы имели вид, типичный для Strongylocentrotus, скак если бы сперма не принесла в развивающийся зародыш никакого наследственного материала» (рис. 39) (см. Loeb, King and Moore, 1910).
 Рис. 39. Псевдогибридные плютеусы морского ежа Strongylorentrotus purpuratus при оплодотворении спермой морских звезд (по Лебу, Кинг и Муру). (Ср. рис. 86 и 87).
Рис. 39. Псевдогибридные плютеусы морского ежа Strongylorentrotus purpuratus при оплодотворении спермой морских звезд (по Лебу, Кинг и Муру). (Ср. рис. 86 и 87).
Дальнейшими исследованиями было установлено, что при других экспериментальных условиях (комбинация искусственного партеногенеза с естественным осеменением) возможно получить плютеусов,
у которых одна половина тела будет иметь гибридный характер, а другая — опять-таки чисто материнский (Гербст), причем гистологическим анализом было выяснено, что ядра в гибридной половине крупные, диплоидные, а в материнской половине — мелкие, гаплоидные. Очевидно, что в этом случае оплодотворенной оказалась не вся зигота, а только один из бластомеров (рис. 40).
 Рис 40. Половинные гибриды Strongylocentrotus × Sphaerachinus; при а — справа ядра гаплоидной, слева диплоидной части, при b — наоборот (по Гербсту).
Рис 40. Половинные гибриды Strongylocentrotus × Sphaerachinus; при а — справа ядра гаплоидной, слева диплоидной части, при b — наоборот (по Гербсту).
В вышеприведенном случае ультрагибридизации дело обстояло так, что чужеродный сперматозоид хотя и проникал в яйцо и производил оплодотворение его, возбуждая к развитию, но сам (в частности его ядро) участия в дальнейшем развитии не принимал. Здесь получаем как бы искусственный партеногенез при помощи чужеродного сперматозоида. Потомки оказываются вовсе не гибридами. Этих потомков Г. Гертвиг предложил называть ложными гибридами (pseudonoth) в противоположность настоящим гибридам (ortonoth).
Однако правильнее вовсе не называть их гибридами или же противопоставлять их уже не «настоящим гибридам» (ортонотам), а просто гибридам. Ложные гибриды могут быть получены разными способами (например повреждением спермы радием — Гертвиги Г. и П.), но гибридизатора они интересовать особенно не могут, так как тут никакой гибридизации нет, а получаются действительно материнские формы (при этом гаплоидные) и дело идет о своеобразном искусственном партеногенезе.
Что касается гетерогенного оплодотворения, в результате которого мы получаем гибридов, то наши сведения о нем сводятся в основных чертах к следующему.
1. Гетерогенное оплодотворение может происходить в очень многих случаях, начиная от близких видов до весьма отдаленных.
2. В общем может быть констатировано, что чем дальше отстоят друг от друга скрещиваемые виды, тем затруднительнее становится оплодотворение и наконец делается вовсе невозможным.
3. Однако строгого параллелизма между таксономической отдаленностью видов и легкостью гетерогенного оплодотворения также не существует, как и между другими элементами получения гибридов (гетерогенным осеменением, жизнеспособностью гибридов).
4. Во многих случаях гетерогенное оплодотворение возможно тогда, когда возникающие эмбрионы оказываются нежизнеспособными.
5. Это однако не исключает важной для гибридизации возможности возникновения жизнеспособных гибридов там, где без специальной помощи гетерогенное оплодотворение не происходит.
6. У нас уже имеются возможности оказывать содействие гетерогенному оплодотворению и получать его и в тех случаях, когда оно естественно не происходит.
Последние два момента выдвигают проблему гетерогенного оплодотворения как самостоятельную проблему гибридагогии. Пункт 6 позволяет нам путем дальнейшей разработки теории и практики помощи гетерогенному оплодотворению расширять гибридоресурсы. Правда, пункт 5 нам пока приходится в значительной мере постулировать, но верность этого постулата оправдывается например тем, что: 1) часто близкие виды обнаруживают совершенно различную способность к гетерогенному оплодотворению (например упомянутый случай Fundulus heteroclitus и F. majalis); 2) при гетерогенном оплодотворении обычно получается низкий процент оплодотворений, даже когда полученные гибриды вполне жизнеспособны (следовательно в этих скрещиваниях оплодотворения многих яиц не произошло, хотя эмбрион
мог бы быть вполне жизнеспособным); 3) нередко при реципрокных скрещиваниях одно — оплодотворение получается, а реципрокное не получается, хотя можно думать, что это не связано, с жизнеспособностью гибридов. Только изучив досконально процесс гетерогенного оплодотворения и овладев им, гибридагог сможет выделить в полной мере ту часть гиркона, которая способна давать жизнеспособное гибридное потомство, и очертить тем поле своей деятельности, которое можно будет обрабатывать без разрешения следующей и притом труднейшей проблемы: борьбы с нежизнеспособностью гибридов — проблемы, которая конечно в полной мере, по крайней мере для ультрагибридов, разрешена не будет.
Говоря о проблеме гетерогенного оплодотворения, нужно иметь в виду, что в настоящее время она изучена еще чрезвычайно неполно и притом крайне односторонне. Односторонне в том отношении, что объектами исследований служили лишь виды с внешним оплодотворением — оплодотворением, происходящим вне организма самки в воде, — иглокожие, молюски, амфибии, рыбы и т. п. Что же касается объектов, — особенно интересных (кроме рыб) для народнохозяйственной гибридизации — птиц и млекопитающих, то здесь мы пока почти ничего еще не знаем. У этих объектов оплодотворение столь тесно переплетено с осеменением, что необходимо разработать специальную методику осеменения или оплодотворения in vitro (см. ниже) для того, что бы мы имели возможность изучать эти проблемы раздельно. В самом деле, если мы сейчас констатируем неудачу получения гибридов овец с козами, собаки с лисой или если мы констатируем, что при осеменении фазаном курицы получается низкий процент гибридных цыплят, то мы еще ничего не можем сказать, является ли причиной этого трудность гетерогенного оплодотворения или просто сперматозоиды не доходят в чуждом для них яйцеводе до встречи с яйцом, отмирая по дороге (пли наконец, что эмбрионы оказываются нежизнеспособными).
С другой стороны, мы можем ожидать, что именно у животных с внутренним оплодотворением перспективы гетерогенного оплодотворения
могут оказаться особенно широкими. В самом деле несомненно, что виды имеют все основания избегать гетерогенного оплодотворения, в большинстве случаев способного нанести им лишь вред (нежизнеспособностью гибридов или их бесплодием). Поэтому несомненно, что на базе естественного отбора у видов должны были вырабатываться защитные приспособления против возможности гетерогенного оплодотворения. У видов с внешним оплодотворением Такие защитные приспособления должны были приурочиваться к самим гаметам, так как после выбрасывания их в воду родители уже не могут предохранить гаметы от гетерогенного оплодотворения, особенно в богатых фаунах, где по выражению Леба во время нереста морская вода превращается в «суспензию сперматозоидов».
Иное дело например у млекопитающих. Здесь гетерогенному оплодотворению поставлены как правило непреодолимые преграды задолго до проникновения сперматозоида к самому яйцу: экологические преграды, мешающие встрече разных видов, рефлексологические преграды, делающие невозможным сближение самцов одного вида с самками других, механические преграды, делающие невозможным совокупление и эякуляцию, и наконец физиологические преграды, затрудняющие или делающие невозможным для чужого сперматозоида прохождение по длинным путям чужих половых органов до встречи с яйцом. Несомненно, что перечисленные элементы во многих случаях нацело устраняют возможность гетерогенного оплодотворения у млекопитающих и птиц в природе, и «службу защиты вида» от такого оплодотворения естественный отбор может всецело возложить на эти элементы. А так как всякой приспособление при утрате своего значении вообще говоря разрушается в ходе эволюции, то имеются все основания ожидать, что у видов с внутренним оплодотворением сами гаметы лишены уже защитных приспособлений против гетерогенного оплодотворения. Наше предсказание может оказаться неверным лишь при наличии более общих приспособлений клеток животных против проникновения в них инородных белков (реакция типа иммунитета). Однако известные опыты Мерфи (см. ниже главу VII) показали, что например у цыпленка на ранних стадиях его развития ткани эмбриона столь толерантны к чужим белкам, что допускают пышный рост совершенно чуждых трансплантатов (например тканей человека). Можно думать поэтому, что и чужой сперматозоид будет встречаться яйцом млекопитающих и птиц очень толерантно.
Эти соображения заставляют считать изучение гетерогенного оплодотворения у млекопитающих и у птиц (и разработку соответствующей техники) проблемой, чрезвычайно интересной как в гибрида готическом, так и в общебиологическом отношении.
Процесс оплодотворения и его элементы
Для того чтобы овладеть гетерогенным оплодотворением, получить возможность управлять им и устранять препятствия, необходимо изучить все различные их элементы, входящие в этот сложный процесс, и вскрыть все многочисленные отдельные препятствия, которые могут встать на пути гетерогенного оплодотворения.
Можно наметить следующие элементы процесса оплодотворения:
1) возбуждение яйцом подвижности сперматозоидов;
2) привлечение яйцом сперматозоида и тактическую реакцию сперматозоида на близость яйца;
3) внедрение сперматозоида в яйцо;
4) выделение яйцом оболочки, предохраняющей от полиспермии, без чего часто невозможно правильное оплодотворение;
5) возбуждение яйца к дроблению;
6) движение ядра сперматозоида к ядру яйца и слияние с ним;
7) участие обоих ядер в дроблении.
На каждом из перечисленных участков могут возникнуть препятствия к гетерогенному оплодотворению, и борьба за него должна вестись по всем участкам этого фронта.
Усиление активности сперматозоида
Усиление активности сперматозоида при приближении к яйцу наблюдается у многих видов. Повидимому оно особенно свойственно некоторым гетерогенным соединениям; так, Дунгерн (Dungern) считал, что такое усиление активности свойственно только чужой сперме. Лилли (1914) высказал предположение, что это явление может носить специфический характер. Лоб (1915)
[30] исследовал этот вопрос экспериментально на морских ежах и звездах. У морских звезд Asterias и Asterina были взяты незрелые яйца, а у морских ежей Strongylocentrotus franciscanus и S. purpuratus — яйца зрелые. У тех же видов была взята сперма и помещена, как и яйца в
п/
2 растворе NaCl. Здесь сперматозоиды оставались неподвижными. Затем производилось смешение каждого вида яиц с каждым видом спермы в том же растворе. При этом в ряде комбинаций наступала яркая активность, в других же — нет, как видно из таблицы, на следующей странице.
Из этой таблицы можно сделать ряд поучительных выводов. Прежде всего видно сильное возбуждающее действие яиц на сперматозоиды того же вида (клетки большой диагонали, идущей слева сверху. направо книзу). При этом возбуждающее действие оказывают на сперматозоиды, как зрелые, способные к оплодотворению яйца, так и незрелые, к оплодотворению совершенно еще не способные.
Специфичность в активировании спермы яйцами

Действие яиц на чужие сперматозоиды, наоборот, очень разнообразно. Так, яйца обоих видов морских ежей действуют одинаково возбуждающе как на сперму того же вида, так и на сперму другого вида морских ежей. Наоборот, на сперму морских звезд (дело идет, надо помнить, об ультрагибридизации — скрещиваются представители разных
классов) яйца ежей действия не оказывают совсем или очень слабое.
Яйца Asterias ведут себя еще более специфично и, возбуждая свою сперму, совершенно не возбуждают спермы другого рода морских звезд. Несколько своеобразным оказывается их более-заметное, хотя и слабое влияние на сперму морских ежей — форм, во всяком случае гораздо более удаленных, чем Asterina.
Наоборот, у Asterina мы видим совершенно неожиданное явление: ее яйца одинаково сильно возбуждают свою сперму и сперму одного из ежей — St. franciscanus, очень слабо — сперму другого вида ежей и совершенно не возбуждают сперматозоидов Asterias, другой звезды.
То же можно сказать и обратно, исходя от спермы, но придется лишь отметить, что сперма морских звезд сильно реагирует на свои яйца и почти или вовсе не реагирует на чужие яйца; сперма St. purpuratus реагирует на яйца обоих видов ежей и слабо — на яйца морских звезд. И наконец сперма St. franciscanus реагирует ярко на свои яйца и яйца другого вида ежей и столь же ярко на яйца морской звезды Asterina и наконец на яйца Asterias реагирует «умеренно активно».
Обсуждая эти опыты, Леб придает повидимому большее значение возбудимости сперматозоида, чем возбуждающему действию яйца. Ол указывает, что и в морской воде для-возбуждения сперматозоидов звезд требуется более сильное воздействие, чем для сперматозоидов ежей.
Во всяком случае несомненно, что проблема возбуждения сперматозоидов яйцами еще очень неясна. Подчеркнем например дне реципрокные встречи Asterina с St. franciscanus: в то время как сперматозоиды St. franc, ярко реагируют на яйца Asterina, при реципрокной встрече не получается никакого эфекта. Уже это одно говорит об отсутствии какой-либо
простой связи между возбудителем и таксономическим родством — вывод, чрезвычайно важный для гибридагогии, так как мы видим, что возбуждение может иметь место и при чрезвычайно отдаленных встречах (звезда — еж), которые имеют лишь-общетеоретический интерес, а не практический — вряд ли мы будем-пытаться скрещивать голубя с кроликом, возможность чего допускал. Реомюр.
Необходимо иметь в виду и то, что сам вопрос о роли возбуждения сперматозоида еще не ясен. Так, Леб и Уэстенейс (Loeb a. Wasteneys) — тщетно пытались обнаружить активирующее действие яиц разных морских костистых рыб на сперму тех. же видов.
Таким образом в этом разделе мы встречаемся с необходимостью-дальнейшей разработки следующих проблем: 1) значение возбуждения сперматозоидов яйцом для оплодотворения в связи с различными условиями этого оплодотворения; 2) какими веществами вызывается это возбуждение у разных видов, являются ли эти вещества специфическими видовыми или родовыми, или более общими, какова химическая природа этих веществ (см. ниже), какова природа возбудимости сперматозоидов и какими факторами это возбуждение можно вызвать и т. д.
Очень интересны были бы опыты со смешанными порциями яиц, своего и чужого вида. В предыдущей главе мы видели например опыты Штандфусса, возбуждавшего самцов бабочек, помещая в одну клетку самок как того же, так и другого вида — таким способом ему удавалось заставить возбужденных самцов итти на самок другого вида. Эту идею можно было бы применить и здесь и возбудить активность сперматозоидов некоторым добавлением «своих» яиц, быть может облегчив им проникновение в чужие яйца.
Далее в качестве возбудителя должна быть изучена температура, так как влияние повышения температуры на подвижность спермы уже известно. Несомненно, что поле для экспериментальных исследований под углом зрения
содействия гетерогенному оплодотворению здесь еще чрезвычайно широко.
В настоящее время в некоторые из перечисленных вопросов уже внесена доля ясности. Работами главным образом Ф. Р. Лилли, Леба и др. было показано, что в активности сперматозоидов большую роль-играет концентрация в воде водородных ионов и гидроксильных ионов, причем гидроксил-ионы повышают активность, а
Н-ионы оказывают тормозящее действие. Что последнее в частности имеет место у некоторых видов, доказывает явление агрегации сперматозоидов: скопляясь случайно вместе, сперматозоиды выделяют такое количество СО
2, что их движение замирает, а к образовавшейся кучке направляются другие благодаря, очевидно положительному хемотаксису к повышенной концентрации СО
2 — и комки сперматозоидов еще больше нарастают, становясь. видимыми простым глазом (например у червя Nereis).
Более детально природу веществ, выделяемых яйцами, можно исследовать методом так называемой «яичной воды». Помещая в небольшой объем воды значительные количества яиц, можно получить настойку, которую далее можно слить с яиц и изучать действие «яичной воды» на сперматозоиды в отсутствие яиц.
Эта яичная вода (с яиц иглокожих и червей) обладает сильным действием, способным активировать сперматозоиды, вызывать агломерацию и внешне сходную с нею аглютинацию, т. е. слипание, связанное очевидно с каким-то изменением поверхности головки сперматозоида, делающим ее липкой и вероятно играющим роль при оплодотворении.
Лилли установил некоторые черты вещества (веществ), выделяемого яйцами. Оно бесцветно, не проходит через фильтр Беркефельда и через уплотненную фильтровальную бумагу, не подвергается диализу, устойчиво по отношению к высоким температурам и медленно разрушается при кипячении. По некоторым данным сила действия этого вещества пропорциональна квадратному корню на концентрации. Эти признаки указывают на сходство этого вещества с ферментами. Признавая за ним видную роль в оплодотворении, Лилли назвал его «фертллизином» — веществом оплодотворения.
Следует однако иметь в. виду, что подобное вещество удается установить лишь у немногих видов, и обобщать точку зрения Лилли нельзя, на чем особенно настаивает Леб. Нужно также иметь в виду, что эти исследования относились к немногим видам с внешним оплодотворением — и у таких форм, как млекопитающие и птицы, отношения могут быть существенно иными. Так например активизация спермы у млекопитающих и птиц достигается не выделениями яйца, а секретами добавочных желез мужского полового аппарата, и эякулированная сперма у млекопитающих и птиц находится в бурном движении без всякого участия яиц.
Таким образом анализ отношений между яйцом и сперматозоидом должен быть проведен для различных форм конкретно. Данные, полученные на иглокожих и пр., совершенно невозможно переносить на такие формы, Как млекопитающие и птицы, так как там сперматозоиды являются аэробами, а здесь — анаэробами.
Привлечение сперматозоида в яйцу
Следующей кардинальной для гибридагогии проблемой является вопрос о том, происходит ли встреча сперматозоида с яйцом в случайном порядке, или же сперматозоид специально привлекается яйцом каким-либо специфическим образом, благодаря которому к яйцу направляются сперматозоиды именно того же вида, а не чужие. Если такого специфического привлечения нет, то задача гибридизатора сразу упрощается — для того чтобы обеспечить встречу, ему достаточно привести яйцо, подлежащее гетерогенному оплодотворению, в соприкосновение с каплей возбужденных сперматозоидов. Если же необходимо со стороны яйца специальное и специфическое привлечение сперматозоида, то проблема осложняется, и гибридизатору необходимо ее разрешать. Впрочем вряд ли вопрос стоит здесь в порядке альтернативы; вероятнее, что в разных случаях в зависимости от различной экологии оплодотворения у разных видов дело обстоит различно, и гибридизатору предстоит исследовать эту конкретную специфику.
Еще со времен Пфефера известно хемотропическое привлечение сперматозоидов папоротника слабыми растворами яблочной кислоты (0,01 %), в то время как живчики мхов не реагируют на эту кислоту, но привлекаются слабым (0,1 %) раствором тростникового сахара; живчики же харовых но реагируют ни на то ни на другое.
Эти наблюдения были многими истолкованы так, что здесь идет речь о специфической реакции сперматозоидов на вещества, выделяемые яйцом как приспособление для привлечения сперматозоидов своего вида. Пфеферу (1881–1885) удалось обнаружить присутствие яблочной кислоты в тех частях папоротника, где размещены архегонии, почему он предположил, что она же имеется и в архегониях. Подобные же эксперименты пропели затем Шибата (Shibata) и Брухман (Bruchmann, 1909). Последний нашел, что сперматозоиды положительно хемотактичны к лимонной кислоте и ее солям. Также хемотактичны они к вытяжке из архегониев. Но при всем том открыть в архегониях лимонную кислоту никак не удается.
Если таким образом у этих растений хемотаксис выращен ясно и может играть роль при оплодотворении, то обобщать эти факты никак нельзя. У иглокожих например ни Буллер (Buller, 1992), ни Дьюитц (Dewitz), ни Леб не обнаружили хемотропизма сперматозоидов к яйцам того же вида. Дэкин (Dakin) и Фордгэм (Fordham, 1924), наоборот, обнаружили явный хемотропизм у морского ежа Echinus esculentus. Лилли настаивал на хемотропизме у видов, с которыми работал и Леб, но Леб решительно возражал ему, как и вообще возражал против гипотезы фертилизина.
Что касается других животных, и в частности млекопитающих и птиц, то данные здесь отсутствуют нацело (см. ниже о работе Пинкуса). Продвижение сперматозоидов вверх по яйцеводам если и носит тропический характер (вероятно реотропизм), то это несомненно не относится к привлечению сперматозоида яйцом.
Таким образом на сегодня этот вопрос остается еще неразработанным и во всяком случае в, положении, совершенно не могущем удовлетворить гибридизатора. Однако можно констатировать, что у некоторых форм, вроде иглокожих, оплодотворение может наступать без всякого хемотропического привлечения сперматозоидов яйцом. Что касается приведенных выше данных ботаников, то Леб считает важным подчеркнуть, что химические вещества, вызывающие тропическое привлечение сперматозоидов к своим яйцам, не имеют по своей химической природе никакого отношения к Test специфическим белкам, которые составляют химическую характеристику вида и дают себя знать в явлениях иммунитета и анафилаксии, преципитиновых реакциях и пр. Это указание Леба представляет несомненный интерес.
Внедрение сперматозоида в яйцо
Находящиеся в момент оплодотворения вокруг яйца сперматозоиды возбужденно двигаются it активно внедряются в яйцо. Наблюдая in vitro (см. ниже) оплодотворение яйца кролика сперматозоидами кролика же, Пинкус рисует картину, напоминающую игру в пушбол (большой мяч). Если яйцо свободно от окружающей его слизи, то сперматозоиды с такой силой толкают его, что яйцо начинает кататься по капле — и оплодотворение даже не может осуществиться.
Однако и яйцо не остается совершенно пассивным при оплодотворении, оно посылает навстречу сперматозоиду так называемый воспринимающий бугорок, который можно сравнить с псевдоподией амебоидной клетки. В таком случае поглощение сперматозоида воспринимающим бугорком яйцеклетки оказывается аналогичным фагоцитозу, захватыванию амебоидной клеткой бактерий и частиц. Мы еще не знаем достоверно природы тех сил, которые действуют в случае фагоцитоза, хотя высказывались предположения относительно сил поверхностного натяжения и аглютинации; оба эти явления имеют место на поверхности и быстро обратимы. Следовало бы сказать поэтому, что специфичность, наблюдаемая при процессе оплодотворения, заключается в отличительных свойствах поверхности яйца и сперматозоида.
Различные авторы согласны, что в процессе оплодотворения огромную роль играет именно поверхность яйца, свойства его кортикального слоя. Именно здесь по мнению как Лилли, так и Леба сосредоточены те препятствия, которые мешают как гетерогенному оплодотворению, так в некоторых случаях и самооплодотворению. Роль кортикального слоя в последнем случае была экспериментально показана Морганом (1923) на Ciona intestinalis, у которой самооплодотворение обычно встречается с большими затруднениями. Зрелое яйцо этой асцидии бывает покрыто слоем фоликулярных клеток, остающихся на яйце в качестве оболочки. Сняв эту оболочку, в результате чего вероятно изменились поверхностные свойства яйца, Морган сделал самооплодотворение легко возможным.
Лилли считает, что и при затруднениях к гетерогенному оплодотворению играет роль тоже кортикальный слой и именно на его изменение необходимо направить в этих случаях внимание.
Впервые такую задачу удачно решил Леб. Нормально яйца морского ежа Strongylocentrotus lividus не оплодотворяются спермой морских звезд. Но желая изменить поверхностные условия, Леб прибег к слабому повышению щелочности морской воды (прибавкой едкого натра, аминов и др.). Попытки его удались, и наилучшие успехи достигались при прибавлении 0,6 см
3 n/
10 NaOH к 50 см
3 морской воды. Большая часть яиц при этом оплодотворялась.
Для характеристики роли щелочи в данном случае интересно отметить, что это гетерогенное оплодотворение наступало только в самом щелочном растворе. Если же Леб подвергал яйца и сперму действию щелочи отдельно, а потом помешал их в нормальную воду, — оплодотворения обычно не наступало. При переносе их вместе снова в подщелоченную воду оплодотворение происходило. Таким образом наличие щелочи производит какое-то быстро. обратимое изменение поверхностных свойств гамет, делающее возможным гетерогенное оплодотворение.
Леб (1903) открыл такой же эфект повышенной концентрации кальция, чем ему удалось добиться оплодотворения яиц ежа St. purpuratus спермой звезды Aster.as ochracea и A. capitata спермой офиурид и голотурий, причем. процент оплодотворения достигал 50. Худшие результаты получились со спермой другой звезды — Picnopodia spuria (5 %) и спермой звезды Asterias (1 %).
Этим же методом Годлевскому (1906) удалось оплодотворить яйца неаполитанской морской звезды спермой морской лилии.
Однако этот, метод не оказался универсальным, и уже сам Леб не смог добиться оплодотворения спермой морской звезды яиц другого уже упоминавшегося калифорнийского ежа — St. franciscanus. Хотя сперматозоиды встречались с яйцами и последние образовывали оболочки, но проникновения сперматозоидов наблюдать не удалось. Не удались также попытки оплодотворения яиц ежа Arbacia спермой звезды Asterias.
Несмотря на эти неудачи, опыты Леба имеют для нас исключительное значение, так как они впервые поставили и экспериментально разрешили проблему помощи гетерогенному оплодотворению. Не надо забывать, что в этих опытах дело идет об ультрагибридизации, и если крупные успехи были получены даже здесь, то тем больших результатов мы можем ожидать от дальнейшей разработки этого химического метода при более близких гетерогенных оплодотворениях. Следует отметить, что эти гениальные открытия, сделанные в Америке 30 лет назад, не получили до сих пор никакого практического использования (например хотя бы в рыбоводстве и т. п.), хотя постоянно цитируются в различных книгах по биологии. Это тем более заслуживает быть отмеченным, что Леб же показал, что эти факторы играют роль не только при гетерогенном оплодотворении, но и при нормальном, так как например оно может быть сделано невозможным простым понижением щелочности и содержанием кальцин в воде. В нейтральной среде нормального оплодотворения яиц морских ежей совершенно не наступало несмотря на отличную подвижность сперматозоидов (Леб, 1914; KupeIvieser, 1909, 1912).
Таким образом очевидно, что, уменьшая щелочность окружающей яйцо среды и удаляя из этого раствора CaCl
2, мы создаем для проникновения сперматозоида Arbacia в яйцо того иге. вида такие же препятствия, какие в нормальной морской воде задерживают проникновение спермы морской звезды в яйцо St. purpuratus. «Препятствие», задерживающее проникновение спермы Arbacia в яйцо того же вида, созданное этим путем, точно так же быстро обратимо
[31].
Сущность самого препятствия к гетерогенному оплодотворению этими опытами вскрывается еще очень мало. Картина была бы проще, если бы при данной щелочности яйцо было более или менее одинаково доступно различным сперматозоидам. Этого конечно нет. Таким образом вопрос о том, почему при данной среде (например нормальной воде) в яйцо легко проникает свой сперматозоид и вовсе не проникает чужой — нимало не решается. Мы лишь получаем — правда, очень важные — указания, что при повышенной щелочности барьер понижается, делая яйцо доступным даже для чужих сперматозоидов, а при нейтрализации барьер повышается, делая яйцо недоступным даже для своих сперматозоидов.
Сущность «барьера оплодотворения» нам пока не ясна, но открыта возможность экспериментального изменения высоты этого барьера.
Изменение щелочности среды является не единственным фактором, при помощи которого можно изменять «барьер оплодотворения». Так, еще в 1887 г. О. и Р. Гертвиги обнаружили здесь роль возраста яиц иглокожих и показали, что если яйцам дать постоять несколько времени, то они становятся более восприимчивыми к чужой сперме.
Несколько иначе смотрит на вопрос Вернон (1900), занимавшийся тоже многочисленными гибридизациями ежей (см. также Теннент, 1910, 1911).
Что касается других объектов помимо упомянутых, обычных в такого рода, исследованиях (иглокожие, амфибии), то сведения здесь, отсутствуют повидимому нацело. Нацело отсутствуют они в частности и для млекопитающих и птиц. Приходится обратить внимание гибридагогов на этот зияющий пробел, дабы принять меры к его заполнению.
По остальным проблемам оплодотворения, перечисленным выше, сказать нечего. Следует однако еще раз вернуться к вопросу о судьбе сперматозоида, проникшего в чужое яйцо. Далеко не всегда это приводит к действительному оплодотворению, т. е. к возбуждению, к развитию с участием обоих ядер. Купельвизер (1909,1912) заметил, что сперматозоиды молюсков и кольчатых червей проникают иногда случайно в яйца ежа St. purpuratus в нормальной морской воде. В этих случаях развития яиц однако не наступало. Но в других случаях развитие наступает, и сказать сейчас, в чем заключается причина различной реакции яиц на чужие сперматозоиды, — еще невозможно.
Интересно однако проследить, как на границе возможного оплодотворения вариирует участие отцовского ядра в развитии гибрида.
Слияние гетерогенных ядер при оплодотворении
Нормальное оплодотворение должно сопровождаться более или менее полным слиянием семенного и яйцевого ядер в процессе оплодотворения.
В гибридных оплодотворениях наблюдающиеся здесь явления, оказываются весьма разнообразными, открывая широкое поле для дальнейших исследований и попыток вмешательства в этот процесс.
В случаях ультрагибридизации, как например при сочетании морской еж × молюск (Купельвизер, 1909), мы видим гибель ядра сперматозоида, и если на некоторых препаратах мы и наблюдаем картину как бы слияния ядер, то это вероятно более или менее случайная картина. Во всяком случае уже в первом делении дробления участвует только материнское ядро. В другом аналогичном случае оплодотворение яиц ржа Echinus спермой кольчатого-червя (Купельвизер, 1912) хотя слияние ядер почти всегда и наблюдается, но уже в первом делении хромосомы отца ведут себя ненормально — не включаются в дочерние ядра первых двух бластомеров. В несколько менее отдаленных скрещиваниях (еж × лилия, Годлевский, 1906) слияние ядер происходит благополучно, и хромосомы обоих родителей участвуют (изучено недостаточно!) в дроблении яйца.
Тем интереснее, что у рыб при значительно более близких скрещиваниях слияние ядер идет с большими затруднениями. Уже по материалам, полученным Менкхгаузом (1904) при гибридизации Fundulus ♂ × ♀ Menidia, можно видеть, пользуясь различием в хромосомах обоих видов, что хотя слияние ядер при этом гетерогенном оплодотворении и происходит, но в веретене первого дробления оба типа хромосом, не перемешиваясь, расположены билатерально двумя группами. В третьем делении дробления эта группировка уже нарушается, и по мере дальнейших делений происходит полное перемешивание в митозах хромосом обоего типа, хотя разница в этих типах сохраняется попрежнему.
Совершенно такую же картину наблюдала М. Моррис (1904) при оплодотворении яиц Fundulus спермой Ctenolabrus. Хромосомы этих рыб тоже отчетливо различны; палочкообразные — у первого вида, точкообразные — у второго (рис. 91).
При сравнении процесса слияния ядер при гомо- и гетерооплодотворении и ясно заметна разница. В чистом оплодотворении оба ядра, подходя друг к другу, быстро сливаются, и только уже после этого центросома делится на две дочерние, и начинается первое деление… При встрече чуждых ядер, наоборот, граница между ними ясно сохраняется и тогда, когда сияют уже две центросомы. Далее после того как оболочка ядер исчезает, видны еще в веретене два остатка ядра, и когда оба ядра наконец сливаются в метафазе — круглые и палочкообразные хромосомы лежат двумя неперемешанными группами. Та же картина сохраняется — и в анафазе и лишь в следующих делениях в расположении хромосом не чувствуется изолированности.
Ультрагибридизация
Гибридизация морских ежей с молюсками. В опытах оплодотворения яиц морских ежей спермой молюсков мы имеем дело с попытками гибридизации крайней степени — речь идет о представителях различных
типов животного царства. Более отдаленными гетерогенными сочетаниями будут уже лишь сочетания Metazoa × Protozoa и животные × растения. Так как тем не менее в этой «фантастической» гибридизации иглокожих с молюсками мы встречаем ряд интереснейших явлений вплоть до почти правильного оплодотворения, то интересно изучить эти явления подробнее. Здесь мы имеем краткую, но содержательную работу Леба (1908) и подробную работу его ученика — Купельвизера (1909). Они несколько различаются и объектами: у Леба — морской еж Strongylocentrotus franciscanus. и молюск Chlorostoma funebrale, у Купельвизера — преимущественно еж Echnus microtuberculatus и молюск Mytilus.
Для успеха опытов необходимы чистота и предохранение от инфекций. Животные тщательно обмывались пресной водой, по вскрытии гонады тщательно отмывались морской водой, взятой из чистого места, отфильтрованной, нагретой до 70° и снова охлажденной до температуры морской воды, и помещались в сосуды с морской водой. Для опытов использовались яйца, сами собой выходившие из гонад. Mytilus выбирались лишь со. зрелой спермой и притом хорошоподвижной. Оказалось полезным перед опытами держать сперму на воздухе
1/
2—1 час без воды или с небольшим количеством ее — это способствует оживлению спермы.
Для опытов яйца раскладывались порциями в сосуды с 50 см
3 воды, и прибавлялась сперма, предварительно разбавленная вдвое, в разные сосуды в таком количестве: 0 (контроль); 0,2; 0,4 … 2 см
3. Пребывание яиц вместе со спермой («экспозиция») ограничивалось 3 часами, после чего яйца переносились в свежую воду.
Оказалось однако, что-эта техника, пригодная для St. purpuratus, недостаточна для St. lividus и для Echinus. Яйца последнего покрыты студенистой оболочкой, абсолютно непроходимой для спермы молюсков, к которой они только густо прилипают. Оболочка может быть; уничтожена кратким содержанием в подогретой воде или 1–1
1/
2-минутной обработкой раствором 0,5 см
3 n/
10 HCl в 50 см
3, морской воды. Наилучшим способом является однако механический, путем сильного потряхивания. Часть яиц при этом разрушается, но часть утрачивает лишь оболочку.
К освобожденным от оболочки яйцам Echinus прибавлялось 5—10 капель полуразбавленной спермы и 50 см
3 воды, экспозиция здесь продолжалась не более 1 часа. Более длительная экспозиция оказывалась вредной, так как после проникновения спермы Mytilus в яйцо Echinus последнее не образовывало оболочки, предохраняющей от — внедрения других сперматозоидов, и в большинстве случаев это приводило к полиспермии (наблюдалось внедрение до 17 сперматозоидов). Между тем дальнейшее правильное дробление возможно лишь при моноспермии. Именно полиспермией и объясняется низкий процент нормального развития яиц Echinus × Mytilus, в среднем равнявшийся у Купельвизера 3,7 % до бластулы и 1 % до гаструлы.
Леб применил несколько иную методику, работая с яйцами St. franciscanus и спермой молюска Chlorostomi funabrale. Яйца одной самки раскладывались равными порциями в сосуды с 50 см
3 морской воды и к ней прибавлялось
n/
10 NaOH в различном количестве: 0,0; 0,2; 0,4; 0,6;0,8; 1,0. Оплодотворение наступало очень быстро. В сосуде с 0,8 NaOH до 80 % яиц почти тотчас образовывали мембрану, в 0,6 NaOH — меньше, а 1,0 NaOH вызывало уже аглютинацню спермы. Рядом опытов Леб доказал, что здесь происходил не искусственный партеногенез химическим раздражением, а действительно процесс гетерогенного оплодотворения (Леб, 1908). Повторные опыты дали столь же прекрасные результаты
[32], и Леб получил тысячи нормально развитых плютеусов. Плютеусы эти все без исключения были типичными для St. franciscanus (рис. 41).
 Рис. 41.Псевдогибридные плютеусы Strongylocentrotus franciscanus при оплодотворении спермой молюска Chlorostoma (по Лебу). (Ср. рис. 87 и 88).
Рис. 41.Псевдогибридные плютеусы Strongylocentrotus franciscanus при оплодотворении спермой молюска Chlorostoma (по Лебу). (Ср. рис. 87 и 88).
Яйца Echinus значительно прозрачнее яиц крупного St. franciscanus, что позволило Купельвизеру дать подробное описание явления оплодотворения in vivo. Если студенистая оболочка яйца не была удалена совершенно, сперматозоиды прилипали к ней хвостами, Впрочем и Леб и Купельвизер отмечают, что у довольно значительного числа самок опыт не удается вовсе — очевидно в связи с какими-то индивидуальными свойствами их яйцеклеток, оставаясь на ее поверхности, пока все яйцо не обволакивалось ими. Внутрь оболочки ни разу сперматозоиды не попадали. При удалении оболочки сперматозоиды упираются в яйцо и пытаются внедриться в него. В большинстве случаев однако им удается воткнуть лишь острие головки, и они затем либо снова отрываются либо остаются длительно в таком положении. Изредка удается однако увидеть проникновение головки, после чего в течение четверти часа при помощи оживленных движений хвостика сперматозоид погружается в яйцо целиком. Никакого воспринимающего бугорка яйцо навстречу этому чужому сперматозоиду не образует, и проникновение происходит в любом месте, что доказывается уже одновременным прониканием многих сперматозоидов в различных местах. После проникновения, как упомянуто, не происходит образования мембраны, почему проникновение сперматозоидов продолжается, и моноспермия является исключением.
Приблизительно через полчаса от начала опыта можно видеть проникшие сперматозоиды в виде больших, сильно преломляющих свет телец в протоплазме яйца. Через 10–20 минут возле такого тельца возникает отчетливое сияние, лучистая сфера. Лучистая сфера сперматозоида, проникшего первым (часто также и второго и третьего), начинает правильно, хотя и медленно, двигаться к ядру яйца, ведя за собой и ядро (пронуклеус) спермия. (Сперматозоиды, проникшие позднее, также образуют сферы, но остаются на периферии и могут не поворачиваться сферой к ядру яйца.) Далее образуется уже центральное слияние, и оболочка ядра яйца исчезает, начинается первое дробление. Нередко это центральное слияние захватывает и другие сперматозоиды, — образуется многополюсный митоз, и яйцо гибнет.
Темп развития идет нормально. По данным, полученным на фиксированном материале, сроки наступления стадий таковы:
Центральное сияние — 1 час 13 мин.
Веретено — 2 часа 5 мин.
Два бластомера — 3 часа 0 мин.
Бластула — 17 часов 15 мин.
На фиксированном материале Купельвизер изучил и более детальные процессы, происходящие при этом. Они дополняются следующими существенными деталями: ядро сперматозоида, проникнув в яйцо, несколько изменяется, разбухает, что впрочем свойственно сперматозоидам Mytilus и при нормальном оплодотворении. Направляясь вслед за, своей центросомой к ядру яйца, ядро сперматозоида однако дальше не обнаруживает никакой активности. В то время как ядро яйца теряет оболочку и распадается на свои нормальные 9 хромосом (рис. 42), ядро сперматозоида остается округлым, достаточно плотным тельцем и лишь изредка образует нечто вроде хромосом. Во всяком случае уже в первом делении оно не принимает участия, и лишь в виде исключения
[33] оказывается в веретене, но попрежнему в виде цельного тельца. В результате при первом же делении ядро сперматозоида оказывается в одном из бластомеров, а при втором дроблении Купельвизером уже не было найдено. Общий итог своих наблюдений Купельвизер выражает следующей схемой (рис. 43).
 Рис. 42. Поведение ядер при оплодотворении спермой молюска яйца морского ежа (по Купельвизеру).
На левом рисунке маленький пронуклеус сперматозоида виден при правой центросоме; на правом рисунке — при верхней центросоме. На правом и среднем рисунке видно гаплоидное число хромосом.
Рис. 42. Поведение ядер при оплодотворении спермой молюска яйца морского ежа (по Купельвизеру).
На левом рисунке маленький пронуклеус сперматозоида виден при правой центросоме; на правом рисунке — при верхней центросоме. На правом и среднем рисунке видно гаплоидное число хромосом.
Обсуждая эти выводы, Купельвизер отмечает их полнейшее, до мельчайших подробностей совпадение с аномальным процессом оплодотворения яиц Echinus его же спермой, но обработанной едким кали (см. Boveri, 1888; Teihmann, 1909). Ядро сперматозоида в обоих случаях оказывается убитым, и яйцо и дальнейший организм получают от сперматозоида лишь центросому, нормальный характер которой описывает детально Купельвизер.
Гибель хромосом, показанная Купельвизером, объяснила, почему полученные им и Лебом плютеусы оказываются чисто матроклинными. Вместе с тем она лишила обоснования и вывод Леба, сделанный им в 1908 г. что «чисто материнский характер гибридов еж × молюск указывает на то, что не хромосомы являются носителями наследственности, а что передача наследственных особенностей есть проблема чисто химическая, но не гистологическая».
 Рис. 43. Схема оплодотворения яйца морского ежа спермой молюска (по Купельвизеру).
Пронуклеус яйца изображен кружком или рядом хромосом на веретене (4); пронуклеус сперматозоида — точкой при луче сияния.
Рис. 43. Схема оплодотворения яйца морского ежа спермой молюска (по Купельвизеру).
Пронуклеус яйца изображен кружком или рядом хромосом на веретене (4); пронуклеус сперматозоида — точкой при луче сияния.
Гибридизация морских ежей с морскими лилиями осуществленная в 1904–1906 гг. Годлевским (Godlewsky Е. jun., 1906), представляет. как несколько более близкая, межклассовая, ряд интересных моментов для сравнения с данными Купельвизера. Годлевским была произведена гибридизация по методу Леба яиц морских ежей Sphaerechmus granularis, Echinus microtuberculatus u Strongylocentrotus lividus спермой лилии Antedon rosacea. Так же, как и в скрещивании еж × молюск, полученные плютеусы оказались чисто материнскими, и ни в темпе деления, ни в образовании мезенхимы, ни в самом плютеусе не было обнаружено никаких изменений, которые можно было бы приписать влиянию отцовского вида. Однако в гистологическом отношении имеются интересные отличия от картины, полученной — Купельвизером в скрещивании еж × молюск.
 Рис. 44. Процесс оплодотворения яиц морского ежа спермой морской лилии (по Годлевскому).
Рис. 44. Процесс оплодотворения яиц морского ежа спермой морской лилии (по Годлевскому).
Морские лилии и морские ежи представляют собой две чрезвычайно древние
[34] ветви иглокожих, разделившиеся еще до палеозоя, сотни миллионов лет назад. Тем замечательнее факт, что оплодотворение яиц морских ежей спермой лилии происходит совершенно нормальным образом. Если исключить то, что для оплодотворения необходимо некоторое повышение концентрации ОН-ионов, то во всем остальном процессе нельзя заметить никаких уклонений от нормального процесса оплодотворения.
Проникнув в яйцо, сперматозоид образует лучистую сферу, поворачивается ею по направлению к ядру яйца, начинает двигаться к атому ядру, очевидным образом оба ядра сливаются в одно, В этом последнем пункте у еж × молюск было отличие, так как картины слияния ядер с полной убедительностью установить было невозможно, и даже в тех случаях, когда ядро молюска оказывалось в тесном контакте с ядром ежа, первое имело все же характер некоторого инородного тела. Рис. 44 показывает картину типичного слияния ядер и совместное участие обоих уже в первом веретене. Точный анализ хромосом здесь проделан не был, и так как хромосомы Antedon были не учтены, а для ежей оставалось невыясненным, не существует ли двух рас, с 9—18 и 18–36 хромосомами, то по полученной пластинке метафазы, на которой можно насчитать несколько более 20 хромосом, установить уверенно судьбу хромосом лилии невозможно. Однако каких-либо признаков элиминации хромосом в первом делении Годлевский не видел.
Полученные от Echinus × Antedon и Strongylocentrotus × Antedon плютеусы ничем не отличимы от контрольных чистых видов, и можно думать, что рано или поздно эта элиминация произошла. Однако вопрос этот не вполне ясен (о нем см. ниже — в главах VII и IX).
Наконец при гибридизации, еще более — близкой, ежей Strongylocentrotus lividus с ежом Arbacia Бальцер (Baltzer, 1909) хотя и получил тоже матроклинных личинок, отцовские хромосомы элиминировались уже лишь на стадии бластулы (см. главу VII).
Заканчивая краткий обзор фактов, относящихся к гетерогенному оплодотворению, необходимо отметить несмотря на их огромный интерес и значение для гибридагогии их крайнюю неполноту и односторонность. Неполнота состоит в том, что ни на один вопрос, нас интересующий, мы не имеем еще ответов, настолько глубоко проработанных, чтобы они могли служить путеводителем в работе гибридизатора. На основной вопрос — почему в одних случаях при гибридизации мы получаем оплодотворение, а в других нет — мы удовлетворяющего ответа не имеем кроме ответа общего, что чем дальше отстоят друг от друга скрещиваемые формы, тем меньше шансов на успех. Также и на второй основной для нас вопрос — что нужно делать для того, чтобы заставить произойти оплодотворение там, где оно не идет, — мы также почти не имеем ответа кроме отдельных указаний эмпирического характера, касающихся некоторых иглокожих. Очевидно здесь предстоит еще огромная исследовательская работа со всевозможными объектами.
До сих пор мы говорили о гетерогенном оплодотворении как о методике получения F
1 гибридов. Не менее интересна теоретически и важна практически и проблема гибридного оплодотворения, т. е. оплодотворения гибридных яиц и оплодотворения гибридной спермой. Здесь вопрос стоит так: какая сперма должна лучше всего оплодотворять гибридное яйцо (яйцо гибрида) и какие яйца должны — лучше всего оплодотворяться гибридной спермой? Этот вопрос имеет ту же самую формулировку, как и соответствующий вопрос в главе об осеменении.
В самом деле в пределах вида А. свойства яйца и свойство сперматозоида так взаимно согласованы, что обычно яйцо оплодотворяется легче всего спермой, того же вида и обратно — свойства спермы приспособлены к свойствам яйца того же вида. Но у гибрида свойства его яиц и спермы очевидно уже не те, что у чистых видов. Специальному приспособлению друг к другу яйца и сперма этих гибридов не подвергались. Значит ли это, что все-таки яйца гибрида будут легче всего оплодотворяться спермой подобного же гибрида или нет, или может быть более подходящей будет сперма родительских видов и т. д.? Для размножения гибридов эти вопросы их оплодотворения будут иметь конечно большое значение. Тем печальнее, что мы не располагаем по этому поводу нужными фактами.
Гетерогенное оплодотворение теплокровных in vitro
Если прямые попытки получения таких гибридов, как овца × коза, корова × буйвол, заяц × кролик, не приводят к результатам, то это может происходить либо от того, что сперматозоид не достигает яйца или достигает его в поврежденном состоянии, или от того, что оказывается невозможным само гетерогенное оплодотворение, или же от того, что гибридный эмбрион погибает на самых ранних стадиях развития. Преодоление этих препятствий возможно будет конечно лишь тогда, когда для каждой гибридной комбинации будет выяснено, где именно это препятствие лежит. В частности необходимо выяснить, в каких случаях оказывается невозможным или затрудненным сам процесс гетерогенного оплодотворения.
У млекопитающих и птиц с их внутренним оплодотворением выяснение этого вопроса связано с большими трудностями и даже почти невозможно in situ. Отсюда, возникает необходимость разработать такую методику, при которой можно было бы непосредственно наблюдать оплодотворение яйца млекопитающего и птицы in vitro. Разработка этой методики в настоящее время уже началась и идет с серьезными успехами.
Помимо очевидного значения для изучения процесса гетерогенного оплодотворения методика оплодотворения in vitro, имеет и другое неменьшее значение для гибридизации, так как она позволяет получать гибридную зиготу, минуя все те трудности, которые связаны с естественным прохождением сперматозоида до яйца. Однако оплодотворение in vitro получит полное значение только в том случае, если мы разрешим вопрос о дальнейшем выращивании полученной in vitro зиготы. Так как выращивание до взрослого состояния эмбрионов млекопитающих и птиц in vitro пока совершенно невозможно, то нами намечена такая схема использования для гибридизации методики оплодотворения in vitro:
1) изъятие яиц из самки;
2) оплодотворение in vitro изъятых яиц спермой другого вида;
3) имплантация полученной зиготы, обратно в фаллопиевы трубы оперированной или другой самки.
Для судьбы метода решающее значение имеет разрешение третьего пункта схемы. В настоящее время однако Можно считать перспективы этого разрешения вполне благополучными.
Еще в 1890 г. Хипе (Неаре М.) удалось трансплантировать оплодотворенные яйца кроликов ангорской породы в трубы самкам-фландрам. Из трех опытов один дал отличные результаты, и приемная мать, будучи затем покрыта самцом-фландром, дала помет, состоявший как из крольчат-фландров, так и из двух типичных белых ангорских кроликов. Эти замечательные опыты остались если не забытыми, то неповторенными до 1930 г. Пинкус (Pincus, 1930) из трех опытов в одном получил положительный результат — самка принесла помет из трех крольчат на 31-й день. Асдель и Хэммонд поставили 21 опыт трансплантации и получили результат у трех самок, В общей сложности в этих опытах на 27 трансплантаций получено 5 удач, т. е. 18,5 %, Эти результаты приходится признать безусловно чрезвычайно интересными и обнадеживающими, учитывая неразработанность ряда вопросов, связанных, с этой операцией. Таковы например опасность ввести яйца слишком рано в матку, еще не подготовленную к восприятию яйца, опасность температурных и кислородных шоков и пр. Пинкусом было поставлено несколько опытов имплантаций яиц, оплодотворенных in vitro, с отрицательными результатами, но в силу неразработанности этой методики подобная неудача не имеет серьезного значения и требует лишь дальнейшей работы в том же направлении. Техника трансплантации описана в работе Хипе 1897 г.
Таким образом уже сейчас можно считать третье звено указанной выше схемы принципиально разрешенным, и вопрос должен итти лишь о дальнейшем. усовершенствовании гибридагогами техники трансплантации, с применением ее к яйцам, оплодотворенным in vitro, может быть о пересадках яиц одного вида в самок другого, более удобного для: работы вида, применении методики к другим видам млекопитающих и т. д. В свете задач, стоящих перед гибридизацией, разработка этой проблемы приобретает исключительный интерес.
Само оплодотворение яиц млекопитающих, in vitro в настоящее время тоже принципиально разрешено. Впервые опыты подобного рода были поставлены Шенком (Schenk, Вена) и Груздевым (Петербург, 1896), но эти опыты в настоящее время имеют лишь исторический интерес. Шенк производил оплодотворение на нагревательном столике Под микроскопом, а Груздев смешивал яйца и сперму в особой пипетке, после чего они вдувались в трубы самки кролика. Однако правильного дальнейшего развития зигот Груздев не получил, и возможно думать в свете работ Пинкуса, что наблюдавшееся Груздевым неправильное дробление носило партеногенетический характер или касалось одной лишь цитоплазмы.
В новейшее время. опыты оплодотворения яиц
кроликов in vitro были поставлены Пинкусом (Pincus, 1930), Ямане (Jamane), Красовской и Диомидовой (1934), Диомидовой и Кузнецовой, и техника его разработана уже на достаточно высоком уровне, будучи заимствованной из техники культур тканей (эксплантации). Объектом этих опытов явились яйца кроликов благодаря тому, что провоцированный характер овуляции у этого животного позволяет более или менее безошибочно определять необходимый момент операции для получения годных для опытов яиц.
Это обстоятельство важно потому, что яйца кролика пригодны для: оплодотворения лишь в течение короткого срока по выходе из яичника. В это время они одеты тонкой белковой оболочкой, которая быстро утолщается по мере продвижения яйца вниз по фаллопиевой трубе; и уже через 18 часов после коитуса яйцо становится недоступным для оплодотворения. Для получения годных к оплодотворению яиц самка покрывается вазэктомированным самцом, который, — не эякулируя в самку семени, провоцирует овуляцию. Овуляция наступает примерно через. 10 часов после коитуса, когда и производится операция, В это время яйца находятся уже в трубе. Труба осторожно вырезается, и яйца вымываются из нее раствором Паннета-Комптона или Рингера тонкой пипеткой, вводимой либо с маточного либо с фимбриального конца в зависимости от ожидаемого по времени местоположения яиц. Капли вытекающего раствора принимаются на стерилизованные слюдинки и просматриваются под микроскопом для отыскания яиц.
Когда яйцо найдено, к промывной капле прибавляются капля спермы, капля куриной или кроличьей плазмы и эмбриональный экстракт. Когда прибавленная плазма образует сгусток, фиксирующий яйцо, культура заделывается по методу висячей капли и сохраняется в термостате при 37°. Все манипуляции, начиная с промывания труб, проводятся стерильно в стерильной комнате (боксе).
Через 40–50 минут после прибавления спермы сперматозоиды проникают в яйцо. Яйца млекопитающих входят в трубу с выделившимся первым направительным тельцем. Второе деление созревания наступает естественно лишь в результате оплодотворения. В культурах in vitro это второе деление созревания наступает через 1–2 часа после прибавления спермы. Часов через 8 наступает первое дробление. Удавалось довести in vitro до третьего деления дробления, до стадии 8 бластомеров, на что требуется до 46 часов, — значительно больше, чем при развитии in situ, при котором стадия 8 бластомеров достигается уже через 24 часа.
Для интересующих нас целей — дальнейшей имплантации зиготы в трубу — достаточно убедиться лишь в наступлении гетерогенного оплодотворения, т. е. продержать яйцо in vitro лишь 1–2 часа. Дальнейшее культивирование может понадобиться лишь для того, чтобы вообще можно, было убедиться в удаче гетерогенного оплодотворения, и для цитологической проверки. Опытами Пинкуса было установлено, что в нынешних условиях культуры in vitro неоплодотворенные яйца с некоторым запозданием, но все же начинают обнаруживать признаки как бы партеногенетического дробления. Хотя деление ядра при этом еще не доказано, но по внешнему виду, принимаемому яйцом, его оказывается почти невозможно отличить от нормально дробящегося яйца. Правда, в большинстве случаев, но не всегда, образующиеся шары дробления оказываются при таком «партеногенезе» неодинаковой величины. Так как не вполне равномерное дробление происходит и при нормальном развитии яиц кролика и тем большей неравномерности возможно ожидать при гетерогенном оплодотворении, — то наступление оплодотворения для каждой гибридной комбинации должно быть проверено на окрашенных и нарезанных препаратах.
Для оплодотворения in vitro, особенно для длительных опытов, сперма должна быть получена стерильно из эпидидимиса или из vas defferens в стерильный капиляр.
Гибридизация млекопитающих in vitro в настоящее время произведена Ямане и Диомидовой и Кузнецовой (1933, неопубл.) и Красовской (1934, неопубл.). Ямане оплодотворял яйца кролика спермой крысы и даже лошади. В обоих опытах получено в надлежащем виде выделение второго полярных телец, что Ямане и считает доказательством произошедшего оплодотворения.
Целью его работы была однако не сама гибридизация, но выяснение того, является ли стимул, вызывающий второе деление созревания, специфическим, видовым, или он может быть привнесен и сперматозоидом другого вида млекопитающего. Цитологического исследования судьбы проникшего сперматозоида и участия его ядра в дальнейшем дроблении Ямане произведено не был».
Диомидова и Кузнецова оплодотворяли яйца кролика спермой морской свинки (другой
подотряд грызунов) и наблюдали несколько раз правильное дробление, в отдельных случаях до 8 бластомеров. Цитологического подтверждения наступившего оплодотворения произведено еще тоже не было, но правильность и своевременность стадий дробления делают вероятным, что гетерогенное оплодотворение действительно имело место. Наконец в только что законченных опытах Красовской цитологически показано, что при оплодотворении яиц кролика спермой крысы (другой подотряд!) в 75 % происходит настоящее дробление и образующиеся бластомеры имеют ядра. Происходит также выделение и второго направительного тельца. При оплодотворении спермой морской свинки картина оказывается уже существенно иной, — отшнуровываются лишь участки дейтоплазмы, без участия ядра, и не достаточно еще ясно-, происходит ли действительное оплодотворение. Наконец сперматозоиды собаки повидимому вовсе не проникают в яйцо кролика.
Оплодотворение in vitro является делом очень кропотливым, требующим большой тщательности и большого материала. Тем по менее тот свет, который эти работы проливают, на проблему гетерогенного оплодотворения млекопитающих, с лихвой окупает затраченный труд. Если, как и можно было ожидать (см. выше стр. 134), даже столь гетерогенное оплодотворение, как кролик × крыса (разные подотряды), протекает легко, то проблема помощи ему оказывается снятой и все внимание гибридизатора при работе с млекопитающими придется сосредоточить на проблеме осеменения, в общем гораздо более легкой и перспективной. Необходимо произвести подобные же исследования с другими видами млекопитающих и с птицами.
Глава VII. Проблема нежизнеспособности гибридов
Нежизнеспособность гибридов
После того как гетерогенное оплодотворение достигнуто и преодолены стоящие на этом пути препятствия, гибридизатор встречается с новыми препятствиями — в виде неспособное и гибридного эмбриона к нормальному развитию в той или иной степени; в той или иной форме. Наша работа упирается в явление нежизнеспособности гибридов, при
настоящем уровне наук ставящее наиболее трудно преодолимые препятствия. Однако, как увидим ниже, и здесь возможно, по крайней мере в некоторых случаях, вполне плодотворно ставить вопрос
о преодолении нежизнеспособности гибридов, что естественно делает необходимым анализ степеней и форм нежизнеспособности и выяснение их причин. В этом анализе мы должны прежде всего различить две проблемы: первая, более общая, связана с жизнеспособностью или нежизнеспособностью первого поколения (F
1) гибридов, возникающей непосредственно. в результате гетерогенного оплодотворения; вторая связана с такими явлениями, когда, с одной стороны, от вполне жизнеспособных гибридов F
1 получается в той или иной степени нежизнеспособное дальнейшее потомство, или, наоборот, от мало жизнеспособного F
1 в следующих поколениях возникает потомство, более жизнеспособное. Обе эти стороны вопроса очевидно имеют для гибридизации и ее практического использования самое серьезное значение.
Как увидим ниже, ряд авторов связывает общим объяснением как. явление нежизнеспособности гибридов, так и явление повышенной: их жизнеспособности, — так называемый гетерозис. Если однако некоторые стороны теории гетерозиса и нежизнеспособности имеют родство, то во всяком случае рассматривать оба вопроса отдельно вполне возможно. И тем более — возможно, что в работе гибридизатора явление нежизнеспособности гибридов играет роль препятствия, которое нужно преодолевать, тогда как гетерозис является чрезвычайно ценным свойством, вопрос о преодолении которого может ставиться, лишь в исключительных случаях, когда он действительно переходит-в свою противоположность, например осложняя роды слишком — сильным развитием плода:
Формы и степени нежизнеспособности гибридов чрезвычайно разнообразны. Гибрид может, быть вполне жизнеспособным, а в случае наличия гетерозиса даже более жизнеспособным, чем чистые виды. В других случаях гибрид может оказаться с более или менее ослабленной жизнеспособностью, которая восстанавливается до нормы лишь при специальном подборе внешних условии (см. ниже случай гибридов дрозофил). Далее мы встречаемся с так называемым явлением сублетальности, при котором, как у некоторых гибридов бабочек, развитие идет вполне нормально, а затем на стадии куколки останавливается. В еще более тяжелых случаях гибрид гибнет еще раньше благодаря дисгармоничному, или прямо тератологическому развитию, как например в отдаленных скрещиваниях рыб. Эго отмирание может опускаться к еще более ранним стадиям, и наконец гибрид может погибать уже при самом начале дробления.
Чрезвычайная распространенность явлений нежизнеспособности гибридов, особенно при гибридизации более отдаленной, и «абсолютность» нежизнеспособности при ультрагибридизации привели ж попыткам широкого и несколько убаюкивающего обобщения в этом допросе, утверждавшего, что нежизнеспособность гибридов пропорциональна систематическому удалению друг от друга родительских форм, и сводившего эту нежизнеспособность непосредственно к степени биохимического различия между скрещиваемыми видами. Это обобщение включало в единую схему как явление нежизнеспособности гибридов, так и явление ослабления жизнеспособности, наблюдающееся нередко при инбридинге, так наконец и явление гетерозиса (W. Schultz, 1912). Сторонники этого обобщения утверждают, что для нормального развития организма необходима некоторая степень различия между «наследственными плазмами» отца и матери. При полном сходстве этих плазм, как при размножении в тесном родстве, а тем более при самооплодотворении, жизнеспособность лежит ниже оптимума, в иных случаях довольно низко. При некотором различии, сопровождающемся например по Кэстлу (Castle) слабым «взаимным отравлением» встретившихся зародышевых плазм, развитие усиливается, достигает при известном оптимальном «взаимном отравлении» степени пышного развития — гетерозиса, а при дальнейших степенях различия и соответственно при все усиливающихся степенях отравления приводит снова к понижению жизнеспособности вплоть до полной нежизнеспособности при отдаленной гибридизации.
Исходя из этой «биохимической» точки зрения, В. Шульц в ряде работ пытался сопоставить явление жизнеспособности при гибридизации с явлениями, наблюдаемыми при трансплантациях. Как известно, судьба трансплантата сильно связана со степенью родства донора и рецептора и обычно трансплантация удается лишь у более близких форм, особенно у высших животных. В опытах же с видами удаленными трансплантат всегда более или менее быстро отмирает. Поэтому Шульц счел возможным не только сопоставить три параллельные ряда: 1) систематическое родство, 2) трудность трансплантации и 3) трудность гибридизации, — но и свести все эти три ряда — к степени биохимического различия. Отмирание трансплантата и нежизнеспособность гибридного эмбриона в этой концепции — результат биохимической несовместимости соединяемых компонентов, соединяемых либо хирургическим либо гибридизационным методом,
Подобная чисто биохимическая упрощенная схема, смешивающая в единый ряд самые различные в качественном отношении явления, совершенно неудовлетворительна несмотря на наличие в ней и некоторых здоровых элементов. Уже чисто биохимическая трактовка трансплантации не выдерживает экспериментальной проверки.
Некоторые причины того, что ткани чужого вида в случае теплокровных животных не в состоянии развиваться на любом хозяине, были выяснены замечательными работами Мерфи (Murphy, 1912, 1914, 1915), Мерфи нашел, что можно с большим успехом производить трансплантацию любой ткани чужого вида на зародыш курицы в ранних стадиях его развития. Даже столь чуждые ткани, как человеческие, пересаженные на такой зародыш, быстро и хорошо растут. Отсюда ясно, что на такой ранней стадии развития зародыш курицы еще не реагирует на пересаженную чужую ткань. Отсутствие реакции наблюдается приблизительно до 19—21-го дня жизни эмбриона; с этого же момента не только прекращается рост трансплантата, но последний или отпадает сам собой или же подвергается разрушению.
Мерфи заметил, что критический период совпадает с развитием селезенки и лимфатической ткани зародыша и что так называемые лимфоциты, развивающиеся в лимфоидной ткани, скопляются в большом количестве по краям трансплантата. Мерфи показал, что эти лимфоциты и разрушают чужую ткань. Мысль свою он подтвердил двумя опытами.
Во-первых, он показал, что если пересадить в зародыш небольшой кусочек селезенки взрослого цыпленка, то зародыш теряет свою толерантность по отношению к чужеродным трансплантатам.
Второе доказательство состояло в следующем. Известно, что лимфоциты животного могут быть разрушены рентгеновскими лучами. Можно было ожидать, что рентгенизованное животное потеряет свойственную ему способность сопротивляться трансплантатам. Мерфи подтвердил это опытом. На взрослых крысах, лимфоциты которых были разрушены рентгеном, наблюдался прекрасный рост тканей, принадлежавших другим видам. Наоборот, увеличивая количество лимфоцитов путем умеренной рентгенизации, можно добиться усиления иммунитета по отношению к гетеротрансплаптатам.
Из этих интересных опытов следует, что причины отмирания трансплантата хотя и имеют иммунологический характер, по связаны не непосредственно с «биохимическим различием)) соединяемых видов, а с характером деятельности определенных органов и систем. До развития этих органов несмотря на чрезвычайную «биохимическую удаленность)) частей курицы и человека они могут благополучно развиваться рядом. И лишь когда в эмбрионе курицы начинает функционировать определенный типично-видовой орган, продукция этого органа оказывается гибельной для трансплантата.
Ничего подобного в гибридном эмбрионе мы не имеем. Когда мы получаем гибриды
А ×
В, то гибридными оказываются все его клетки и все его органы. У гибрида нет например селезенки одного вида и тканей другого вида., которые могли бы оказаться в положении антагонистов. Наоборот, селезенка гибрида будет также гибридной, имеющей функцию и биохимию, более или менее промежуточную между видами
А и
В. И объекты воздействия этого органа, остальные ткани будут тоже структуры
АВ, и нет никаких оснований полагать, что: между селезенкой
АВ и другими тканями
АВ возникнут отношения, сколько-нибудь подобные тем, которые существуют между селезенкой
А и трансплантатом
В. Скорее, наоборот, с этой точки зрения можно было бы ожидать, что жизнеспособность даже самых отдаленных гибридов будет более или менее нормальной.
Эти соображения конечно не исключают того, что биохимические отношения между соединяемыми в процессе гибридизации видами должны иметь важное значение и могут играть роль и при определении жизнеспособности. Однако уже-решительные, нередко наблюдающиеся различия в результатах реципрокных скрещиваний говорят против возможности какого-либо упрощенного толкования этих, явлений. Данные Мерфи о возможности воспитания трансплантата Млекопитающего на эмбрионе курицы говорят за то, что «биохимическая толерантность» клеток различного генотипа достигает чрезвычайной степени.
Как связана жизнеспособность гибридов со степенью родства скрещиваемых видов?
По этому вопросу существует интересное разногласие между авторами. Что некоторая степень связи здесь имеется, вообще говоря несомненно. Но какова эта степень, может ли рассмотрение родства служить нам руководством в гибридагогической работе — это гораздо более сложный вопрос, и решать его повидимому приходится отрицательно.
Решительным защитником тезиса о зависимости жизнеспособности гибридов от степени родства родителей выступил Менкгауз: «удача гибридизации, т. е. стадия, до которой данный гибрид развивается, стоит в корреляции со степенью родства» (Менкгауз, 1910).
Против этого положения можно выдвинуть следующие факты.
1. Реципрокные скрещивания дают часто различные результаты в смысле жизнеспособности несмотря на то, что родство родителей одинаково.
2. Видовые скрещивания часто дают нежизнеспособное потомство, в то время как отдаленные дают гораздо более жизнеспособное.
Скрещивания Fundulus diaphanus × Cyprinodon variegatus в направлении С♂ × ♀F погибают в начале гаструляции, a F♂ × ♀C проходят стадию гаструляции благополучно. Различна жизнеспособность в скрещивании ежей St. purpuratus и S. franciscanus и др.
Шесть межродовых скрещиваний (три вида Fundulus × Суprinodon) идут гораздо хуже несмотря на принадлежность обоих родов к сем. Cyprinodontidae, чем другие гораздо более далекие скрещивания с тем же Fundulus. Так, F. heteroclitus (отряд Haplomi) × Gasterosteus bispinosus (отр. Hemibranchiae) дают эмбрионов, у которых хотя зародышевый диск и развит лишь до
1/
3 вокруг желтка, но «эмбрион короткий и вполне сформирован». Точно так же скрещивания того же F. heteroclitus с Menidia notata (представители различных отрядов) дают гибрида, описанного Менкгаузом, как достаточно диференцированного, чтобы обнаруживать зрительный пузырек. Много подобных же примеров можно найти у самого Менкгауза. В опытах Г. и П. Гертвигов среди многих нежизнеспособных гетерогенных комбинаций встречается одна — Box♂ × ♀Crenuabrus, давшая, почти совершенно нормальных гибридных мальков, которые однако не были партеногенетическими, что доказывается величиной их ядер и тем, что они развивают хроматофоры типичной для отцовского вида формы.
Эти факты — что важно отметить — решительно говорят против — того, что причина нежизнеспособности гибридов лежит в сфере, близкой к токсическим явлениям при сливании кровей различных видов, — в сфере, связанной с химическими различиями организмов. Эти различия, поскольку они улавливаются например преципитиновой реакцией, дают всегда весьма правильный ряд, отвечающий филогенетическому отстоянию, и также не дают резких сдвигов у близких видов и у реципрокных сочетаний. Существуют поэтому несомненна другие более сильные факторы, определяющие жизнеспособность гибрида, и эти факторы несомненно очень конкретны.
Классификация явлений нежизнеспособности
Классификация явлений нежизнеспособности гибридов, или, что то же, патологии гибридов, может быть дана в следующей предварительной форме:
1. Патология зиготы, связанная с неправильностями гибридного оплодотворения и в первую очередь с нередко наблюдающейся полиспермией.
2. Патология ранних стадий эмбриогенеза, связанная с патологией гибридной клетки, в свою очередь связанной с неправильностями гибридных митозов, нарушением клеточного деления, отмиранием, хромосом и, может быть другими физиологическими явлениями пока не проанализированного характера.
3. Патология поздних стадий, связанная с нарушением взаимной приспособленности органов эмбрионов, их непропорциональным или тератологическим развитием, нарушением функции важных органов и т. п.
4.. Патология родов и вылупления, связанная с несоответствием анатомии и физиологии гибридного эмбриона и материнского организма (у живородящих) или его производных (строением скорлупы яйца и пр.). Эти патологические явления могут начинаться и на более ранних стадиях эмбрионального развития.
Полиспермия как причина гибели гибридных зигот
В качестве одной из причин ранней гибели гибридных зигот следует, отметить прежде всего полиспермию — фактор, не связанный с генотипом. Возникновение полиспермии, т. е. оплодотворение яйца многими сперматозоидами, начиная с двух, наблюдали у гибридов уже Менкгауз (1904), Купельвизер (1909) и др.
Проникновение нескольких сперматозоидов, каждый из которых приносит о собой активный элемент — центросому, приводит к тому, что в яйце возникают своеобразные многополюсные митозы (рис. 45).
Уже при двух центросомах возникает не одно нормальное веретено, а шесть, так как каждая центросома, разделившись на две, дает веретено между своими дочерними центросомами, да кроме того все 4 дочерние центросомы еще дополнительно попарно образуют веретена, т. е. образуют тетраэдр, ограниченный 6 веретенами. Хромосомы ядра оплодотворения (у Купельвизера даже только ядра яйца) распределяются при этом самым разнообразным способом по этим многим веретенам. Если такие четырехполюсные клетки закончили бы деление, то образовалось бы сразу 4 клетки с совершенно различными наборами хромосом, что привело бы к гибели эти клетки. Но и само дробление при полиспермии идет неправильно и эмбрион погибает.
 Рис. 45. Многополюсные митозы при полиспермии (по Купельвизеру).
Рис. 45. Многополюсные митозы при полиспермии (по Купельвизеру).
Причиной возникновения полиспермии при гибридизации в случае Echinus (морской еж) × Mytilus (молюск) выяснил Купельвизер. Она состоит в том, что при проникновении первого сперматозоида в яйцо последнее не отвечает на него образованием оболочки, — предохраняющей от проникновения дальнейших сперматозоидов, и они могут внедриться один за другим вплоть до нескольких десятков.
Вариируя условия оплодотворения, полиспермии можно не допустить. Так, Купельвизер выяснил, что срок. «экспозиции» т. е. срок от момента допуска сперматозоидов к яйцам до момента отмывки, имеет большое значение, и при слишком длительной экспозиции почти все яйца оказываются полиспермными и погибают, не давая дробления. Повышение концентрации спермы имеет обратное — значение — под влиянием концентрации спермы может быть вызвано образование оболочки (и даже при чересчур сильной концентрации так быстро, что не успевает произойти оплодотворение). Леб применяя свою методику в опыте со спермой Chlorostoma, повидимому совершенно не допускал полиспермии.
Для наших целей чрезвычайно ценны наблюдения Менкгауза (1904) над различной частотой полиспермии в реципрокных скрещиваниях. Производя у рыб оплодотворение яиц Fundulus спермой Mcnidia, Менкгауз наблюдал в 50 % диспермню. В реципрокном скрещивании, наоборот, только в немногих случаях была ди- или полиспермия. Иными словами, жизнеспособные зиготы образовывались, в реципрокных скрещиваниях в совершенно различном проценте (оставляя пока вопрос об их дальнейшей жизнеспособности). В противоположность тому, что для удачи скрещивания казалось бы желательной большая легкость проникновения сперматозоида в яйцо, здесь мы видим обратную картину: там, где. проникновение сперматозоидов в яйцо идет беспрепятственно, зиготы в массе оказываются нежизнеспособными…
Еще более показательны в этом отношении данные Ньюмана (1914). Им были поставлены упомянутые реципрокные скрещивания! между тремя видами Fundulus и Cyprinodon. При реципрокных скрещиваниях в пределах Fundulus и при оплодотворении яиц Cyprinodon всеми тремя видами Fundulus полиспермия никогда не наблюдалась. Наоборот, при оплодотворении яиц Fundulus спермой Cyprinodon закономерно возникала в значительном проценте полиспермия, и это повторялось в скрещиваниях с каждым из трех видов.
Патология гибридов морских ежей
Очень ценные материалы, освещающие природу патологических явлений, возникающих при гибридизации, дают нам исследования Бальцера (Baltzer, 1910) над развитием гибридных эмбрионов морских ежей. Производя различные межродовые скрещивания неаполитанских ежей, он тщательно изучил отмечавшиеся уже прежними исследователями картины как бы внезапных заболеваний гибридных личинок. При этом им было обнаружено два совершенно различных По природе случая, которые мы и рассмотрим отдельно.
I. Sphaerechinus-тип. При оплодотворении яиц Strongylocentrotus lividus, Echinus microtuberculatus и Arbacia pustulosa — всех, каких только мог испробовать Бальцер, — спермой ежа Sphaerechinus granularis всегда как общее правило на самых ранних стадиях образования бластулы гибридные эмбрионы обнаруживали «заболевание» со следующими признаками. Как только начиналось образование бластоцеля, т. е. через 13–20 часов по оплодотворении, в полость бластоцеля начинали выходить многочисленные клетки. В это время эмбрионы находятся еще в оболочке, и нормальное образование мезенхимы, сопровождающееся тоже проникновением клеток в бластоцель, начинается лишь гораздо позднее, часов через 24–26 по оплодотворении. Таким образом описываемое раннее внедрение клеток в бластоцель не имеет никакого отношения к нормальному процессу образования мезенхимы. Чем больше становится бластоцель, тем гуще он набивается кучами клеток, подвергающихся частично распаду. Личинки становятся болезненными, непрозрачными и массами опускаются на дно. Рис. 46 представляет две таких больных бластулы в возрасте 12 часов и 25 часов после оплодотворения. На правом рисунке видны помимо целых и распадающихся клеток внутри бластоцеля еще патологические клетки с гигантскими ядрами в стенке бластулы.
 Рис. 46. Больные бластулы Strongylorentrotus ♂ × ♀ Sphaerechinus, 12 и 25 час. (по Бальцеру).
Рис. 46. Больные бластулы Strongylorentrotus ♂ × ♀ Sphaerechinus, 12 и 25 час. (по Бальцеру).
Это болезненное состояние может длиться несколько дней, в течение которых развитие останавливается. Многие личинки на этом и кончают свою жизнь. Но некоторые переболевают и снова начинают развиваться дальше, давая плютеусов с опозданием на несколько дней. В редких случаях течение болезни оказывается короче, длится около суток. Вернон (Vernon, 1900) отметил даже, что в июле и августе значительное число эмбрионов развивается до плютеуса.
Такова картина этого тяжелого «гибридного» заболевания. Бальцер вскрыл его этиологию. Оказалось, что оно имеет прямое отношение к происходящей у всех гибридов полученных от Sphaerechinus
♂ элиминации хромосом, приносимых сперматозоидом. Эта элиминация будет описана в другом месте. Здесь однако отметим элементы, важные для нас. Элиминация происходит здесь уже в первом-втором делении дробления, когда обнаруживается неспособность хромосом Sphaerechinus проделывать правильный митоз вместе с хромосомами яйца. Хромосомы Sphaerechinus отрываются от веретена и остаются целиком или группами в некоторых бластомерах. При этом однако они не умирают тотчас же, а оказываются способными удваиваться, размножаться, в результате чего в некоторых клетках возникают совершенно неестественные скопления хромосом. На рис. 47 виден налево отрыв группы хромосом от веретена, направо — патологически размножившаяся куча. элиминированных хромосом. Эти скопления хромосом и может быть продукты их распада повидимому и являются источником заболевания. Так, по дальнейшим исследованиям Бальцера видно, что именно эти части клеток и начинают выбрасываться в бластоцель (ср. его рис. 7, 8), где и происходит их постепенный распад. Этот процесс является своего рода оздоровительным, так как в результате его ядра освобождаются от всех или почти всех чужих хромосом, клетки освобождаются от патологических включений, и эмбрион становится гаплоидом материнского вида, после чего и начинает развитие дальше, давая в результате плютеуса чисто материнского типа с частыми однако аномалиями, наличие которых после пережитой болезни конечно вполне естественно.
 Рис. 47. Размножение элиминированных хромосом в клетках бластул Strongylocentrotus ♀ × ♂ Sphaerechinus.
Рис. 47. Размножение элиминированных хромосом в клетках бластул Strongylocentrotus ♀ × ♂ Sphaerechinus.
Таким образом причиной этого заболевания является неправильность механизма митоза, неспособность хромосом Sphaerechinus проходить правильно митозы в обстановке чужих яиц. О каком-либо физиологическом антагонизме здесь повидимому речи не идет, так как в реципрокных сочетаниях хромосомы обоих видов благополучно уживаются, давая гибридных личинок промежуточного типа.
II. Arbacia-тип. В том случае, когда яйца ежей Strongylocentrotus, Sphaerechinus и Echinus оплодотворяются спермой ежа Arbacia pustulosa, также, с полной закономерностью возникают патологические явления у гибридов. Здесь также возникает массовое заболевание, отличающееся несколько от предыдущего случая своей внезапностью. Раннее развитие идет совершенно нормально (рис. 48), бластулы начинают оживленно двигаться, после чего внезапно обнаруживается как бы массовое отравление и личинки оказываются в состоянии тяжелого заболевания. Начало болезни совпадает со стадией бластулы, наступая однако позже, чем и Sphaerechinus-типе, часов через 26 после оплодотворения, при начале образования мезенхимы. Заболевшие бластулы становятся непрозрачными, и их бластоцель опять наполняются кучами дегенерирующих клеток (рис. 49). Лишь немногие оправляются от заболевания и дают матроклинных плютеусов.
 Рис. 48. Нормальное развитие Strongylorentrotus × Arbacia до гаструлы.
1 — гаструла гибрида, 2 — его ядра, 3 — для сравнения чистый Strongylocentrotus и его ядра (по Бальцеру).
Рис. 48. Нормальное развитие Strongylorentrotus × Arbacia до гаструлы.
1 — гаструла гибрида, 2 — его ядра, 3 — для сравнения чистый Strongylocentrotus и его ядра (по Бальцеру).
 Рис. 49. Развитие гибрида Strongylocentrotus ♀ × ♂ Arbacia.
1 — 22 часа; 2 — 26 1/2 часов, начало заболевания; 3 — 47 часов, сильное заболевание (по Бальцеру).
Рис. 49. Развитие гибрида Strongylocentrotus ♀ × ♂ Arbacia.
1 — 22 часа; 2 — 26 1/2 часов, начало заболевания; 3 — 47 часов, сильное заболевание (по Бальцеру).
Этиология этого гибридного заболевания, которое можно назвать
арбацитом в противоположность предыдущему
сферэхиниту, также до некоторой степени разъяснена.
Гистологическое исследование показало, что заболевание связано опять-таки с элиминацией
:хромосом, повидимому хромосом Arbacia. Эта элиминация идет здесь однако совершенно иначе— до заболевания материнские и- отцовские хромосомы нормально участвуют в митозах, но в начале заболевания ядра принимают болезненный вид и выбрасывают часть хроматина в плазму: ядра становятся приблизительно вдвое меньше, а выброшенные части ядер вместе с болезненного вида протоплазмой переполняют бластоцель, что затрудняет гаструляцию. Рис. 50 дает картину больной гаструлы.
 Рис. 50. Гаструла Strongylocentrotus × Arbacia в возрасте около 30 часов (по Бальцеру); больные ядра.
Рис. 50. Гаструла Strongylocentrotus × Arbacia в возрасте около 30 часов (по Бальцеру); больные ядра.
Во многих местах видны сплошь окрасившиеся ядра, а в нижней части левого рисунка видна довольно вероятная картина выбрасывания хроматиновых кусков из ядер. Правый рисунок представляет картину при большем увеличении.
Причина «арбацита» несомненно иная, чем, «сферэхинита», так как здесь указаний на какое-либо расстройство митозов нет. В соответствии с взглядами Бовери Бальцер дает такое истолкование явлений: первые стадии дробления проходят под влиянием «общих» воздействий хромосом, вероятно одинаковых как у хромосом Strongylocentrotus и др., так и у Arbacia. А так как и те и другие хромосомы способны делиться в одинаковом темпе, то до бластулы дело идет благополучно. Но здесь начинается уже функционирование специфических видовых особенностей хромосом, и с этого момента обнаруживается антагонизм между хромосомами Arbacia и окружающей их плазмой Strongylocentrotus (или Echinus), приводящей к смерти хромосомы Arbacia. Убитые или пораженные, они и выбрасываются ядром в порядке самооздоровления. Если же это самооздоровление почему-либо не может осуществиться, эмбрион погибает.
Оба эти заболевания гибридных эмбрионов имеют то общее, что они не связаны с неправильным и несогласованным развитием
органов, которое естественно ожидать у гибридов, вынужденных развиваться по некоторой равнодействующей между двумя гармоничными родительскими типами развития, примером чего невидимому могут служить некоторые гибриды Fundulus (например F. diaphanus ♀ × ♂ F. majalis), у которых не устанавливается желточное кровообращение. Здесь же патологические факторы лежат глубже, еще в жизни самой клетки.
Расстройства клеточного деления в результате гибридизации
Для понимания природы летальных явлений, возникающих в результате гибридизации интересные: данные доставляют наблюдения М. Моррис (1914) над гибридами рыб Fundulus × Ctenolabrus, Как было ужа изложено, процесс, оплодотворения здесь происходит вполне нормально, и нормально же начинается дробление, в котором отчетливо участвуют хромосомы обоих видов. Эта правильность продолжается однако сравнительно, недолго, и уже через 12 часов после оплодотворения начинают замечаться ненормальности, выражающиеся в том, что некоторые клетки видимо утрачивают способность к делению, в результате чего в бластодерме возникают гигантские клетки с неправильными ядрами (рис. 51).
 Рис. 51. Расстройство клеточного деления у гибридов рыб Fundulus х Ctenolabrus (по Моррис).
Рис. 51. Расстройство клеточного деления у гибридов рыб Fundulus х Ctenolabrus (по Моррис).
Возникновение гигантских клеток является несомненно результатом гибридизации, так как у контрольных оплодотворенных спермой своего вида яиц развитие идет совершенно нормально. Эти неправильности деления клеток становятся повидимому причиной массового отмирания эмбрионов и уродств тех из них, которые продолжают еще некоторое время свое развитие.
Гистологические ненормальности гибридных эмбрионов наблюдали у рыб также Г. и П. Гертвиги (1914).
Здесь в момент массового отмирания бластул наблюдалось ненормальное увеличение размеров ядер, но без нарушения правильности клеточного деления.
О возможности, борьбы с этими патологическими явлениями у гибридов вопрос еще но ставился. Однако на возможность такой борьбы имеется серьезное указание прежде всего в виде чрезвычайной вариации степени заболевания эмбрионов. Почти всегда часть эмбрионов быстро погибает, другая часть, остановившись в своем развитии, долгое время сохраняет жизнь. Наконец некоторые, переболев, развиваются дальше. Правда, не все они с точки зрения гибридизации сохраняют ценность, ибо те из выживших, которые нацело освободились от отцовских хромосом, уже перестают быть гибридами, совершают процесс
дегибридизации.
Однако исследования Бальцера показали, что в случае как сферэхинита, так и арбацита элиминация хромосом происходит неполно, и в случае Sphaerechinus-отца в плютеусах сохраняется одна из его хромосом, а во втором случае — 2–3 хромосомы. Но и здесь вероятно наличие вариаций, и Бальцер приводит несколько исключений, не вполне правда выясненных, когда личинки сохраняли нормальную или почти нормальную величину ядер и давали плютеусов со столь измененным скелетом, что в них можно было допустить сохранение хромосом Arbacia в полном или более или менее полном количестве.
Здесь представляется благодарное поле для исследований по линии различных изменений внешней среды, иммунологических и других воздействий на скрещиваемые организмы и т. д., для того чтобы отыскать возможности передвинуть вариацию в благоприятную сторону. За возможность этого говорит и весьма различный ход отдельных скрещиваний. Если взять например приведенные Бальцером протоколы опытов Strong. ♀ × ♂ Arbacia
А, В, С, D, то в опыте
D например на 6-й день большинство зародышей погибло, а выжившие не развились дальше стадии гаструлы без скелета и погибли на следующий день, тогда как в опыте
С уже на 3-й день у некоторых началась гаструляция, отмечены стадии призмы и к вечеру отдельные плютеусы.
Не менее интересны были бы и опыты «исправления» неправильных митозов в случае сферэхинита путем изменения. внешними факторами темпа дробления, например охлаждением, прекращением дыхания цианом и др.
Патология гибрида как целого
Если до сего времени шла речь о патологических явлениях, связанных с нарушением нормальной физиологии клеток, составляющих гибридный организм и приводящих его к гибели, то в этом разделе мы встречаемся с совершенно новым классом явлений, когда первоначально организм составляется из вполне здоровых и жизнеспособных элементов и приходит к гибели лишь благодаря несогласованному развитию отдельных
органов. Конечно далеко не всегда легко решить, с каким классом явлений мы имеем дело, и ответить на вопрос о том, являются ли источником нежизнеспособности отдельные дефектные органы, дисгармоничные соотношения отдельных органов или же более глубокая нежизнеспособность самих клеток благодаря например несоответствию физиологии хромосом с цитоплазмой или хромосом разных видов друг с другом. В настоящее время мы имеем указание на то, что кроме разобранных выше примеров клеточной патологии имеются и другие не менее тонкие источники нежизнеспособности, связанные уже с организмом как целым. Тот же Бальцер нашел (Baltzer, 1930), что при опытах с мерогонией у тритонов отцовское ядро Triton cristatus в материнской цитоплазме T. taeniatus может действовать лишь ограниченное время. Все мерогоны погибают на стадии эмбрионов с глазными пузырями, когда ядра мезенхимы головы и склеротомов начинают становиться пирнотическими. Продолжая анализ этого явления, Хадорн (Hadorn, 1932) трансплантировал части этих мерогонов на ранних стадиях ортотопно в зародышей taeniatus и имел возможность выращивать эпидермис, мозг, хорду и сомиты, имеющие в клетках чистые cristatus-ядра, далеко за критическую, указанную выше стадию. Но если трансплантировалась мезенхима головы, то наступала дегенерация клеток. И пожалуй наиболее интересно, что если ткани мерогона, пересаженные в головную часть эмбриона, детерминировались здесь в мезенхиму, то дегенерация наступала также. Эти чрезвычайно интересные опыты учат нас, что летальное несоответствие ядра ii цитоплазмы может не быть абсолютным для всех тканей данной гибридной комбинации и возникать лишь тогда, когда в процессе эмбриогенеза некоторые ткани подвергаются той или иной спецификации. Как увидим ниже, это обстоятельство открывает перед нами некоторые перспективы преодоления нежизнеспособности гибридов.
Аналогичные факты получил Битинский-Зальц (Bytinski-Salz, 1933) у гибридов бабочек. В скрещивании бражников Deilephila galii ♂ × ♀ D. euphorbiae гибридные самки оказываются неспособными преодолеть стадию куколочного развития и после нескольких зимовок погибают. Путем трансплантаций зачатков крыльев и яичников из этих сублетальных куколок в нормально развивающиеся куколки Битинский-Зальц показал, что зачатки эти в нормальной куколке уже вполне способны к нормальному развитию и что следовательно какой-либо летальной дисгармонии в самих этих тканях не имеется. Подробнее об этих опытах будет сказано еще в следующей главе.
Хорошие примеры такой «вторичной» патологии гибридных эмбрионов мы находим в ряде экспериментальных работ Ньюмана. Рыбы вообще являются очень благоприятным объектом для изучения этого вопроса, поскольку здесь может быть, с одной стороны, широко применено искусственное осеменение для массового получения гибридов, а с другой стороны— эмбриональное развитие может легко наблюдаться. В лаборатории морской станции в Вудс-Голе Ньюман произвел до 80 гибридных сочетаний морских и частично пресноводных рыб, часто весьма отдаленных систематически, и изучил характер их эмбриогенеза. Ряд поучительных данных мы находим у него и по проблеме нежизнеспособности.
В отдаленном скрещивании Fundulus heteroclitus с Menidiu возникали например совершенно уродливые эмбрионы благодаря необычайному разрастанию сердечной оболочки (перикардия). Она занимала большую часть яйца, оттесняя эмбрион, и само сердце оказывалось далеко отдаленным от остального эмбриона, соединяясь с ним лишь длинной нитевидной связью (рис. 52, вверху). При всем том сердце билось, ткани его, как и остального эмбриона, очевидно были жизнеспособными. Но очевидно, что подобное непропорциональное развитие сердечной сумки и странное положение сердца уже не могли обеспечить дальнейшего правильного функционирования организма как целого, и он погибал.
 Рис. 52. Уродливые эмбрионы рыб (по Ньюману).
h — сердце, р.с. — перикардий, Fht, M.t и P.t. — пигментные клетки различных родительских типов, подтверждающие гибридность эмбрионов.
Рис. 52. Уродливые эмбрионы рыб (по Ньюману).
h — сердце, р.с. — перикардий, Fht, M.t и P.t. — пигментные клетки различных родительских типов, подтверждающие гибридность эмбрионов.
В другом случае при скрещивании того же Fundulus heterpclitus с Poronothus triacantus эмбрионы возникали с систематическим уродством в виде недоразвития передней части головы, кпереди от глаз (рис. 52, внизу). Остальные части эмбриона, наоборот, развивались более, или менее нормально. Ясно, что тератологическое развитие головы не могло привести к жизнеспособному организму.
Поучительно, что характер уродств и ненормальностей в различных скрещиваниях был различен. Часть этих ненормальностей, правда, была не очень специфичной: например раздвоение хвоста (рис. 52, посредине) и тому подобные уклонения встречались в разных комбинациях видов. Но ряд уродств вроде изложенных выше характеризовал собой определенные гибридные комбинации, указывая на то, что они возникают как результат полученной комбинации наследственных элементов (не только генов, но может быть и строения всего яйца).
Очень поучительно изученное С. Г. Крыжановским (Москва; не опубликовано) эмбриональное развитие гибридов карпа и леща. Эти гибриды оказываются тоже нежизнеспособными. Крыжановский изучил анатомию эмбрионов, обнаружив у них например чрезвычайно своеобразное развитие кровеносной системы в области жаберного аппарата (о чем подробнее см. главу IX). У обоих родительских видов эмбриональное кровообращение, связанное с желточным пузырем, происходит различно благодаря различному расположению и развитию подкишечной вены и пр. «Сложенная» из двух таких различных компонентов кровеносная система эмбриона является новообразованием, отличается от обоих родительских видов и носит весьма аномальные черты, например аномально развита подкишечная вена. Может быть в связи с этим и сердце оказывается развитым слабее нормы, более вяло пульсирует и очевидно перестает справляться со своей задачей. У эмбрионов развивается околосердечная водянка, увеличивается перикардий, и эмбрионы гибнут (рис. 53). Этот пример особенно показателен в том отношении, какой степени конкретности может достигнуть анализ причин нежизнеспособности гибридов, коль скоро мы перестанем удовлетворяться голой формулой «биохимического различия» между скрещиваемыми видами.
 Рис. 53. Околосердечная водянка у гибрида карась × лещ (по Крыжановскому).
Рис. 53. Околосердечная водянка у гибрида карась × лещ (по Крыжановскому).
Неправильное развитие эмбрионов наблюдалось и при гибридизации жаб с лягушками — форм, при внешнем сходстве сильно различающихся анатомически. К сожалению детальных анатомических исследований этих нежизнеспособных эмбрионов, подобных последованию Крыжановского, произведено не было.
Относительно случаев нежизнеспособности гибридных эмбрионов млекопитающих определенных данных тоже еще нет кроме отрывочных указаний. Так, при попытках гибридизации домашних коз с гривистым бараном дважды получались мертвые эмбрионы — Мильн-Эдвардсом в Париже и в Аскании-Нова (1931). Здесь полуразвитой эмбрион подвергся резорбции и был абортирован, однако уже в таком виде, что не только не было возможности изучить его анатомически, но и подтвердить его гибридное происхождение.
_______
При обсуждении проблемы жизнеспособности гибридов приходится больше задумываться не столько над их нежизнеспособностью, сколько над их жизнеспособностью. В самом деле мы привыкли считать организм определенного вида результатом длительной приспособительной эволюции, в результате которой естественным отбором отметались даже сравнительно очень незначительные отклонения, так как только этим объяснялись малая изменчивость представителей каждого вида, устойчивость его характерных признаков и свойств. Если мы обсуждаем например особенности ибиса и колпицы, то у каждого из этих видов мы находим множество специфических приспособительных признаков хотя бы в виде резко различных и характерных форм их клювов, оперения и т. д. Когда мы производим гибридизацию и соединяем в единый генотип два различных генотипа, эмбриональное развитие должно направиться по некоторой равнодействующей между обоими видами, что и реализуется в виде промежуточной или хотя бы мозаичной формы гибрида. Спрашивается, как получается то, что эта равнодействующая, нарушающая тонкое приспособительное строение обоих родов, взятых для скрещивания, все-все-такиможет привести к жизнеспособному организму? Факт полной жизнеспособности многих видовых и даже родовых гибридов заставляет признавать или второстепенность различия между видами даже тогда, когда они носят очень резкий характер, как например ибис и колпица, фазан и кракс, зебу и як, либо высокую регуляторную способность организма к согласованию темпов и характера развития различных его органов — способность, оказывающуюся недостаточной лишь при чрезвычайной степени несходства скрещиваемых видов. Несомненно, что для гибридизации эта проблема имеет крупнейшее значение, и изучение эмбриогенеза с этой точки зрения должно привлечь к себе внимание гибридагогов как морфологов, так и физиологов.
Патология гибридной беременности
В некоторых случаях констатированы патологические явления, могущие приводить к гибели гибридов благодаря ненормальным взаимоотношениям, устанавливающимся между гибридным эмбрионом и родительским организмом: у млекопитающих непосредственно с беременной самкой, вынашивающей гибрида, у птиц, рыб и др. — между эмбрионом и окружающими его яйцевыми оболочками и пр. Эта категория патологических явлений, будучи изучена очень слабо, вероятно играет значительную роль в проблеме нежизнеспособности гибридов и должна привлечь гораздо большее внимание исследователей.
Гибридной беременностью можно назвать беременность, при которой в матке самки развивается гибрид. Самка и ее родовые пути могут оказаться при атом неприспособленными к новым свойствам гибридного эмбриона. Это может иметь место и тогда, когда самка чистого вида вынашивает гибрида первого поколения, и тогда, когда у гибридной самки на базе расщепления возникает плод с новыми дальнейшими особенностями.
Подобная патологическая беременность была установлена при гибридизации быков. При покрытии домашних коров зубрами и бизонами не всегда удается получить потомство несмотря на нормальное развитие эмбрионов. Но благодаря, как полагают, сильному развитию плода конец беременности и роды протекают очень трудно, часто с летальным исходом для матери и плода, или сопровождаются околоплодной водянкой и абортом. Бойд (Boyd, 1908,1914) и Гуднайт (1914) считают, что эти тяжелые явления происходят почти исключительно тогда, когда возникает мужской гибрид, благодаря сильному развитию остистых отростков в области холки, раздражающих родовые пути. По данным Аскании-Нова особенно должны страдать молодые коровы. Попытки гибридизации ячек с зубробизонами, правда не многочисленные, привели также к гибели матерей и плодов.
Патологии вылупления
Так как скорлупа и подстилающие ее оболочки у птиц, оболочка икринок у рыб, хитиновые оболочки яиц насекомых и пр. — являются дериватами материнского организма, то при гибридизации, как и в случае беременности, могут возникнуть несоответствия между свойствами оболочки и свойствами эмбриона. Интересный пример подобного явления изучен Крыжановским (неопубликованные данные). Для вылупления из оболочки икринки мальки например леща (Abramis brama) и карася (Carassius carassius) растворяют ее при помощи мелких многочисленных железок (рис. 54), расположенных на голове и выделяющих специфический секрет. У леща (фиг. 2) эти железки развиваются уже на 2-й день эмбрионального развития, у карася (фиг. 1) — на 5-й день, когда и происходит соответственно вылупление. При оплодотворении яиц леща спермой карася вылупление происходит, как и у отцовского вида, на 5-й день. Исследование показало, что и железки гибрида развиваются тоже лишь на 5-й день.
Расположение и число железок у обоих видов различно. У гибридов (фиг. 3) очевидно в результате специфической комбинации генов число железок резко сокращается, они остаются лишь на местах, общих для обоих видов. Легко представить себе, что при еще большем сокращении числа этих железок, при нарушении их функции в результате гибридизации эмбрионы утратят вообще способность к вылуплению и погибнут.
Возможно, что нечто аналогичное мы имеем в описанных Штандфуссом случаях гибели гибридных гусеничек, которые с трудом прогрызали скорлупу.
Относительно птиц изученных примеров подобного рода мы еще не имеем. Однако нам известны случаи достаточно резких различий в строении скорлупы близких видов. Так например один африканский вид франколинов (Francolinus granti) имеет столь толстую и прочную скорлупу, весящую 5,7 г против нормального веса в 1,6 г у близкого вида F. clappertoni, что яйцо выдерживает перекатывание ветром по каменистой почве. Можно ожидать, что гибриды от такой самки с самцами других более тонкоскорлупных видов могут оказаться в затруднительном положении при вылуплении.
 Рис. 54. Развитиижелезок.
У леща (слева), карася (сверху) и их гибрида (справа) (по Крыжановскому).
Рис. 54. Развитиижелезок.
У леща (слева), карася (сверху) и их гибрида (справа) (по Крыжановскому).
Неясна причина резко пониженной жизнеспособности многочисленных гибридов петух × цесарка, полученных Oropoдним (не опубликовано). В качестве одной из гипотез может быть указана разница и величине яиц кур и цесарок. О понижении жизнеспособности цыплят при скрещиваниях крупных кур с мелкими петухами писал Кондырев (1926), но неясно, насколько это понижение не зависело от иных факторов — плохой инкубации, подготовки кур к яйценоскости и пр.
Экологическая нежизнеспособность гибридов
Каждый вид животного имеет свою более или менее специфическую экологию. Он приспособлен к той или иной нище, температуре, влажности и т. д. и является жизнеспособным лишь постольку, поскольку он в соответствующих условиях находится. Белый медведь в Сахаре и лев на Шпицбергене окажутся формами нежизнеспособными благодаря несоответствию их организации ноной экологической обстановке.
Это обстоятельство необходимо иметь в виду при обсуждении проблемы жизнеспособности гибридов. В самом деле, соединяя воедино генотипы двух видов, каждый из которых имел свою особую экологию, мы получаем новую форму животного. Какой должна быть
экология гибрида? Экологические особенности должны в общем подчиняться закономерностям наследственности при гибридизация. Будет ли нормальная экология для гибрида промежуточной между родительскими, мозаичной, атавистической, материнской и т. д. — будет зависеть от конкретных особенностей соединяемых видов.
Весьма трудной задачей является определить, какие экологические условия надо считать оптимальными и даже нормальными для гибрида, располагая лишь данными об экологии его родителей.
Штандфуссом был например получен гибрид между шелкопрядом Изабелла, живущим на приморской сосне, и другим видом, живущим на лиственных деревьях. Чем следует кормить гусеничек этих гибридов, чтобы они не погибли от неподходящей пищи? Гусенички отказались есть хвою сосны. Случайно им могла быть предложена хвоя лиственницы — растения весьма своеобразного, соединяющего в физических свойствах своей хвои элементы настоящей хвои и мягких листьев. Гибридные гусеницы начали ее благополучно поедать. Не располагай экспериментатор лиственницей, он мог бы погубить всех гибридов и отнести их к категории нежизнеспособных.
Не менее поучителен для гибридагогии и пример гибридов Drosophila melanogaster × D. simulans. Оба эти вида разводятся в лабораториях при оптимальной температуре в 26°. Однако уже Стертевантом было установлено, что эта температура является не только не оптимальной для гибридов, но почти летальной. Личинки, правда, при этой температуре развиваются, по куколки почти нацело погибают. Наоборот, при более низкой, комнатной, температуре выход мух из куколок (не считая того пола, который гибнет в зависимости от скрещивания) вполне нормален.
Следующие цифры из работы Керкиса (Kerkis, 1933) наглядно иллюстрируют это решающее влияние разницы в 10°: при 16° вылупилось 86 % вполне нормальных гибридов, а при 26°—0 %. Вредное влияние 26° сказывается уже при действии на личинок, и хотя развитие личинок идет и благоприятно, по вылупление из куколок, даже перенесенных в комнатную температуру, идет очень плохо и лишь у 10 % куколок. Не менее интересно далее, что развитие мужских куколок, получающихся при скрещивании sim. ♂ × ♀ mel. построения XXY, идет одинаково хорошо при обеих температурах. Следует впрочем иметь в виду, что хотя здесь и говорится о влиянии температуры на развитие, но неизвестно, влияет ли температура здесь непосредственно на эмбриогенез или изменяет другие экологические условия — понижает кислотность среды и т. д.
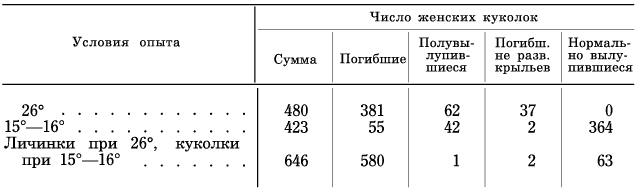
Во всяком случае важно подчеркнуть, что-как бы мы тщательно ни изучили экологию обоих родительских видов, мы не могли бы предсказать, что полученный от них гибрид потребует совершенно иной температуры для своего нормального развития. Только поставив точное изучение самих гибридов, можно было показать, что они не вообще нежизнеспособны, но нежизнеспособны в обстановке, нормальной для чистых родительских видов. Здесь доказать и выяснить преодолимость нежизнеспособности оказалось сравнительно простым благодаря простоте решающего фактора — температуры. Но очевидно, что в других случаях выяснение факторов гибели гибридов может оказаться и чрезвычайно сложной проблемой, до разрешения которой гибрид будет относиться нами к категории «нежизнеспособных» только потому, что мы будем ставить его в неподходящие экологические условия, не умея подобрать подходящие.
Генетический анализ жизнеспособности гибридов
К анализу явлении жизнеспособности (или нежизнеспособности) гибридов можно подходить с двух точек зрения. Можно стремиться к вскрытию непосредственных морфологических и физиологических причин гибели организма, изучая, как мы делали на предыдущих страницах, различные ненормальности в эмбриогенезе и физиологии органов и клеток. Это будет
феногенетический анализ, данные которого чрезвычайно важны для уяснения того, в каком направлении должен быть «подправляем» гибрид для того, чтобы он мог стать жизнеспособным. В другой плоскости анализируется проблема методами
генетического анализа. Здесь ставится вопрос о том, от каких хромосом, участков хромосом и отдельных генов скрещиваемых видов зависит то или иное патологическое изменение гибрида. Данные генетического анализа не менее важны для разрешения проблемы преодоления нежизнеспособности гибридов, так как они освещают вопрос о том, в каком направлении должна быть изменена наследственная структура родительских видов для того, чтобы они не давали нежизнеспособной комбинации при скрещивании друг с другом.
Указания на то, что нежизнеспособность гибридов может в решающей степени зависеть от отдельных хромосом, мы получаем из сравнения результатов реципрокных скрещиваний. Как будет подробнее рассматриваться ниже, при реципрокных скрещиваниях двух чрезвычайно близких видов дрозофил — Drosophila melanogaster и D. simulans — получаются совершенно различные результаты. В обширном последнем опыте Биддля (Biddle, 1932) он получил:
Дочерей/Сыновей
melanogaster ♀ × ♂ simulans — 20 388/41
simulans ♀ × ♂ melanogaster — 437/10 870
Аналогичную картину получил впервые Стертевант, открывший это замечательное явление, при котором одно скрещивание дает лишь дочерей, а другое — лишь сыновей (о незначительных исключениях скажем далее). Объяснений этим фактам может быть два: либо в результате скрещивания нарушается правильное определение пола и мы принимаем одни пол за другой, либо происходит отмирание одного из полов. Стертевантом была показана правильность второго объяснения. Если продержать мух-родителей в банке один день и иметь таким образом яйца, отложенные в одно время, то очень скоро, на третий день развития, можно различить в культурах два типа личинок — более крупных и более мелких. Первые развиваются нормально и дают исключительно самок (в скрещивании mel. ♀ × ♂ sim.). Более мелкие личинки продолжают отставать, низ них ничего не вылупляется. Они оказываются мужскими личинками.
Обратим внимание на следующее: гибель мужского пола не может быть объяснена тем, что особенности строения этого пола делают вообще невозможной жизнь самцов-гибридов двух названных видов. В этом случае самцы гибли бы и в реципрокном скрещивании, однако здесь они живут нормально. Следовательно причину разницы в этих реципрокных скрещиваниях надо искать в том же направлении, как и разницу в реципрокных скрещиваниях, вообще — в особенностях половых хромосом. Данный случай является, правда, несколько более сложным, и к нему мы вернемся ниже. Однако гибель одного пола встречается при гибридизации нередко.
При гибридизации бабочек из 24 гибридных сочетаний, известных Штандфуссу, в 5 сочетаниях были получены только самцы, в других 5—только самки, а в 7 случаях — оба пола, но самки значительно реже.
Аналогичные явления мы имеем повидимому и в скрещивании шпорцевого гуся (Plectropterus) с мускусной уткой (Cairina moschata). Гейнрот (Heinroth, 1910) получил в этом скрещивании одних самцов, причем и среди задохликов ни одной самки не было обнаружено (Poll. 1920, стр. 383).
Случай с бражниками был подвергнут Федерлеем детальному генетическому анализу, выяснившему хромосомный механизм гибели определенных категорий гибридов. При скрещивании двух видов бражников (Deilephila elpenor — вьюнковый бражник с Deilephila porcellus — кипрейный бражник) реципрокные скрещивания дают различный результат. В скрещивании elp. ♂ × ♀ роге, получаются оба пола в нормальном отношении. В реципрокном скрещивании роге. ♂ × ♀ elp. возникают одни самцы. Самки достигают стадии куколки, но не превращаются в имаго и умирают после двукратного перезимовывания. Федерлей назвал это явление «сублетальностью», так как до стадии куколки жизнеспособность сохраняется.
Обозначив маленькими буквами
е и
р хромосомы соответствующих видов, можно дать такую схему наследования половых хромосом в этих скрещиваниях:

Комбинация хромосом X
рY
е оказывается таким образом сублетальной. Правильность гипотезы, что сублетальность действительно связана с этой комбинацией хромосом, Федерлей доказывает рядом дальнейших скрещиваний.
Скрещивания эти ставятся таким образом, чтобы разными способами и в разных пропорциях получить X
pY
e — комбинацию.
Эксперименты эту гипотезу подтверждают — независимо от типов скрещивания — каждый раз, как возникает сублетальная комбинация хромосом, соответствующая категория бабочек теряет способность к гистолизу и гистогенезу и гибнет.
Приведем пример. Гибридный самец, которого Федерлей обозначает porcelp, может быть скрещен с чистой самкой elpenor по следующей схеме:
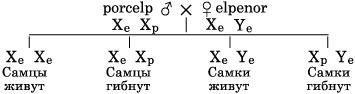
В этом скрещивании лишь половина самок получает сублетальную комбинацию хромосом и должна по гипотезе погибнуть. В результате должно получиться ненормальное соотношение полов — 2 самца: 1 самка. Фактически было получено 67 самцов и 37 самок, т. е. отношение, почти совпадающее с теоретическими ожиданиями 69,3 самцов: 34,7 самок. Разница в 2,3 бабочки совершенно несущественна. Что касается сублетальных женских куколок, то их было получено меньше 25 % общего числа куколок, и, судя по значительной гибели гусениц женского пола, Федерлей полагает, что сублетальная комбинация X
pY
e начинает здесь действовать еще в гусеничной стадии (см. ниже).
Описанные явления не являются исключительными и наблюдаются и у других видов в различных оттенках. Особенно трудно совместима Y-хромосома elpenor с Х-хромосомой других видов — с Deilephila euphorbiae, D. galii. Самцы в этих скрещиваниях вполне жизнеспособны. В свою очередь Y-хромосома D. euphorbiae, молочайного бражника, образует летальные комбинации с Х-хромосомой других видов — D. elpenor, D. galii. Здесь самки с Y
euph. не появляются никогда или лишь в виде редчайших исключений (Федорлей, 1930).
Подобные же явления наблюдаются и в других группах бабочек. При скрещивании бабочек-кисточниц Pygaera curtula и P. anachoreta гибридные самцы Х
аХ
с и самки X
aY
c очень быстро заканчивают свое развитие, в то время как самки X
cY
a развиваются очень медленно. Эта задержка расстраивает метаморфоз и вызывает гибель гусениц, так что лишь в виде исключения удавалось довести их до вылупления. Та же самая хромосома Y
a оказывает задерживающее действие и в скрещивании с третьим видом P. pigra: гусеницы с комбинацией X
pY
a не доходят до стадии куколки и погибают.
В чистом генотипе P. anachoreta это тормозящее действие хромосомы Y
a очевидно уравновешивается противоположным убыстряющим действием хромосомы Х
а (у самок X
aY
a). Это убыстряющее действие хромосомы Х
а сказывается и у гибридов, и особенно заметно у гибридов с pigra. Здесь гусеницы строения Х
аХ
р развиваются очень быстро и вылупляются после кратковременной куколочной стадии и никогда не перезимовывают.
Очень важно подчеркнуть вытекающий из этих опытов и подчеркиваемый Федерлеем вывод о том, что необходимо говорить не о летальных, (или сублетальных), свойствах топ или иной хромосомы, но именно о летальных
комбинациях. Сама по себе Y
a-хромосома P. anachoreta не является летальной уже потому, что она эволюционно развилась во вполне жизнеспособном диком виде и в нем играет определенную не летальную, а витальную роль. В этом виде бабочек генотип так распределен между хромосомами Х
а и Y
a, что обеспечивает нормальное согласованное развитие обоих полов. То же самое П: у видов P. curtula и P. pigra. Но соединенные в одну комбинацию ×
pY
a уже не обеспечивают нормального развития, и организм гибнет; комбинация оказывается летальной или сублетальной. Еще яснее это видно у Drosophila melanogaster. Здесь самки имеют нормальное строение ХХАА, где буквами Х обозначаются хромосомы половые, а буквами А — аутосомы. Возможно получение мух строения ХХХАА. Это так называемые сверхсамки. Жизнеспособность их чрезвычайно понижена, и они вылупляются в качестве сравнительно редких исключений. Таким образом комбинация ХХХАА оказывается в высокой степени летальной, хотя состоит из вполне нормальных хромосом. Что летальным здесь оказывается не факт наличия трех Х-хромосом (XXX), показывает значительно более нормальная жизнеспособность триплоидных самок строения ХХХААА. Здесь благодаря прибавке третьего А, набора аутосом, восстанавливается нормальное (витальное) отношение 1X:1А и жизнеспособность восстанавливается.
Подобный генетический анализ нежизнеспособности гибридов может быть доведен и дальше, показывая, что в определении ее решающую роль могут играть не целые хромосомы, но отдельные гены. В этом отношении чрезвычайно интересны экспериментальные исследования Мирона Гордона, Коссвига и др. над гибридами двух близких родов аквариумных рыбок-живородок: меченосца Xiphophorus и Platypoecilus. Platypoecilus maculatus встречается в виде разнообразных цветных. рае — золотистые, с мелкими и более крупными черными пятнышками, с большими черными пятнами на боках и пр. При скрещивании золотистой расы с меченосцем гибриды получаются вполне жизнеспособными. При скрещивании же пятнисто)! расы с меченосцем результат получается неожиданный. Черные пятна, наследуемые гибридом, получают столь сильное развитие, что приводят к патологическому развитию пигмента (меланозису) — черным раковидным опухолям на теле, к разрушению плавников и к массовой гибели мальков на разных стадиях (см. цветную таблицу I). Здесь следовательно гибридизация приводит к патологическим, нежизнеспособным формам гибридов.
Гордон показал, что этот меланозис развивается вполне закономерно и лишь в том случае, если в скрещивание вводятся определенные расы платипецилии, так называемые pulchra, rubra или nigra. Эти три расы, имеют, общие генотипические черты — наличие в их половой хромосоме особого гена Sp (Spotted — пятнистый). В различных расах носителями черного пигмента являются или мелкие более или менее округлые пигментные клетки, микромеланофоры или, наоборот, крупные, ветвистые макромеланофоры. Возникновение макромеланофоров именно и связано с наличием гена Sp; Специальными скрещиваниями легко показать, что меланозис у гибридов. развивается не вообще от скрещивания с названными тремя расами, но лишь в присутствии гена Sp. Этот ген, как сказано, локализован в половой хромосоме, и его роль легко выясняется сравнением реципрокных скрещиваний.

В скрещивании самца Plat, с самкой Xiph. все потомство, как сыновья, так и дочери, получает ген Sp и оказывается пораженным меланозисом. В реципрокном скрещивании ген Sp от матери попадает лишь к сыновьям, дочери же вовсе не получают гена Sp и оказываются здоровыми, тогда кап все их братья заболевают.
Что здесь играет решающую роль не сам факт гибридизации, а лишь комбинация генов, показывает дальнейшее скрещивание. Если из второго скрещивания взять гибридную здоровую самку и скрестить ее обратно с самцом Plat, несущим ген Sp, то меланозис возникает вновь, так как гибридная самка привносит часть генов Xiphophorus. Что касается летального действия гена Sp, то опять приходится сказать, что было бы неверно приписывать летальное действие лишь этому гену. В чистых расах Platypoecilus он вызывает лишь развитие макромеланофоров и нисколько не детален. Но, попадая в генотип гибрида, он образует с некоторыми генами вида Xiphophorus как повое качество уже летальную
комбинацию, которая и приводит к развитию меланозиса, раковоподобных опухолей и губит организм.
Роль цитоплазмы в определении жизнеспособности гибридов
Генетический анализ явлений нежизнеспособности гибридов показал, что они не могут быть сведены к действию лишь одного генотипа, по крайней мере в некоторых случаях, и что для полного выяснения этих явлений необходимо учитывать и нехромосомные элементы. Эти нехромосомные элементы обозначаются обычно как цитоплазма. Хотя вероятно правильнее иметь здесь в виду строение яйца как целого.
Первым и до сего времени наиболее изученным случаем такого рода является упомянутое скрещивание двух видов дрозофил. Стертевантом (1920) были получены гибриды, как simulans
♂ × ♀ melanogaster, так и реципрокно. При этом обнаружилось замечательное явление, что гибридное потомство оказывается всегда одного пола, женским — в скрещивании sim.
♂ × ♀ mel.. и мужским — в скрещивании mel.
♂ × ♀ sim.
Если проследить судьбу различных хромосом, участвующих в этих скрещиваниях, то мы получим следующую схему:
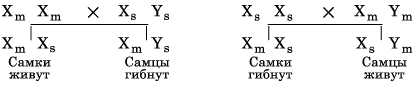
Сравнив хромосомную структуру дочерей в обоих скрещиваниях, мы видим, что она генотипически совершенно одинакова. Однако в первом случае самки живут, во втором — гибнут.
При таких условиях объяснить гибель личинок одним лишь неудачным хромосомным генотипом гибридов оказывается невозможным. К чему же может быть в таком случае сведено различие в жизнеспособности обоих типов самок? Единственное различие между ними сводится к различию в происхождении
плазмы тех яиц, из которых развиваются личинки. Обозначив через С
m и С
s эти плазмы, получим следующую структуру гибридных самок:
C
mX
mX
s — живут
C
sX
mX
s — гибнут
Объяснения этой роли цитоплазмы яйца мы пока еще не имеем, и еще трудно дать гипотезу о причинах нежизнеспособности самок С
sХ
mХ
s. Привлечение двух типов самцов
С
mХ
mY
s — гибнут
С
sХ
sY
m — живут
тоже недостаточно для понимания, хотя сопоставление, различающееся лишь по одному элементу, а именно:
C
mX
mX
s — живут
C
mX
mY
s —гибнут
C
sX
sX
m — гибнут
C
sX
sY
m — живут
говорит за то, что в обстановке С
m замена Х
s на Y
s приводит к гибели, а в обстановке С
s — замена Y
m на Х
m.
Значительную ясность вносит в вопрос скрещивание самок mel. XXY, т. е. имеющих сцепленные X-хромосомы. Они же имеют всегда Y
m-хромосому.
X
m-X
mY
m × X
sY
s
X
m-X
mY
m X
sY
m
Самки Самцы
гибнут живут
C
mX
mY
s — самцы гибнут
C
mX
sY
m — самцы живут
C
mX
mY
s — самки живут
C
mX
mX
mY
s — самки гибнут
Сравнивая эти категории, мы видим, что гибнут мухи всякий раз, как Y
s встречается с С
m, а в отсутствии Y
s как C
mX
mY
m, так и C
mX
sX
m живут. К сожалению относительно С
s за отсутствием самок X
s-X
s такой анализ еще не выполнен. И хотя об окончательном решении вопроса о причинах гибели рассматриваемых гибридов говорить еще трудно, тем не менее роль различия в плазме кажется вероятной.
Жизнеспособность гибридов дальнейших поколений
Изучение дальнейших поколений гибридов F
2, F
3 и т. д. и возвратных скрещиваний F
b знакомит нас с новыми явлениями. В ряде случаев вполне жизнеспособные гибриды F
1 дают в F
2 резкое понижение жизнеспособности вплоть до невозможности получения жизнеспособного потомства вообще. В других случаях, наоборот, жизнеспособность гибридов дальнейших поколений возрастает по сравнению с F
1. Это разнообразие явлений может быть уже во многих случаях объяснено.
Таблица I
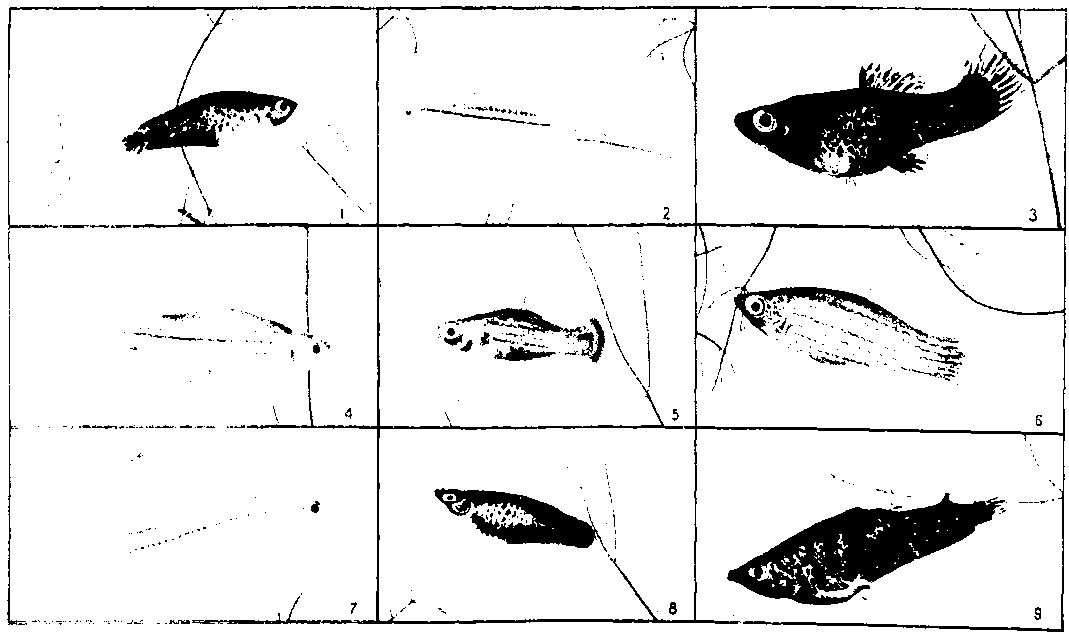 Меланозис у гибридов Xiphoporus и Platypoecilus.
1 — ♀ Platy. (Sp.), 2 — ♂ Xiph., 3 — их гибрид Sp с меланозисом, 4 — ♀ Xiph., 5 — ♂ Platy. (Sp.), 6 — их гибрид sp без меланозиса, 7 — ♀ Xiph., 8 — ♂ Platy. (Sp.), 9 — их гибрид Sp с меланозисом.
Меланозис у гибридов Xiphoporus и Platypoecilus.
1 — ♀ Platy. (Sp.), 2 — ♂ Xiph., 3 — их гибрид Sp с меланозисом, 4 — ♀ Xiph., 5 — ♂ Platy. (Sp.), 6 — их гибрид sp без меланозиса, 7 — ♀ Xiph., 8 — ♂ Platy. (Sp.), 9 — их гибрид Sp с меланозисом.
В опытах Штандфусса гибридные самцы Saturnia pavonia × pyri скрещивались обратно с ♀ pavonia. В пяти случаях вылупилось соответственно 43, 45, 48, 54 и 62 % отложенных яиц, 50 % в среднем. При этом в двух первых случаях гусенички лишь отчасти прогрызли скорлупу яиц и погибли, не выведшись окончательно. В следующий раз было получено значительное количество гусениц, которые тоже погибли, не успев даже принять пищи. Полного развития достигли лишь 3 ♂ и 1 ♀. От 4-й и 5-й кладок было получено 80 гусениц, которые были вначале вполне здоровы, но большинство погибло от грибного заболевания после 3–4 линек кроме 12 самцов. Одна из гусениц имела вместо 4 нормальных 5 линек. Всего получилось следовательно 15 ♂ и 1 ♀.
При скрещивании таких же гибридных самцов F
1 обратно с другим родительским видом, pyri, от 9 самок из около 1 600 яиц вылупилось всего 10 гусеничек. Из этих 10 две погибли, не успев принять пищи, две другие — взрослыми от грибного заболевания, а 5 превратились в бабочек — 3 самца, 2 самки и 1 гермафродит.
В этих тщательных и труднейших опытах обнаружились все три характерных явления — резкое понижение вылупляемости и жизнеспособности, особенно резкая убыль самок — гетерозиготного пола и резкие колебания в жизнеспособности отдельных особей, как например в последнем случае.
Сильное понижение жизнеспособности в F
2 и в F
b возникает прежде всего тогда, когда у гибридов оказывается нарушенной правильность гаметогенеза, например при нарушении правильности конъюгации хромосом и возникновении гамет с неполным или излишним числом хромосом. Штандфуссу удавалось не раз получать гибридов сатурний, но лишь ничтожное число дальнейших потомков. В подобных случаях благодаря недоброкачественности гамет идут параллельно два явления — понижение плодовитости гибрида, появление нежизнеспособных ненормальных яиц и малая в среднем жизнеспособность и тех гибридов, которые начинают все же свое развитие. Эта картина и по внешности и по существу напоминает то, что мы встречаем при размножении триплоидов, большинство гамет которых оказывается с неправильными наборами хромосом.
Далее нежизнеспособные формы гибридов F
2 получаются и при правильном гаметогенезе на почве комбинаторики генов. Адаптивные комбинации генов, которые мы находим в чистых видах, подвергавшихся длительному естественному отбору, в F
2 начинают рассыпаться на составные части, образуя множество новых комбинаций, среди которых очевидно высокий процент должен давать мало или вовсе нежизнеспособные формы гибридов. О подобных явлениях упоминает например Федерлей, получивший F
2 от вполне жизнеспособных и плодовитых гибридов бражников elpenor ♂ × ♀ porcellus. При скрещивании шелкопрядов гарпий Федерлей наблюдал у вполне жизнеспособных самок разрушение ядер во внешне нормальна развитых яйцах, а при скрещивании шелкопрядов падубного с осиноволистным — даже недоразвитие яиц (см. следующую главу).
В F
2 от скрещивания Xiphophorus × Platypoecilus возникают целые категории карликовых самцов, о чем
-будет подробнее сказано тоже ниже. Подобные факты не представляют собой не только чего-либо странного, но целиком предсказываются, исходя из современного учения о наследственности.
Также понятны в общем и противоположные факты повышения жизнеспособности в дальнейших поколениях. Как. было упомянуто, при скрещивании бражников porcellus ♂ × ♀ elpenor все самки оказываются сублетальными в связи с комбинацией X
pY
е. В следующем поколении куколок с такой комбинацией оказывается заметно меньше теоретически ожидающихся 25 %. Так в обратном скрещивании porcelp. ♂ × ♀ elpenor получено:
самцов Х
eХ
e и Х
eХ
р — 50
самок Х
еY
е — 26
самок (куколок) X
pY
e — 12
Разница между 12 и 26 очень отчетливо указывает на то, что здесь в F
b летальность комбинации X
pY
e усилилась за счет очевидно комбинации с другими генами. Наоборот, в скрещивании самок X
eY
e (аналогичных 26 указанным в табличке) с ♂ porcellus, где сублетальная комбинация X
PY
e вновь должна была бы получиться (Х
рХ
р × X
eY
e → ♂ X
pY
e + ♀ X
pY
e), из 6 женских куколок три дали бабочек, т. е. комбинация хромосом X
pY
e уже оказалась сублетальной лишь в 50 %. Правда, эти бабочки вывелись с запозданием. Самцы вылупились 16–29/VII, в то время как первая ♀ только обнаружила признаки метаморфоза. 2/VIII она полумертвой была извлечена из оболочки куколки. Вторая самка вылупилась 8/VIII и третья — 6/IX. Тем не менее это лишь подчеркивает, что на почве генетической комбинаторики в F
2 и пр. жизнеспособность обнаруживает большую изменчивость и у некоторых особей оказывается пониженной по сравнению с F
1; а у других особей, наоборот, повышенной. Это открывает перспективы преодоления нежизнеспособности и генетическими методами.
Проблема преодоления нежизнеспособности гибридов
Эта проблема в общем виде является очевидно самой трудной проблемой гибридизации по глубине и разнообразию тех причин, которые могут приводить к гибели гибрида, по обширности физиологических, морфологических и экологических знаний, которые должны быть мобилизованы для ее разрешения. Очевидно поэтому, что нежизнеспособность гибридов кладет те пределы гибридизации, которые будет труднее всего преодолевать.
Тем не менее предыдущее изложение убеждает нас в том, что нежизнеспособность гибридов ни в коем случае не может рассматриваться как нечто абсолютное даже там, где она установлена прямым опытом. Мы только что видели, что гибриды дрозофил превращаются из нежизнеспособных в жизнеспособных простым понижением температуры; что гибриды Xiphophorus × Platypoecilus превращаются из патологических в совершенно жизнеспособных простым устранением из скрещивания, одного лишь гена Sp; что процент благополучных родов и скрещивании корова × бизон может быть повышен подбором много-рожавших коров и т. д.
Дать какие-либо универсальные методы для борьбы с нежизнеспособностью гибридов (там, где она наблюдается) конечно невозможно по многообразию причин, могущих убивать гибрида. Однако усвоение относительности нежизнеспособности очень важно, так как оно должно направить нашу мысль на отыскание таких условий, которые делали бы гибрида жизнеспособным. Где и как эти условия искать— должно быть вскрыто анализом каждого конкретного случая. При воспитании гибридов надо вариировать условия их питания, температуру, влажность и другие внешние факторы. Нужно иметь в виду при этом частое явление атавизации гибридов (глава IX) и на основании изучения зоогеографии и эволюции данной группы видов делать опыты к воссозданию для гибрида экологических условий его предков. Например если два гибридизируемые вида недавно распространились на север, можно ожидать, что гибрид между ними потребует более южной обстановки для своего развития и т. п.
При получении мало жизнеспособных гибридов с нашими домашними животными, учитывая опыт гибридизации Xiphophorus × Platypoecilus, следует вводить в скрещивание различные, самые разнообразные породы домашних животных в расчете на то, что хотя бы случайно попадется порода, дающая более жизнеспособную комбинацию с данным видом. Следует испытывать также не чистые породы, а сложных метисов или даже гибридов в расчете на то, что в числе многочисленных генокомбинации могут возникнуть такие, которые дадут с диким видом жизнеспособную комбинацию. Должны быть обязательно испробованы оба реципрокных скрещивания, так как мы видели, что результаты их могут оказаться существенно различными. Следует наконец применить метод гибридизационных мостов, рассчитывая на то, что если виды
А и
С дают друг с другом нежизнеспособное потомство, то может быть найден промежуточный вид
В, который дает жизнеспособных гибридов как с
А, так и с
С и в дальнейшем даст возможность получить жизнеспособных гибридов и от скрещивания
АВ ×
ВС (подробнее см. следующую главу).
Таким образом поле для исследовательской работы по этой проблеме чрезвычайно широко, и уже имеющиеся скудные исследования Показывают, что и здесь могут быть достигнуты крупнейшие результаты, которые шаг за шагом будут раздвигать границы гирконов.
Глава VIII. Проблема размножения гибридов
Значение проблемы
Проблема размножения гибридов является — по крайней мере в настоящее время — главнейшей проблемой гибридизации. Такое ее значение обусловливается тем, что уже сейчас мы имеем возможность получать огромное разнообразие гибридов, дальнейшая работа с которыми упирается в бесплодие громадного большинства этих гибридов, неспособность их давать дальнейшее потомство. Поэтому, С одной стороны, уже одно разрешение проблемы преодоления этого бесплодия открыло бы перед нами необъятно широкие перспективы гибридизации, перспективы коренного переустройства животных организмов, и, с другой стороны, до разрешения этой проблемы утрачивают значительную долю своей актуальности и такие проблемы, как проблема гетерогенного оплодотворения, проблема жизнеспособности, ибо там, где очень затруднено оплодотворение и получаются мало жизнеспособные гибриды, в большинстве случаев мы можем ожидать при преодолении этих затруднений появление лишь бесплодных гибридов.
Приведем в качестве примера мулов и других гибридов в роде лошадей (Equus). Человек получает в огромных количествах и ужо в течение нескольких тысячелетий гибридов лошади с ослом, руководясь. ценными качествами F
1 поколения. В последнее столетие часто получались и разнообразные гибриды лошадей и ослов С зебрами, куланами и другими лошадиными. Техника получения этих гибридов оказалась достаточно простой и доступной малокультурным коневодам, а с применением искусственного осеменения эта гибридизация еще упрощается, как показали опыты получения зеброидов в Аскании-Нова.
Вводимые в эти скрещивания виды лошадей и ослов различаются между собой чрезвычайно существенно, — достаточно сравнить обычную лошадь с ослом. Несомненно, что эти различия зависят от множества генов. И поэтому несомненно также и то, что если бы мы преодолели бесплодие мулов и лошадей, то получили бы возможность иметь в F
2 богатейшее разнообразие генотипов, богатейшее многообразие сочетания признаков осла и лошади, с возникновением новых, неожидаемых фенотипов (см. главу IX). И конечно среди этого многообразия можно было бы найти формы, гораздо более интересные, для народного хозяйства, чем каждый из исходных видов. В то же время эти расщепления позволили бы подвергнуть оба вида глубокому генетическому анализу и добыть много ценнейшего материала для понимания строения и жизни организма лошади.
Сейчас эти возможности закрыты от пас фактом бесплодия мулов и зеброидов. Правда, время от времени в литературе появляются сведения об отдельных случаях плодовитых мулиц. Ничего невероятного в этих сообщениях конечно нет. Возможно, что при тщательном и широком изучении такие экземпляры и будут обнаружены. Но на сегодня все сообщенные сведения о плодовитых мулах либо сомнительны (так как всегда есть возможность принять за мулицу обычную лошадь, несколько уклоняющуюся по экстерьеру) либо не имеют уже значения, поскольку сами животные уже не существуют и для проверки фактов нужны дальнейшие поиски таких плодовитых мулов.
Еще интереснее в этом отношении птицы, в частности утиные и гусиные. Здесь уже в настоящее время еще до применения метода искусственного осеменения известно огромное количество разнообразнейших гибридов.
Так, среди гибридов с участием утки-кряквы мы видим представителей почти всех подсемейств обширного семейства вплоть до гусей к крохалей. Это дало нам возможность занести в гибридоресурсы утки все семейство Anatidae с его более чем 250 видами самого разнообразного характера по своей анатомии, морфологии, экологии, очевидно и физиологии.
Тем не менее в настоящее время эти ресурсы остаются за семью печатями бесплодия, так как до сего времени плодовитыми оказались лишь гибриды утки с шилохвостью и с некоторыми другими уже совсем близкими к крякве видами. На другом конце мы имеем плодовитых, гибридов гусей серого, гуменника и сухоноса (уже давших такие хозяйственно ценные гибридные формы, как холмогорские гуси) — и этим пока исчерпываются наши фактические возможности размножения гибридов Anatidae.
Можно сказать поэтому, что в настоящее время в группе уток в качестве плодовитых гибридов мы имеем примерно 1 % всех возможных, а вернее и того меньше, и при этом плодовитой оказывается наименее интересная часть всех возможных гибридов, так как она получается в результате скрещивания очень близких видов.
Этими соображениями мы хотим обратить внимание исследователей на то, какой переворот обещает гибридизационной работе хоти бы частичное разрешение проблемы преодоления бесплодия гибридов. Необходимо сомкнутыми рядами, с разных сторон атаковать эту проблему из проблем, чтобы в кратчайший срок разрешить ее. Никакие усилия не будут здесь нелишними.
Проблема бесплодия гибридов распадается на две более или менее самостоятельные — бесплодие первого поколения гибридов и бесплодие дальнейших поколений. В свою очередь эти проблемы теснейшим образом связаны — mutatis mutandis — с проблемой пола гибридов. Для разрешения проблемы преодоления бесплодия предстоит мобилизовать весьма различные науки, о чем мы скажем по ходу изложения.
Вариация степени плодовитости гибридов первого поколения
Фактический материал, имеющийся по этому вопросу, обнаруживает, как и следовало ожидать, полные и постепенные переходы от полной плодовитости до полного бесплодия и притом несколько различно у разных полов. С другой стороны, и бесплодие тоже не является однородным— оно имеет свои степени, вскрываемые более глубоким анатомо-гистологическим и Цитологическим изучением.
Вполне плодовиты прежде всего гибриды домашнего тура с зебу, овец с муфлоном, собаки с волком, свиньи с кабаном, коз с дикими козлами и т. д. В последних случаях однако мы имеем дело не с настоящей гибридизацией, так как упомянутые дикие виды являются родоначальниками домашних форм.
Однако и среди настоящих видов мы имеем примеры полной или почти полной плодовитости, границы между которыми нелегко обнаружить. Так, не подлежит никакому сомнению, что утка-кряква и шилохвость (Dafila acuta) являются хорошими видами, тем более что были попытки выделить шилохвости даже в отдельный род Dafila. Также несомненными видами и даже родами являются гусь-гуменник (YIelanotus segetum) и гусь-сухонос (Cygnopsis cygnoides), дающие тем не менее плодовитое потомство. Плодовиты гибриды домашних коз с альпийскими козерогами (Capra ibex), хотя их принадлежность к разным видам не подлежит сомнению. Плодовиты гибриды двугорбого и одногорбого верблюдов. Плодовиты зубро-бизоны.
Среди рыб вполне плодовиты такие несомненные гибриды, как Platypaecilus × Xiphophorus.
Однако несомненно, что случаи полной плодовитости составляют незначительное меньшинство среди гибридов.
Первой ступенью к бесплодию является более или менее полное нарушение плодовитости одного пола. Так, у гибридов як × тур самки плодовиты и невидимому вполне нормально, самцы же вовсе бесплодны (Лус) либо может быть изредка плодовиты. Такая же картина наблюдается у гибридов як × зебу (Завадовский, 1931), тур × зубр, тур × бизон, тур × гаял и т. п.
Мужская плодовитость нарушается чаще, чем женская, по это повидимому только в случае гетерогаметичности мужского пола, как например у млекопитающих. Что касается бабочек, у которых гетерогаметичны самки, то последние страдают обычно сильнее самцов. Так, в скрещивании Saturnia pyri × S. pavonia в опытах Гольдшмидта и Паризер все самки 1 были совершенно бесплодны, тогда как у самцов «из дюжин скрещиваний с тысячами яиц вывелось лишь 5 гусеничек». В более ранних опытах Штандфусса (1908) было получено в F
a 27 гермафродитов на 282 индивидуума (см. также Standfuss, 1908, 1914).
Штандфусс скрещивал гибрида F
1 (Saturnia pavonia
♂ × ♀ S. spini) с обоими родительскими видами и с ♀ S. pyri. Интересно, что с pavonia удались 2 скрещивания и процент вылупления гусениц был 16 и 92. С самками второго родительского вида удались то же 2 скрещивания с 94 и 98 %. И наконец с третьим видом pyri получилось тоже 92 % гусеничек. Таким образом самцы показали высокую плодовитость несмотря на полное бесплодие самок. Это обстоятельство будет иметь для нас существенное значение при обсуждении причин бесплодия и методов преодоления его.
Полученные Штандфуссом гибриды-самки сатурний (S. pavonia ♂ × ♀ spiui), как и другие гибридные самки, показали, что яичники их не содержали способных к развитию яиц. В них заключались лишь незначительные зачатки яичек в самых тонких, верхних частях яичников. Однако одна из таких самок резко выделилась тем, что отложила 16 яиц, которые, правда, были очень малы и уродливы, большей частью полушарообразны, как бы разрезанные пополам нормальные яйца. Вскоре после откладки они высохли.
При дальнейших ступенях бесплодия нарушается плодовитость и другого пола.
Постепенно мы подходим уже к бесплодным гибридам. Здесь мы встречаем например такой случай, как гибриды мускусной и Домашней утки, где в одном скрещивании самки еще могут откладывать яйца, неспособные однако к развитию. В реципрокном сочетании тех же видов самки оказываются утратившими и эти последние отзвуки плодовитости.
Эта разница в плодовитости реципрокных гибридов, к сожалению совершенно недостаточно изученная у животных, чрезвычайно важна для нас 1) потому, что указывает на отсутствие полной связи между таксономическим родством и плодовитостью гибридов, и 2) тем, что снова, как и при оплодотворении, подчеркивает возможность добиться плодовитости путем применения реципрокного скрещивания там, где первое скрещивание дало бесплодных гибридов. Отметим здесь, что при гибридизации домашних животных с дикими в подавляющем большинстве случаев было поставлено лишь одностороннее скрещивание — обычно самкой служила домашняя форма и самцом — дикая (реципрокные скрещивания проводились лишь менаду лошадью и ослом — мул и лошак, — одногорбым и двугорбым верблюдом, яком и коровой).
Что такое бесплодие гибридов
Поль (H. Poll) был одним из первых, кто подверг проблему бесплодия гибридов более тщательному изучению. Он рассмотрел в основном на гибридах птиц строение гонад гибридов и ход сперматогенеза, а вместе с тем поставил вопрос о том, что надо разуметь под бесплодием гибридов.
Вопрос о том, что надо разуметь под бесплодием гибридов, т. е. бесплодием, которое так или иначе связано с гибридизацией, возникает в результате гибридизации, — вовсе не так прост. К соображениям Поля, о которых будет речь ниже, мы можем прибавить следующее: проблема бесплодия гибридов тесно связана с проблемой экологии и физиологией гибридов, о которой подробнее мы говорили уже и главах V и VII. В самом деле для ряда животных установлено, что они приступают к размножению лишь при наличии определенных экологических условий и определенном физиологическом состоянии. По линии экологии например известно, что птицы, гнездящиеся в дуплах, не размножаются в неволе даже в хороших вольерах, если им не дано дупляков или других домиков. Выяснить, какие условия необходимы для того, чтобы заставить вид размножаться в неволе, далеко не так легко, и известно, что в зоологических садах целый ряд диких животных и б размножается несмотря на все старания ухаживающего персонала. Так например известен всего один случаи размножения в одном английском парке краснозобой казарки (Rufibrenta ruficollis) и совершено неизвестно ни то, чего ей недостает в зоопарках, ни то, почему она стала размножаться в этом английском парке. Как известно, не размножается в неволе и слон.
Это будет
экологическое бесплодие, которое явным образом связано и с рефлексологическими моментами. Несомненно, что с этой формой бесплодия — вернее неразмножения — нужно очень считаться при обсуждении проблемы бесплодия гибридов. В самом деле, в главе VII мы указывали на проблему экологии гибрида и на трудности решения ее. Если мы получаем гибрида двух видов птиц, то естественно возникает вопрос, в какие экологические условия мы должны поместить этих гибридов, чтобы не вызвать у них экологического бесплодия, т. е. чтобы обеспечить достаточный нажим на репродуктивную систему со стороны центральной нервной системы. Поскольку мы в этой области еще чистые эмпирики, поскольку мы не умеем решить например вопрос о причинах, мешающих размножению краснозобой казарки, постольку вполне возможно, что часть бесплодия гибридов, получившихся в зоопарках, может быть отнесена за счет неумения решить,
- в какие экологические условия необходимо этих гибридов поставить.
Не Менее важно и
физиологическое бесплодие, несвязанное с гибридизацией. В тех же зоопарках и зоосадах многие животные требуют специальных условий кормления и ухода, приведение их в соответствующее «тело», без чего они тоже не размножаются. Мантейфель например придает большое значение, с одной стороны, тому, чтобы вес животного находился в нужном соотношении к величине, так как здесь существует повидимому довольно узкий оптимум, и ни недокормленные ни перекормленные животные не размножаются. Не меньшее значение придается и качественному составу пищи, наличию витаминов и т. д. В Московском зоопарке Мантейфелю удалось добиться подобными экологическими и физиологическими приемами размножения соболя, чего не удавалось раньше. По отношению к гибридам
вопрос этот снова осложняется, так как далеко не всегда можно быть уверенным в том, что потребности гибрида будут более или менее промежуточными (ср. случаи с шелкопрядами, глава VII) между потребностями родительских форм.
Таким образом можно с уверенностью думать, что проблема бесплодия гибридов состоит из двух частей: 1) экологическо-физиологического бесплодия, связанного с нашим незнанием потребностей гибрида, 2) собственно гибридного бесплодия, связанного с нарушением его половой деятельности на почве гибридизации.
Подобные соображения заставили и Поля утверждать, что одного факта неполучения потомства от гибрида еще недостаточно для того, чтобы отнести его в категорию бесплодных, и для точного диагноза необходимо более тщательное изучение гаметогенеза
[35].
Классификация гибридов по гаметогенезу. Работы Поля
В 1906–1912 гг; появился ряд гибридологических работ Генриха Поля (Poll Н.), выполненных им по обширным и исключительно интересным материалам преимущественно Берлинского Зоосада, в которых Поль подверг систематическому изучению анатомию и гистологию гонад длинного ряда плодовитых и бесплодных гибридов птиц (главным образом куриных и утиных). На основании этих исследований Поль попытался установить некоторые закономерности в появлении тех или иных ненормальностей половой сферы гибридов II предложил их классификацию, имеющую для гибридологии большой интерес.
Еще в работе 1907 г. Поль и Тифензее (Poll Н. und Tiefensee W., 1907) произвели сравнение гистологии семенников двух групп бесплодных гибридов: 1) реципрокных гибридов домашней утки × мускусной утки и 2) группы гибридов канарейки
с чижом, овсянкой, щеглом и зеленушкой.
Семенники гибридов как Cairina
♂ × ♀ Anas, так и Anas
♂ × ♀ Cairina (отцовский вид упоминается первым, материнский — вторым) вне брачного периода неотличимы от семенников родительских видов. Точно так же сходны они и при грубом гистологическом сравнении. В обоих случаях видны тонкие канальцы с относительно широкими просветами, покрытые однослойным или иногда двуслойным высоким кубическим (до цилиндрического) эпителием.
Совершенно иной вид имеют
деятельные железы у родителей и у гибридов. Сперматогенез превращает клеточную выстилку. нормальных семенных канальцев в многослойный полиморфный эпителий, внешние слои которого содержат материнские клетки, внутренние — почти готовые сперматозоиды. В то же время достаточно одного взгляда на гистологическую картину семенника гибрида, чтобы заметить резкое отличие несмотря на то, что внешний вид семенников, — в частности увеличение размера в брачный период, — сходен с чистыми видами. Вместо правильно расположенного сперматогенного эпителия, который в каждом нормальном семеннике выглядит одинаково, здесь видны в широких границах вариирующие нарушения нормального строения. Просветы канальцев различны по ширине и заполнены более или менее ненормальными, порой гигантскими клетками, содержащими до 20 и более ядер. Эпителий расположен неправильно, на различных местах окружности различно высок, выглядит как бы разорванным. Тщательные исследования однако показывают, что сперматогенез идет довольно далеко от сперматогонии через стадию синапсиса до деления сперматоцитов, Но через эту стадию сперматогенез не идет ни у Cairina × Anas ни у Anas × Cairina.
У исследованных Тифензее гибридов с канарейками семенники дают разнообразную картину, примем различные гибриды образуют довольно правильный ряд. На одном конце ряда стоят семенники
3/
4 гибрида канарейки (Serinus serinus × S. canarius), которые совершенно сходны по гистологической картине с нормальными семенниками. За ними идут семенники гибрида чиж × канарейка (Chrysomitris spinus × Serinus canarius) и еще дальше щегол × канарейка (Carduelis carduelis × Serinus canarius), у которых канальцы сходны с канальцами Anas × Cairina, содержат гигантские клетки и выстланы неправильным, но многослойным эпителием. Однако для всех семенников этой группы характерно, что они содержат сперматозоиды, хотя и в различном количестве и высокой степени анормального вида (иногда лишь проспермии). Здесь таким образом сперматогенез идет до конца или почти до конца, до образования сперматозоидов, хотя и неправильной формы.
Эти и дальнейшие исследования привели Поля к убеждению, что:
1) гибридизация не вообще как-нибудь нарушает половую систему гибридов, но что эти нарушения вполне закономерны и что при равных прочих условиях
определенное гибридное скрещивание не сопровождается всегда определенным изменением нормального гаметогенеза;
2) существует целый ряд различных степеней нарушения этого нормального гаметогенеза;
3) необходимо различать физиологическое и биологическое бесплодие.
Что касается последнего пункта, то он отражает ту мысль, на которой мы останавливались выше, что предпосылки плодовитости, создаваемые данным скрещиванием, с одной стороны, и фактическая плодовитость данной особи — с другой, должны быть ясно различаемы. Так, в упомянутом выше скрещивании щегла с канарейкой. гистологически обнаруживаются в семенниках сперматозоиды хотя и в небольшом числе и неправильной формы, но все же в столь близком к норме состоянии, что можно ожидать при достаточно большом обследовании встретить такую особь, у которой окажутся и вполне нормальные сперматозоиды. Действительно Эрату (Ehrat, St., 1910) удалось найти плодовитого гибрида этого сочетания, в то время как экземпляр, изученный Тифензее, принадлежал к самой крайней минус-варианте в этом отношении.
Поэтому Поль различает «в физиологическом ряду» плодовитых и бесплодных особей и в «биологическом» (генотипическом) ряду токонотов и стейронотов, сопоставляя их в следующей схеме:
Плодовитость (Fertilität) Бесплодие (Sterilität) = Токотония
Бесплодие (Sterilität) = Стейронотия
Различие между
токонотией (τοκος — плодовитый, νοϑος — гибрид) и
стейронотией (στειροσ — бесплодный) состоит в том, что у токонотов в гаметогенезе дело доходит до образования гамет, хотя бы нормальные гаметы образовывались лишь в небольшом количестве у отдельных индивидуумов. Наоборот, у стейронотов дело
никогда не доходит до образований гамет, останавливаясь на одной из более ранних стадий гаметогенеза.
Вместе с тем хотя бесплодными могут быть и токоноты и стейроноты, но по Полю бесплодие токонотов «индивидуально, относительно и факультативно», в то время как бесплодие стейронотов — обще, абсолютно и облигатное.
Плодовитость и бесплодие в «физиологическом» смысле слова не имеют какой-либо резкой границы. Как при чистом разведении, так и при гибридизации в зависимости от условии или идет нормальное размножение или возникают различные степени бесплодия, вплоть до полного. Наоборот, между токонотами и стейронотами по Полю существует резкая граница, и он неоднократно говорит об отсутствии между ними переходов, о «двух константных типах», «двух основных классах», и что переходные формы между этими классами еще не наблюдались (что конечно спорно).
Что касается стейронотов, то они также могут быть разделены на группы в зависимости от того, на какой стадии останавливается гаметогенез. В последнем Поль обращает внимание на три последние митоза (рис. 55):
 Рис. 55. Схема сперматогенеза (по Полю).
Рис. 55. Схема сперматогенеза (по Полю).
1) последнее деление сперматогоний, в результате которого образуются сперматоциты 1-го порядка;
2) первое деление созревания, с синапсисом, дающее сперматоцитов 2-го порядка;
3) второе деление созревания, дающее сперматиды.
Гаметогенез токонотов благополучно проходит все три деления, а у стейронотов останавливается либо на 2-м, либо на 1-м, либо еще раньше.
Поэтому полная классификация Поля приобретает такой вид:
1, Токоноты (тримитозные):
а) плодовитые (особи),
б) бесплодные (особи).
2. Стейроноты:
а) димитозные бесплодные,
б) мономитозные бесплодные,
в) апомитозные бесплодные.
Эту классификацию Поль наполняет богатым содержанием исследованных нм гибридов птиц, отмечая при этом, что плодовитые токоноты встречаются чаще бесплодных, а все вместе — чаще стейронотов, при этом нужно помнить, что гибриды, исследованные Полем, получались в зоологическом саду естественным спариванием, без искусственного осеменения.
В группу
токонотов попадают следующие гибриды:
Сем. Phasianidae — фазановые:
1. Syrmaticus Reevesi
♂ × ♀ Phasmnus colchicus (королевский × охотничий)
2. Phasianus Sbmmeringii
♂ x ♀ Syrm. Reevesi (земмерингов × королевский)
3. Chrysolophus pictus ♂ ×
♀ Syrm. Reevesi (золотистый × королевский)
4. Chrys. pictus ♂ ×
♀ Phas. colchicus (золотистый × охотничий)
5. Gennaeus nyclhemerus ♂ ×
♀ Chrys. pictus (серебристый × золотистый)
6. Genn. nyctheinerus ♂ ×
♀ Phas. torquatus (серебристый × кольчатый)
7. Phas. versicolor ♂ ×
♀ Syrm. Reevesi (пестрый × королевский)
Сем. Anatidae — утиные:
а) подсем. Anatinae — утки
8. Tadorna tadorna
♂ x ♀ Casarca tadornoides (пеганка × казарка новозеландская)
9. Mareca penelope ♂ ×
♀ Chaulelasmus streperus (свиязь × серая утка)
10. Chaul. streperus ♂ ×
♀ Dafila acuta (серая утка × шилохвость)
11. Paecilonelta bahamensis ♂ ×
♀ Nettion hrasiliense — (богамская утка × бразильский чирок)
12. Nettion torquatum ♂ ×
♀ Nettionbrasiliense (красноплечий чирок × бразильский чирок)
б) подсем. Fuligulinae — нырки
13. Metopiana peposaka ♂ ×
♀ Netta rufina (пепозака × красноносый)
14. Aythya ferina ♂ ×
♀ Netta rufina (красноголовый × красноносый)
Сем. Fringillidae — вьюрки
15. Serinus serinus ♂ ×
♀ Serinus canaria
16. Spinus spinus ♂ ×
♀ Serinus canaria (чиж × канарейка)
17. Acanthis cannabina ♂ ×
♀ Serinus canaria (коноплянка × канарейка)
18. Carduelis carduelis ♂ ×
♀ Serinus canaria (щегол × канарейка)
19. Chloris chloris ♂ ×
♀ Serinus canaria (зеленушка × канарейка)
подотр. Columbae — голубиные
Сем. Peristeridae (подсем. Turlurinac — горлицы) × сем. Соlumbidae
20. Turtur turtur ♂ ×
♀ Columbalivia dom. (ropлица × голубь-турман)
21. Streptopelia risoria ♂ ×
♀ Columbalivia dom. (смеющийся голубь × голубь-турман)
К этому списку должны быть далее отнесены и все вообще известные плодовитые гибриды. С другой стороны, следует помнить, что в приведенном списке не все токоноты были плодовитыми, но лишь имели в семенниках более или менее сформированными сперматозоиды. Так как тем не менее токоноты являются наиболее интересной групной гибридов, от которых имеется больше всего шансов добиться дальнейшего потомства, то наличие в ней например столь резких сочетаний, как серебристый × золотистый фазан, представляет выдающийся интерес (рис. 56).
Стейроноты димитозные, первая стадия стейронотии, встречаются (в условиях естественного спаривания) уже реже. Так по Полю у вьюрков и голубей стейронотия еще ни разу не была установлена, хотя конечно исследованные случаи еще очень малочисленны. В семействе уток такие стейроноты встречены уже в 6 случаях и каждый раз при скрещиваниях видов, относящихся к различным подсемействам. Сюда относится известный гибрид кряквы с мускусной уткой. Представленные на рис. 57 срезы яичников двух особей иллюстрируют наблюдающуюся здесь изменчивость.
Стейроноты мономитозные с выпадением обоих делений созревания установлены Полем точно лишь в одном случае и в другом — с большой вероятностью, в третьем же случае — лишь по женскому экземпляру. Гистологическая картина их гонад показана на рис. 58–59.
Наконец
стейроноты апомитозные найдены пока лишь в единственном случае при скрещивании подсемейств (свиязь × каролинская утка). Полное недоразвитие их гонад видно из рис. 60 и 61.
Нарушение плодовитости у разных полов
Широко известно, что бесплодие, и плодовитость Гибридов разных полов могут сильно различаться. Об этом мы уже упоминали и еще будем упоминать. Так, у Saturnia pavonia × pyri самки совершенно бесплодны, самцы хоть слабо, но плодовиты, у Bison × Bos taurus самки плодовиты, самцы почти или вовсе бесплодны. То же у гибридов зебу × як и т. д. Однако здесь следует отметить, что Поль на основании изложенных выше исследований выставляет тезис «идентичности нарушений гаметогенеза у обоих полов». В качестве веских аргументов, в. пользу этого обобщения он выдвигает тот факт, что среди около 30 типов гибридов птиц, изученных им и распределенных по следующим группам: 1) токоноты, 2) стейроноты димитозные, 3) стейроноты мономитозные и 4) стейроноты апомитозные, не было ни одного случая, когда, судя по самцам, гибрида пришлось бы отнести к одной группе, а судя по самкам — к: другой группе. Однако он сам же отмечает, что распределение самок между токонотами и димитозными стейронотами невозможно, так как решающая для этой классификации стадия созревания яйца проходит вне яичника и им не исследовалась. Так как к этим двум группам относятся 26 из 30-изученных им гибридов, то доказательства его тезиса тем самым резко сужаются. Тем не менее совпадение в классификации двух последних групп заслуживает внимания. Хотя для расширенного толкования этого тезиса и нет ни фактических ни теоретических оснований, тем не менее возможно, что в классе птиц и обнаружится подобная закономерность, установить или отвергнуть которую для гибридизации конечно весьма важно.
 Рис. 36. Токонотные гибриды фазанов.
1—королевский фазан; 2 — земмерингов фазан; 3 — гибрид земм. × кор.; 4 — золотистый фазан; 5 — гибрид кор. × зол.
Рис. 36. Токонотные гибриды фазанов.
1—королевский фазан; 2 — земмерингов фазан; 3 — гибрид земм. × кор.; 4 — золотистый фазан; 5 — гибрид кор. × зол.
Нарушение плодовитости в реципрокных скрещиваниях
Точно так же Поль выдвигает тезис и об «идентичности нарушений гаметогенеза в реципрокных скрещиваниях».
 Рис. 57. Разрез через яичник в брачный период гибрида мускусной утки и кряквы (по Полю).
Рис. 57. Разрез через яичник в брачный период гибрида мускусной утки и кряквы (по Полю).
Ввиду важности для преодоления бесплодия этого вопроса остановимся на нем несколько подробнее. Несомненно, что для этого обобщения Поля еще меньше оснований, и известен ряд прямо противоположных случаев. Стоит сравнить например различную судьбу реципрокных скрещивании сильнополых и слабополых рас непарного шелкопряда в опытах Гольдшмидта (см. ниже, стр. 200), в которых водном случае получались самки нормальные, а в другом — интерсексуальные, совершенно бесплодные. Но даже и для птиц это положение не только нельзя еще принять, но самим же Полем добыты и противоположные данные, прекрасным примером чего являются упоминавшиеся уже реципрокные скрещивания мускусной утки × домашней.
 Рис. 58. Разрез через семенник в брачный период гибрида чилийской свиязи и чилийской шилохвости (по Полю).
Рис. 59. Разрез через яичник брачного периода гибрида чилийской свиязи и чилийской шилохвости (по Полю).
Рис. 58. Разрез через семенник в брачный период гибрида чилийской свиязи и чилийской шилохвости (по Полю).
Рис. 59. Разрез через яичник брачного периода гибрида чилийской свиязи и чилийской шилохвости (по Полю).
Гибриды — дочери Cairina
♂ × ♀ Anas имеют совершенно недоразвитый яичник, что подтверждено многократно получавшимися гибридами этого типа, от которых ни разу не удалось получить ни одного яйца. Наоборот, реципрокные гибриды Anas
♂ × ♀ Cairina несут яйца нормального строения, но несколько меньших размеров. Развития их впрочем не удалось получить.
 Рис. 60. Разрез через семенник в брачный период гибрида каролинки и свиязи (по Полю).
Рис. 61. Разрез через яичник в брачный период гибрида каролинки и свиязи (по Полю).
Рис. 60. Разрез через семенник в брачный период гибрида каролинки и свиязи (по Полю).
Рис. 61. Разрез через яичник в брачный период гибрида каролинки и свиязи (по Полю).
Яичники этих гибридов сильно отличаются от реципрокных, содержат мелкие и крупные желтки, хотя размеры этих яичников никогда не достигают даже приблизительно нормальных размеров чистых видов. Но, с другой стороны, они никогда не редуцируются до степени «печоночно-коричневых лепешечек» с очень редкими, часто лишь под микроскопом различимыми яйцами, что типично для гибридов Cairina × Anas. Наиболее дегенерировавшие яичники Anas × Cairina и наилучше развитые яичники Cairina × Anas несколько приближаются друг к другу по внешнему виду и по гистологическому строению, но тем не менее — различия остаются столь ясными, что одного взгляда на яичник достаточно, чтобы определить, с какого типа гибридом мы имеем дело (Poll и Tiefensee, 1907, Соколовская, 1934).
Возможно, что с точки зрения классификации, данной Полем, описанные реципрокные гибриды попадают в одну группу димитозных стейронотов. Но конечно совершенно невозможно утверждать, что степень нарушения гаметогенеза в этих реципрокных скрещиваниях одинакова. Она несомненно различна, и то, что в этом случае различие не перешло за грани группы, никак не дает права обобщать этот и очень немногие другие избранные Полем случаи в гибридологическую закономерность. Она в частности может иметь место только тогда, если ни Y-хромосомы ни цитоплазмы (строение яйца) не имеют никакого значения и баланс М- и F-генов. между половыми хромосомами и аутосомами распределен, у обоих видов совершенно одинаково.
Бесплодие в F2 и дальнейших поколения гибридов
Судьба плодовитости в дальнейших поколениях гибридов весьма поучительная и дает возможность осветить и проблему бесплодия первого поколения. Здесь прежде всего мы встречаемся с большим разнообразием явлений, и представители F
2 и других поколений могут оказаться и более и менее плодовитыми, чем их непосредственные родители, особи F
1. Кроме того естественно результаты оказываются различными в зависимости от того, получается ли F
2 от скрещивания гибридов F
1 друг с другом или же получается F
b от скрещивания F
1 с одним из исходных видов. Наконец играет повидимому роль и то, с каким именно из исходных видов ставится обратное скрещивание. Здесь следовательно возможны 7 различных скрещиваний и 5 реципрокных, т. е. всего 12.
К сожалению ни одно из межвидовых скрещиваний до сих пор не было изучено по всем этим сочетаниям, большей же частью — лишь по 2, 3, 4.
Хороший пример постепенного повышения плодовитости самцов-гибридов в ряде последовательных обратных скрещиваний мы имеем в опытах Киша с гибридами гаял × тур. В/скрещивании в Галле все самки были плодовиты, а самцы бесплодны, но бесплодие здесь уже не носило абсолютного характера, а все время рядом с бесплодными попадались и плодовитые самцы. По Натузиусу (1911) отношение тех и других. было в разных поколениях таким:
Среди
1/
2 кровных 6 бесплодных 1 плодовитый 14 % плодовитых
Среди
1/
4 кровных 4 бесплодных 5 плодовитых 55 % плодовитых
Среди
1/
8 кровных 1 бесплодный 4 плодовитых 80 % плодовитых
Обратный. пример мы имеем в различных скрещиваниях бабочек. Так, Гаррисоном (1916), а затем Мейзенгеймером (1922, 1924) было произведено скрещивание двух пядениц Biston pomonarius × Biston hirtaiius. В первом поколении самцы оказались плодовитыми в скрещиваниях с самками родительских видов. Было получено 159 ♂ и 56 ♀. Однако только самцы явились действительными представителями своего пола, самки же или уродцами или как правило гермафродитами, у которых были развиты (но не вполне) обе системы половых органов (рис. 62). Кроме того самки были и мало жизнеспособны, что видно из того, что их появилась лишь треть по сравнению с числом. самцов. Ясно, что эти самки-гермафродиты были нацело бесплодными.
Цитогенетика бесплодия гибридов
В проблеме бесплодия гибридов можно различить два вопроса: 1) цитологический механизм бесплодия и 2) гено- и фенотипические причины бесплодия.
Рядом прекрасных исследований, произведенных прежде всего на бабочках (Federley Н., 1913; Pariser, 1927; Federley, 1915; Milari, 1917; Harrison a. Doncaster, 1919, и др.) было показано, что бесплодие гибридов бабочек F
b, F
2 непосредственно связано с хромосомными неправильностями в процессе гаметогенеза у гибридов. Эти неправильности сводятся прежде всего к тому, что у гибридов либо вовсе не происходит конъюгации хромосом при редукционном делении либо она имеет место лишь между некоторым хромосомными парами.
 Рис. 62. Половой аппарат трех бабочек Biston.
Справа — нормальной самки, слева — нормального самца, в середине — гибрида Fb. Женские части — белые, мужские — покрыты точками; bc — bursa copulatrix; kdr — придат. железы; Ig — яйцеклад, od — яйцевод; оv — яичник; rs — семяприемник; v — вагина; de — ductus ejaculatnrius, sb — семенные пузырьки; sp — семенник; vd — семяпроводы; р — пенис; uc — ункус; va — вальвы (по Мейзенгеймеру).
Рис. 62. Половой аппарат трех бабочек Biston.
Справа — нормальной самки, слева — нормального самца, в середине — гибрида Fb. Женские части — белые, мужские — покрыты точками; bc — bursa copulatrix; kdr — придат. железы; Ig — яйцеклад, od — яйцевод; оv — яичник; rs — семяприемник; v — вагина; de — ductus ejaculatnrius, sb — семенные пузырьки; sp — семенник; vd — семяпроводы; р — пенис; uc — ункус; va — вальвы (по Мейзенгеймеру).
Федерлей (Federley Н., 1913), скрещивая три вида бабочек Pygaera curtula, P. pigra и P. anachoreta, обнаружил, что у гибридов полученные от разных родителей хромосомы как правило не конъюгируют друг с другом и поэтому в гаметы переходит не редуцированное, а полное или почти полное число хромосом.
Паризер (Pariser, 1927), обработавшая цитологически совместные с Гольдшмидтом опыты по скрещиванию Saturnia pavonia × S. pyri. обнаружила те же явления. У S. pyri гаплоидное число хромосом 30, у S. pavonia — 29. У гибридов же в делениях созревания у самцов в экваториальных пластинках насчитывается не 29 и не 30 и не 59 (30+29) хромосом, но число, колеблющееся между 45 и 51, и частые неправильности приводят к очень пониженной плодовитости, падающей ниже 1 %. Возможно поэтому, что лишь полнодиплоидные сперматозоиды с 59 хромосомами оказываются способными к оплодотворению.
Совершенно такие же явления обнаружил Федерлей (1915) в третьей группе бабочек — у вилохвостов или гарпий (Cerura vinula × С. erminea), где число хромосом соответственно 21 и 28. На пластинках в делении созревания видно, что лишь часть хромосом вступает в конъюгации, давая биваленты (и может быть, судя по величине, даже триваленты), почему число хромосом в этой стадии несколько меньше, чем 21+28 = 49, например 34, 46 (рис. 63).
 Рис. 63. Частичная конъюгация хромосом у F, Cerura vinula × С. erminea:
Слева чистые виды: V — vinula, E — erminea, остальные — 6 гибридов. Числа — число хромосом в пластиках гибридов видны уни-, би- и триваленты (по Федерлею).
Рис. 63. Частичная конъюгация хромосом у F, Cerura vinula × С. erminea:
Слева чистые виды: V — vinula, E — erminea, остальные — 6 гибридов. Числа — число хромосом в пластиках гибридов видны уни-, би- и триваленты (по Федерлею).
Таким образом в результате нарушения процесса конъюгации гибридные гаметы оказываются: 1) диплоидными, содержащими полные гаплоидные наборы хромосом обоих видов (лучше — дигаплоидные) и 2) субдиплоидные, содержащие неполное суммарное количество хромосом обоих видов в случае либо частичной конъюгации либо неправильности митоза.
К чему приводят эти неправильности? Для плодовитости самих гибридов. F
1 наибольшее значение имеет то, что гаметогенез идет неправильно и образующиеся часто в большинстве субдиплоидные гаметы оказываются нежизнеспособными или неспособными к оплодотворению.
Если бы образовывались только диплоидные гаметы, то это могло бы не снизить плодовитости F
1, но зато эти диплоидные гаметы имели бы решающее значение для плодовитости следующих поколений. В самом деле у бабочек хромосомный механизм полоопределения таков, что самец имеет две Х-хромосомы при диплоидном наборе аутосом, а самки — один Х при диплоидном наборе аутосом. Обозначив аутосомы буквой А, имеем (не обращая внимания на Y-хромосому):
ХХАА — самец, ХАА — самка
Гаметы самца все одинаковы — ХА
Гаметы самки двух типов — ХА и А.
В случае отсутствия редукции гаметы самца будут ХХАА и, оплодотворив оба типа яиц, дадут такие зиготы:
ХХАА+ХА = ХХХААА; ХХАА+А = ХХААА.
Первая категория зигот будет триплоидными самцами; триплоидными — потому, что в них гаплоидный набор ХА повторен трижды, Самцами — потому, что у них пропорция 3Х:3А = 1Х:1А, т. е. такая же, как и у нормальных диплоидных самцов, где 2Х:2А тоже равняется 1X:1А.
Вторая же — категория не будет ни самцом ни самкой, так как у нее имеется пропорция 2Х:3А, в то время как у самок должно быть 1X:2А, а у самцов 2Х:2А. Если выразить пропорцию, приравняв всюду к 2Х, то легко заключить, что эта категория зигот будет стоять между самцом и самкой, т. е. будет интерсексом.
2Х:2А — самец
2Х:3А — интерсекс
2Х:4А— самка
Таким образом триплоиды могут быть либо самцами (ХХХААА), либо интерсексами (ХХААА), либо наконец, рассчитывая теоретически, сверхсамками (ХААА). Настоящих самок здесь быть не может.
При обратном скрещивании F
1×P мы получаем таким образом в случае образования диплоидных гамет вместо самок бесплодных интерсексов. Но и триплоидные самцы тоже не могут нормально размножаться, так как триплоидный набор хромосом не может быть правильно распределен в редукционном делении на две части. Правда, хромосомы-одного вида здесь в диплоидном состоянии, и они могут дать правильную редукцию. Но зато третий набор будет распределяться самым Неправильным образом, почему большинство гамет будет опять нежизнеспособным. Даже образование диплоидных гамет будет затруднительнее, чем в F
1. Если следовательно в скрещивание вводятся два вида, хромосомы которых оказываются неспособными давать конъюгацию с хромосомами другого вида, то это приводит к самому тяжелому расстройству плодовитости и обрекает F
1, почти на полное бесплодие несмотря на то, что по обычным зоотехническим расчетам такие гибриды будут уже
3/
4 крови, т. е. стоят ближе к чистому виду, чем стояли
1/
2-кровные F
1.
Ботаники однако на базе подобных случаев достигли интереснейших результатов, получая не F
b, a F
2. В самом деле у растений здесь возможно такое продолжение, при котором и самец и самка образуют диплоидные гаметы такого строения:
ХХ´АА´ × ХХ´АА´ (гибриды)
ХХ´АА´ × ХХ´АА´ (их диплоидные гаметы)
ХХХ´Х´ААА´А´ (зигота)
Образуется форма тетраплоидная, или правильнее дидиплоидная (амфидиплоидиая), в которой собраны вместе оба полных диплоидных набора хромосом обоих видов. — Как показал опыт, такие дидиплоидные формы оказываются вполне жизнеспособными, часто мощно развитыми и притом вполне плодовитыми, так как они получают возможность давать вполне правильную конъюгацию. благодаря повторности гомологичных хромосом:
ХХХ´Х´ААА´А´ → ХХ´АА´/ ХХ´АА´
Гаметы получаются снова дигаплоидными. (ХХ´АА´), и снова дают при оплодотворении дидиплоидпую форму. Возникает морфологически и физиологически «новый вид».
Прекрасный пример этого мы имеем в полученном Карпеченко гибриде редька × капуста, гибриде Primula kewensis и- др.
К сожалению в такой форме этот путь для зоологов закрыт, ибо гетерозиготный пол не может, дать необходимых для этого гамет. Здесь у животных, в частности у бабочек, схема будет такой:
ХХ´АА´ × ХАА´ (гибриды, самец и самка)
ХХ´АА´ × ХАА´ (диплоидные гаметы)
ХХХ´АА´АА´ (зиготы)
Этот потомок окажется всего лишь новым типом интерсекса, имея пропорцию 3Х:4А. Сопоставляя с предыдущими, паходим его место:
12Х:12А, или 2Х:2А самец
9Х:12А, или 3Х:4А интерсекс
8Х: 12А, или 2Х:3А интерсекс
6Х:12А, или 1X:2А самка
Причины бесплодия гибридов
Почему большинство гибридов бесплодно?.На этот вопрос надо отвечать как с точки зрения физиологических причин бесплодия конкретных гибридов, так и с исторической точки зрения. Это важно подчеркнуть потому, что с последней точки зрения вопрос ставится как раз обратно: почему не все гибриды бесплодны?
Остановимся сначала на втором вопросе. Легко показать, что существование гибридов в природе потому прежде всего и возможно, что они бесплодны. Выше мы видели, что наши тетерев и глухарь систематически дают гибридов друг с другом. Что было бы, если бы эти гибриды оказались плодовитыми? Началось бы систематическое перебалтывание генофондов обоих, видов в один. Оба вида как изолированные систематические единицы исчезли бы, а вместе с тем и исчезла бы возможность возникновения гибридов. «Гибриды» существуют только потому, что они бесплодны.
Но почему же тогда не все гибриды бесплодны? На этот вопрос ответить труднее и можно ответить лишь предположительно. Получения плодовитых гибридов можно ожидать там, где они не угрожают самостоятельности материнских видов. Например можно ожидать появление плодовитых гибридов между видами, изолированными географически (если эта изоляция конечно достаточно древняя), и между видами, изолированными экологически. Можно ждать их (при некоторых условиях!) в случае резких различий в размерах и некоторых других признаках родителей, которые исключают возможность естественного возникновения гибридов. Так как в естественных условиях в подобных случаях гибриды возникнуть не могут, то оба вида и могут существовать несмотря на то, что их гибриды плодовиты.
В некоторых случаях однако мы имеем плодовитых гибридов без наличия изоляции между ними. Таковы например фазаны, у которых целые области в стыке ареалов заняты гибридами. Эти состояния по нашему мнению временные. Вероятно виды недавно встретились, будучи ранее изолированы друг от друга, и образовали между собой область смешения. Рано или поздно один вид до конца продифундирует в другой, — и виды как самостоятельные исчезнут, а вместе с тем исчезнут и «гибриды», превратившись просто в полиморфную популяцию.
Одним из наиболее поучительных примеров такого перемешивания встретившихся видов является пример черной и серой ворон. В настоящее время серая ворона занимает сплошь обширную территорию от Средней Германии, Дании и Шотландии до Енисея. На западе и на востоке она встречается с черной вороной, распространенной и в Америке. В месте встреч обеих форм проходит замечательная полоса смешения, наполненная всеми переходами от серой к черной. Эта полоса шириной в 75—160 км пересекает Шотландию, Данию, Германию, огибает Альпы (рис. 64), а в Сибири идет вдоль Енисея, Алтая и где-то через Казакстан.
 Рис. 64. Карта гибридизации черной (Ч) и серой (С) ворон; черные полосы — зона гибридизации (по Мейзе).
Рис. 64. Карта гибридизации черной (Ч) и серой (С) ворон; черные полосы — зона гибридизации (по Мейзе).
Надо предполагать, что в ледниковый период оттесненная на юг ворона, бывшая единым видом, распалась на изоляты, один из которых превратился в серую ворону.
Мейзе, автор обширной монографии, открыто опираясь на Дриша и Унгерера, принимает, наоборот, что как западные, так и восточно-азиатские вороны параллельно и притом в порядке, «регуляторной способности» превратились в черных (Meise, 1928), но это явно маловероятно и имеет специфический привкус идеалистического насилия над наукой. По окончании ледникового периода вороны, расселяясь на север, встретились по указанным границам друг о другом, и так как гибриды их оказались вполне плодовитыми, то и началось перемешивание обеих форм. Замечательна медленность этого процесса. За последние 100 лет ширина полосы смешения заметно не изменилась. Если принять, что встреча произошла например 5000 лет назад и достигла ширины в 150 км, то для полного перемешивания обеих форм потребуется примерно около 100 000 лет.
История Палеарктики создала повидимому не один подобный случай. За последние десятилетия рядом выдающихся русских орнитологов (Зарудный, Мензбир, Сушкин) было описано около 10 новых видов сорокопутов из узкой полосы, идущей по широте Балхаша. Но в 1929 г. сам Сушкин убедился, что эти виды являются лишь продуктами гибридизации, идущей вдоль полосы встречи двух широко распространенных видов — европейского жулана (Lanius collurio) и азиатского буланого сорокопута (L. isabellinus, phoenicuroides). Сушкин обнаружил ясные следы менделистической комбинаторики, что можно легко видеть на материале Зоологического музея Академии наук, любезно продемонстрированном нам Б. К. Штегманом. Сушкин отмечает в области Тунгузки встречу двух видов дроздов Turdus atrogullaris и Т. nificollis, перемешавшихся настолько, что гибридные комбинации преобладают над основными видами. По Штегману (Stegmann, 1928) аналогичный процесс перемешивания двух форм горихвосток проходит вдоль Кавказа.
Эти примеры показывают нам, как благодаря достаточно медленному протеканию процесса взаимной дифузии генофондов двух видов некоторое время возможно сохранение плодовитых гибридов, распознаваемых в качестве таковых лишь вследствие того, что процесс дифузии еще не успел «ликвидировать» самостоятельность начавших смешиваться видов. Можно думать поэтому, что в старых фаунах такие явления должны встречаться реже, чем в Палеарктике с ее бурной послетретичной историей.
Возможно однако и еще одно условие наличия плодовитых гибридов наряду с существованием обоих родительских видов. В главе IX мы познакомимся с распространенным явлением атавизации, при котором гибриды часто обнаруживают ряд атавистических черт. В таких случаях они могут систематически вытесняться в порядке конкуренции своими более прогрессивными родителями, как в свое время были вытеснены и их далекие предки. Наконец это вытеснение гибридов чистыми видами будет особенно интенсивно тогда, когда в результате гибридизации возникают мало жизнеспособные или малоплодовитые комбинации (ср. например случай естественной гибридизации бражников, изученный Мори, 1901): Вряд ли например можно ожидать возможности конкурировать с родительскими видами гибриду колпицы и ибиса.
Эти соображения представляют для гибридагогии интерес потому, что, принимая их во внимание, мы сумеем быть может найти плодовитых гибридов как раз там, где их менее всего казалось можно бы искать (например между географически удаленными видами и родами). Конечно это ни мало не исключает возможности того, что и в этих условиях на базе общих физиологических причин гибриды окажутся бесплодными. Но шансы здесь все же иные.
Сила полоопроделяющих генетических факторов
Р. Гольдшмидту принадлежит честь чрезвычайно важного открытия, что мужские и женские полоопределяющие генотипы могут быть различной силы. Им это было экспериментально показано у бабочки непарного шелкопряда (Limantria dispar) для различных ее географических рас. но несомненно еще с большим основанием его выводы следует распространить на разные виды и роды, и на этом пути мы получаем ключ к пониманию части причин бесплодия гибридов.
Изложенные в ряде работ, начиная с 1912 г., основные факты Гольдшмидта сводятся к тому, что при скрещивании друг с другом европейских и азиатских рас непарного шелкопряда в разных случаях при различных скрещиваниях вполне закономерно возникают интерсексуальные особи, при этом без участия полиплоидии. Основными явлениями здесь будут следующие:
1. Разводимые в себе каждая географическая раса L. dispar дает нормальное в половом отношении потомство.
2. Скрещивание японской самки с европейским самцом дает также нормальное потомство.
3. Реципрокное скрещивание (японский самец и европейская самка) дает, наоборот, нормальных самцов, но интерсексуальных самок.
4. В F
2 от скрещивания японской самки × европейского самца все самки нормальны, но зато половина самцов — пнтерсексы.
5. При более дробном изучении географических рас обнаруживаются систематические различия в степени интерсексуальности потомства.
Гольдшмидт дал в общих чертах следующее объяснение этим (и множеству других) фактам. В каждой особи имеются задатки «мужественности» (М) и «женственности» (F), полоопределяющие гены, и вопрос о том, в какой пол разовьется данная особь, зависит от количественного соотношения Ми F. Если М окажется сильнее, то он подавляет действие F и возникает самец — и обратно.
Фактор М Гольдшмидт связывает с Х-хромосомой. Так как у бабочек половые хромосомы бывают в количестве XX у самцов и XY самок, то самцы имеют формулу ММ, самки — Mm. Следовательно для того чтобы шло нормальное полоопределение, необходимо, чтобы
ММ было сильнее F, тогда MMF будет самцом,
М было слабее F, тогда MmF будет самкой.
Если принять например 3M = 2F по силе действия, то это условие соблюдается.
Например F = 120, М = 80; 3М = 240 = 2F;
самец FMM = 120–160 = -40;
самка FMm = 120 — 80 = +40.
Но условие 3М = 2F может быть соблюдено при самых различных значениях М и F, а тем более, если не требовать точного соблюдения равенств 3M = 2F, что конечно нисколько и не обязательно. Отсюда возникла возможность сильных рас и слабых рас. Именно так и толкует дело Гольдшмидт, считая европейские расы слабыми, а японские — сильными
[36] (с пропорциональным действием
-их F и М).
Следующий числовой пример показывает возможность интерпретации наблюдавшихся фактов с этой количественной точки зрения.
1. Сильная раса (японская)
Фенотип
♂: F
120M
80M
80, F — М = —40 самцы
♀: F
120M
80m, F — М = +40 самки.
2. Слабая раса (европейская)
♂: F
80M
60M
60, F — М = —40 самцы
♀: F
80M
60m, F — М = +20 самки
3. F
1 (сильная ♀ × ♂ слабый)
♂: F
120M
80M
60, F — М = —20 самцы
♀: F
120M
60m, F — М = +60 самки.
4. F
1 (слабая ♀ × ♂ сильный)
♂: F
80M
80M
60, F — М = —60 самцы
♀: F
80M
80m, F — М = 0 самки
5. F
2 (сильная ♀ × ♂ слабый)
♂: F
120M
80M
60, F — М = —20 самцы
♂: F
120M
80M
60, F — М = 0 интерсексуальные самцы
♀: F
120M
80m, F — М = +40 самки
♀: F
120M
60m, F — М = +60 самки
Расчеты удачно, как видно, совпадают с экспериментальными данными. Гольдшмидт допускает также, что для интерсексуальности не обязательно полное равенство М и F-действия, т. е. F — М = 0, но возможны некоторые + и — значения в рамках F — М = ±15. Мужская интерсексуальность начинается при —15 и усиливается по мере изменения баланса вплоть до полного превращения в самку. То же и для самок.
Генетические самцы FMM / Генетические самки
F — М = —20 самцы / превращенные самцы
F — М = —15 начальная интерсексуальность / высшая степень интерсексуальности
F — М = —10 — 5 слабая интерсексуальность / сильная интерсексуальность
F — М = 0 средняя интерсексуальность / средняя интерсексуальность
F — М = + 5 + 10 сильная интерсексуальность / слабая интерсексуальность
F — М = +15 высшая степень / начальная интерсексуальность
F — М = +20 превращенные самки / самки
Затруднительным в теории Гольдшмидта является вопрос о локализации факторов F. На дрозофиле исследованиями Бриджеса было получено, что в женскую сторону развитие направляют Х-хромосомы и IV, а в мужскую сторону — III и II. Поскольку у бабочек аппарат половых, хромосом устроен обратно, то можно было бы принять, что у бабочек Х-хромосомы действуют в мужскую сторону, а аутосомы — в женскую. Однако Гольдшмидтом было обнаружено, что F определенной силы способен передаваться только по женской линии. Такой тип наследования свойственен Y-хромосоме, которая у бабочек передается от матери к дочери. Но Y-хромосома отсутствует у самцов и при присоединении F к Y-хромосоме все самцы оказались бы равными по отсутствию F, что не будет укладываться в схему Гольдшмидта.
Витши (Vitschi E., 1929) поэтому полагает, что носителем F у L. dispar является цитоплазма, которая тоже передается только самкой обоим полам. Именно такая схема принята нами в вышеприведенной таблице для пяти скрещиваний, где F всюду попадает к потомству только от матери, но ко всему потомству, как мужскому, так и женскому.
Отрицать возможность участия плазмы в полоопределении особых оснований нет. Межвидовые скрещивания, как мы видели уже неоднократно, как будто подводят пас к проблеме роли цитоплазмы в наследственности. Возможно поэтому, что у Limantria F в основном связан с плазмой. Однако у той же бабочки дальнейшими исследованиями установлено существование двух специальных аутосомных (т. е. локализованных в аутосомах) генов, влияющих на пол и обозначенных Гольдшмидтом как Т и D. Сочетанием различной силы F и М и наличием или отсутствием генов Т и D определяются разнообразные уклонения от нормального пола, какие наблюдаются в скрещиваниях различных рас непарного шелкопряда. Выводы Гольдшмидта в основном удовлетворяют огромному количеству экспериментального материала, собранного нм. Однако если случай с L. dispar является сейчас одним из немногих наилучше изученных, он несомненно не является исключением.
Поучительный пример полной стерилизации благодаря специфическому действию комбинации X- и Y-хромосом установил Федерлей (Federley, 1930) при скрещивании двух видов шелкопрядов рода Epicnaptera, куда относятся шелкопряды падубный (Е. ilicifolia) и осиновый (Е. tremulifolia). При скрещивании ilicifolia ♂ × ♀ tremulifolia возникает нормальпое гибридное потомство с нормальной пропорцией полов, плодовитое и дающее расщепление в F
2. Реципрокное скрещивание ilicifolia ♀ × ♂ tremulifolia дает гибридов тоже обоего пола в нормальной пропорции, но у самок, очень сильных и крупных, овариальные фоликулы совершенно пусты. При множестве вскрытий Федерлею не удалось найти ни одного яйца. Различие этих реципрокных скрещиваний показывает, что комбинация половых хромосом X
tY
i приводит к нарушению диференциации яйцевых, питательных и фоликулярных клеток, нормальный овогенез оказывается невозможным, и вырастают лишь овариальные трубки, остающиеся пустыми.
При скрещивании шелкопрядов-гарпий, Cerura furcula ♀ × ♂ С. bifida Федерлей получил слабо плодовитых самцов и совершенно бесплодных самок. Последние содержали незначительное количество яиц, иногда внешне нормальных. Однако цитологическое исследование показало полное отсутствие в них ядер и лишь иногда какие-то
- «обрывки» хроматина. Здесь следовательно имела место диференциация овоцитов в яйцевые и питательные клетки, и фоликулярные клетки образовывали нормальную оболочку, но затем повидимому (Ядра яйцеклеток подвергались разрушению. Эта форма бесплодия таким образом непосредственно переходит в нежизнеспособность ранних эмбриональных стадий.
Аналогичным образом Витши интерпретирует интересные отношения полов у лягушек. Так, Rana temporaria оказалась в различных географических пунктах Германии существенно различной по механизму определения пола. В некоторых областях эта лягушка имеет на всех стадиях развития, при которых возможен диагноз пола, достаточно правильную пропорцию полов 1:1. В других областях эта пропорция устанавливается лишь у взрослых, у молодых же имеется огромное преобладание женских фенотипов вплоть до 100 % в некоторых районах (Базель-Эльзас, Witschi, 1923; Иртенгаузен, южнее Мюнхена, Witschi, 1914). В этих случаях организмы являются гермафродитами, и окончательная диференцировка пола происходит при непосредственном участии внешних условий, в частности температуры. Витши дает интерпретацию этим фактам в духе Гольдшмидта, приписывая различную силу полоопределяющим факторам, распространенным географически не одинаково. В детали этого объяснения здесь можно не вдаваться.
О сходных отношениях можно говорить и у рыб. Так, у рыбки меченосца (Xiphophorus) отмечают существование различных рас. У одних половые диференцировка самцов происходят рано, и сами они в. связи с этим остаются мелкими. У других, наоборот, половая
диференцировка идет медленно, и сами они вырастают гораздо крупнее (что вероятно стоит в связи не только с генотипическими различиями, но и с внешним условиями). В 1908 г. Тумм (рыборазводчик) сообщил об опытах, которые повидимому свидетельствуют о том, что большие и мелкие самцы различаются своим половым генотипом, отражающимся и на диференциации пола у их потомков, что в связи с установленными для Xiphophorus нередкими случаями превращения женского пола в мужской приводит к сильному искажению пропорции полов.
Тум скрестил по 5 самок с одно возрастными им самцами разных размеров с такими результатами:
1. Крупные самки × мелкий самец — 85 % самцов + 15 % самок
2. Крупные самки × крупный поздний самец — 76 % самок
3. Средние самки × крупный самец — 92 % самок
4. Средние самки × мелкий самец — 55 % самцов
Здесь «крупные» самцы, которые только очень поздно развивают свои половые признаки и тем самым являются «женственными», дали огромное преобладание самок. Впрочем эти «любительские» факты должны подвергнуться дальнейшей проверке.
Здесь мы пока имели дело с материалом, полученным в пределах видов главным образом на базе скрещивания географических рас. Однако и для видовых скрещиваний мы имеем уже хорошие опыты, показывающие, как может в таких случаях нарушиться определение пола и притом в форме, отвечающей в основных чертах схеме Гольдшмидта о разной силе половых факторов.
Насколько своеобразными, могут оказаться явления полоопределения при межвидовой гибридизации, показывает исследование Коссвига (Kosswig, 1930) над гибридами Xiphophorus × Platypoecilus. Оба эти рода относятся к подсемейству Poeciliinae семейства зубастых карпов, обычных аквариумных живородок. В этом семействе различные подсемейства, различные роды имеют различный тип определения пола: у одних имеется гетерогаметичность женского пола (Platypoecilus), у других гетерогаметичен мужской пол (Lebistes). Высказанное в связи с этим Гольдшмидтом предположение, что рыбки этого подсемейства по сути являются еще гермафродитами и у них лишь начинает устанавливаться механизм хромосомного определения пола, подтверждается тем, что третий относящийся сюда род Xiphophorus имеет еще следы протогинического гермафродитизма. По Эссенбергу (Essenberg) часть молодых самок превращается в самцов, так что с возрастом наступает значительное преобладание самцов.
У аквариумистов Эссенберг, Хармс (Harms) и Витши собрали сведения, что в самцов могут превратиться даже самки, дававшие уже пометы. Эссенберг и Хармс спаривали этих превратившихся самцов с нормальными самками. По Хармсу потомство от подобного скрещивания состояло исключительно из самок, частично превращавшихся затем тоже в самцов.
Хармс объяснял это тем, что меченосец имеет самок гомогаметичных, XX. Эссенберг допускает для самок строение XY, но считает, что у меченосца вообще нет хромосомного определения пола (числовые данные его однако незначительны: он вообще считает, что и нормальное потомство все женское, с возрастом превращающееся частично в самцов). Таким образом у этого семейства несомненно еще очень малая отдиференцированность полов друг от друга, и здесь наблюдения за судьбой пола при скрещивании видов могут осветить интересующую нас проблему. Эти сведения нам помогут понять замечательные явления нарушения полов при гибридизации Xiphophorus.
Коссвиг поставил скрещивания Xiphophorus и Platypoecilus с участием сигнальных генов в половой хромосоме с тем, чтобы можно, было отличать самцов, определяемых хромосомным механизмом, от превращающихся с возрастом.
Первые поколения не обнаружили каких-либо особенностей и дали картину обычной сцепленной с полом наследственности типа Abraxas и птиц: доминантный признак. самок Platypoecilus передавался только самцам-сыновьям от скрещивания с меченосцем и всему потомству при реципрокном скрещивании. Нормально получалось и второе поколение от обратного скрещивания самцов F
1 с самками пецилии.
Если однако для такого обратного скрещивания брались самки F
1 и скрещивались с самцами меченосца, то получались лишь самцы (2 самки на 107 самцов). В детали этих скрещиваний, назначением которых было выяснить, какой тип строения пола имеет меченосец, мы сейчас входить не станем. Отметим однако следующий замечательный факт. Во втором поколении с участием сигнального гена окраски получилось следующее потомство:
Самцов окрашенных — 62
Самок окрашенных — 33
Самок серых — 5
Самцов серых — 38
В этом скрещивании ожидалось отношение примерно 2 самца окрашенные: 1 самка окрашенная: 1 самка серая, как в сцеплен-ном с полом скрещивании типа птиц. Серые самцы оказались совершенно неожиданными, и, судя по числовым отношениям, они появлялись явным образом за счет серых самок, которых появилось слишком мало по сравнению с окрашенными сестрами (5:33). Мало того. В то время как окрашенные самцы в значительной части не отдиференцировались в половом отношении и обнаружили гигантский рост, серые самцы, наоборот, созрели частично в 6–8 недель, т. е. раньше родительских видов, и остались очень мелкими (2–3 см). Те же результаты получились и в других подобных скрещиваниях.
Группа маленьких, рано созревших самцов оказалась очень типичной для второго поколения гибридов. Она появлялась также и тогда, когда в качестве отца брался самец F
1, созревший лишь очень поздно. Таким образом здесь мы имеем интересный и понятный в принципе (но не в деталях) пример появления в результате межвидовой гибридизации самцов, созревающих раньше времени и созревающих позже времени, т. е. являющихся очевидно тем, что можно назвать сверхсамцами и подсамцами. Здесь впрочем они не бесплодны, так что нарушения их пола не слишком сильны. Это стоит очевидно в связи с малым значением (малым половым балансом) половых хромосом, вероятным обилием аутосомных генов, влияющих на половую диференцировку (у меченосца следовательно особый тип полоопределения, Xiphophorus-тип без участия половых хромосом, мало еще ясный Коссвигу).
Пропорция полов у гибридов птиц
В связи с этими фактами интересно остановиться на вопросе о пропорции полов в межвидовых скрещиваниях птиц. Уже давно отмечалось, что среди гибридов птиц преобладают самцы. В предыдущей главе мы уже приводили соответствующие данные и указывали, что этот избыток самцов может быть объяснен различно: 1) большей легкостью признать гибридов среди самцов, 2) меньшей жизнеспособностью гибридных самок как гетерогаметичного пола. Наконец известное значение может иметь и то, что гибридные самки могут легче принимать облик самцов в качестве «превращенных самцов», попадая в списки самцов. Такую возможность указывает Холдейн (Haldane).
Однако в тех, правда очень скромных, материалах, в которых например при скрещивании фазан × курица участвовали сцепленные с полом гены окраски (Серебровский, 1929), еще ни разу не отмечалось, чтобы, птица, отнесенная к тому или иному полу, по этим сцепленным с полом генам оказалась бы при вскрытии определенной, в качестве другого пола.
Гайер (1909), сведя данные о поле 61 гибридов фазанов и голубей, нашел среди них только 4 самки и 10 неизвестных. Отсюда процент самцов 92,2±З,8, а если зачислить всех неизвестных в самки, то все же самцов будет 77,0±5,4. Однако Поль решительно оспаривает эти цифры, считая их прежде всего результатом подбора красивых шкурок в коллекциях и неверного определении пола. Например Гайер среди 13 гибридов петух × фазан не нашел ни одной самки, а Поль среди 17 исследованных нм при помощи вскрытия нашел 10 ♂ и 7♀, или 58,8±12 % т. е. нормальное число самцов. При этом он отмечает, что по внешнему виду различить живых было очень трудно и даже при осмотре их при вскрытии оставались сомнения.
Однако в некоторых случаях самцы действительно резко преобладают. Среди гибридов шпорцевый гусь × утка все вылупившиеся и даже задохлики были самцами. Среди известных до сих пор десятков двух гибридов петух × цесарка только в 1919 г. у Поля вылупились первые самочки.
Противоречие полового отбора
Здесь следует изложить некоторые теоретические соображения о диалектике эволюции пола, которые заставляют нас считать, что различие количественных соотношений М и F факторов очень широко распространено. Эти наши соображения мы уже имели случай кратко изложить в 1930 г. в докладе на Киевском съезде зоологов. До сего времени не обращалось достаточно внимания на то, что эволюция пола связана с одним очень важным противоречием, которое действительно должно приводить к тому, чтобы у каждого вида, у каждой расы уровень М и F (разумея под ними группы генов или гены и плазму — безразлично) приобретал специфический характер.
 Рис. 65. Схема среды для ♂ ♂ (слева) и для ♀ ♀ (справа).
Рис. 65. Схема среды для ♂ ♂ (слева) и для ♀ ♀ (справа).
Противоречие это состоит в том, что
всякое приспособление одного пола к другому неизбежно связано с саморазрушением этого приспособления. В самом деле замечательно, что по линии половых приспособлений вид вынужден приспосабливаться одновременно к двум диаметрально противоположным условиям. Для каждого пола, взятого в отдельности, окружающая среда состоит из двух компонентов: 1) части среды, общей для всего вида, и 2) части, специфической для данного пола. В этой второй части основную роль играют особи противоположного пола. Схематически это окружение, эту среду можно изобразить так, как на рис. 65, где кружками обозначены элементы среды, общие для обоих полов.
Для эволюции собственно половых особенностей, организма особи противоположного пола несомненно должны играть одну из решающих ролей. Таким образом каждый вид в лице самцов должен приспособляться к требованиям женской среды, а в лице самок — к требованиям мужской среды, а вид в целом должен приспосабливаться к двум прямопротивоположным требованиям.
Вместе с тем данное противоречие тесно связано с механизмом наследственности, именно с тем, что наследственные изменения, возникающие в одном поле, неизбежно в виде генов попадают к другому полу (исключением является лишь Y-хромосома, способная облегчить это противоречивое положение).
Чтобы показать, каким образом всякое половое приспособление неизбежно связано
с саморазрушением, разберем такой пример. У кур, где для осуществления полового акта петух должен произвести некоторое насилие над самкой, одним из полезных половых приспособлений является несколько большая величина петуха по сравнению с курицей. Пусть в каком-либо петухе произошла мутация (или комбинация), сделавшая его несколько более крупным. Оказавшись тем самым более приспособленным к половому акту, он оставит потомство, наследующее его полезное приспособление. Но на самом деле этот наследственный прирост согласно с механизмом хромосомной наследственности петух передаст одинаково как своим сыновьям,
так и дочерям. Это последнее подчеркнутое нами обстоятельство сразу же разрушает все приспособление, так как в этом новом поколении никакой более резкой разницы между самцами и самками уже не будет, она не сохранится. Возникшее приспособление тем самым через механизм наследственности само себя разрушило. Но вместе с тем генотип нового поколения не остался неизмененным, в нем наследственный рост увеличился за счет отобранных генов. Но и самцы и самки стали крупнее, и половому отбору снова предстоит отбирать теперь уже более крупных самцов.
Таким образом
через самцов вид должен будет непрерывно накапливать гены, усиливающие рост,
Совершенно обратно будет итти отбор через самок. Здесь будут отбираться, наоборот, гены, тормозящие рост, способные как бы нейтрализовать накопление генов крупного роста (если сам по себе крупный рост не будет, полезен для вида в целом). В общем итоге половая разница в росте между самцами и самками может сохраняться или сохранится более или менее неизменной, но-генотип вида должен будет непрерывно эволюционировать, накапливая через самцов гены — усилители роста, а через самок — гены, тормозящие рост.
Совершенно так же можно рассуждать применительно ко всяким генам, различающим самцов и самок. Пусть это, будут в общей форме гены М и F. Если у самца произойдет мутация, повышающая действие М, то сам он может оказаться лучшим самцом, как и его сыновья. Но зато этот М, — попав к дочерям, окажется уже вредным, сделает дочерей «мужественными» вплоть др интерсексуальности. Корректив к этому может быть двоякий: или М у самок подвергается, отрицательному отбору и начинает элиминироваться или, наоборот, он будет стимулировать отбор плюс-вариант среди F, так как с точки зрения правильного функционирования женского пола самки строения
FM и F
+M
+
будут одинаковы.
С другой стороны, среди самок будет тенденция к отбору F
+, которые, попадая к сыновьям, будут их портить, делая «женственными» вплоть до интерсексуальности, которая у самцов может быть компенсирована либо элиминацией F
+ либо подавлением их при помощи М
+.
Таким образом вид будет находиться все время под двояким действием отбора — через самцов вид будет наполняться мутациями) типа М
+ и через самок — мутациями F
+ (или обратно — через самок М
- и через самцов F
-). Оба пола будут таким образом непрерывно, приспособляться друг к другу перестройкой своей половой специфики. Вид будет эволюционировать в сторону сильнополовости в том случае, если М
+ и F
+ будут полезными не только для пола, но и вообще для вида, например в порядке поднятия жизнеспособности, плодовитости и пр.
Если такой процесс будет итти в различных видах, то конечно отбираемые в разных видах и даже расах гены вовсе не будут одними и теми же, т. е. тождественными и даже гомологичными. Несравненно больше шансов будет за то, что у каждого вида отобранные гены будут своими собственными, специфическими, по крайней мере во многих случаях. Это мы видим особенно ясно хотя бы на примерах гольдшмидтовских генов Т и D, встречающихся у разных рас. Каждый вид таким образом, даже оставаясь более или менее сходным с другими в смысле своего полового диморфизма (например быки и коровы у разных видов Bos, Bison и др.), будет иметь свой собственный генотип, определяющий этот диморфизм, и роль Х-хромосом, Y-хромосом и аутосом может стать у них очень различной.
Какое значение эти соображения будут иметь для интересующей нас проблемы нарушения плодовитости при гибридизации? Очень большое. Следствием такого положения вещей будет почти обязательное нарушение плодовитости уже в F
1, так как, смешав при помощи гибридизации две различные уравновешенные на правильный половой диморфизм системы генотипов воедино, мы с высокой вероятностью должны будем ожидать, что эта смесь уже не обеспечит. правильного полового диморфизма, да и сами полы окажутся искаженными. При этом для каждого пола теоретически возможны три изменения: 1) либо пол нарушится в смысле сдвига по направлению к другому полу — получится интерсексуальность («подсамцы» и «подсамки»), 2) либо пол нарушится в сторону противоположную другому полу, т. е. получится
суперсексуальность («сверхсамцы», «сверхсамки»), или, как было бы правильнее, противопоставить интерсексуальности —
экстрасексуальность, 3) либо, наконец нарушится вообще половая структура в сторону, так сказать, бесполовости, не приближаясь и к другому полу (инфрасексуальность — «недосамцы» и «недосамки»). Такой случай недавно показан Добржанским и Шульцем для гибридов дрозофил.
Необходимость различения интер- и инфра — сексуальности вытекает из следующих фактов. В пределах вида Drosophila melanogaster интерсексуальные формы можно получить скрещиванием триплоидной самки с нормальным самцом. Исследование полового аппарата этих интерсексов обнаруживает у них сильное недоразвитие выводящих путей. Если бы бесплодие гибридов Dr. mel. × sim. вызывалось их интерсексуальностью, то и здесь надо было бы ожидать недоразвития этих выводящих путей. Между тем, наоборот, у гибридных самок эти части полового аппарата развиваются достаточно нормально, но зато резко недоразвиваются сами гонады. Далее колебания в степени интерсексуальности потомков триплоидных самок оказываются одинаковыми (требуется подтверждение!) как в чистом, так и в гибридном скрещивании, из чего приходится заключать, что между, по доопределяющими генами (расположенными в Х-хромосомах) Dr. simulans,п melanogaster заметной разницы (наподобие рас Lymantria) — нет (Schultz и Dobzhansky, 1933; Шапиро 1934).
Следует отметить, что бесплодие этих инфрасексуальных гибридов дрозофил не связано с неспособностью хромосом к конъюгации (Стертевант и др.), так как недоразвитие гонад начинается задолго до половозрелости и (по Керкису) уже у личинок, выходящих из яиц, мужские гонады заметно редуцированы. Скорее наоборот — как показывают исследования «межрасовых» гибридов Dr. pseudoobscura, само нарушение конъюгации хромосом может явиться следствием инфра- или интерсексуальности, так как отказываются конъюгировать и гомологичные хромосомы (в тетраплоидных участках гонад) (Dobzhansky, 1933).
Различение перечисленных выше трех типов нарушения пола для гибридагогии должно быть чрезвычайно важным, так как в зависимости от того или иного диагноза совершенно иной будет и наша борьба с бесплодием. Получив в результате гибридизации бесплодного самца, нам придется:
в первом случае усиливать в нем мужские факторы — подавлять женские;
во втором случае ослаблять мужские факторы, может быть усиливая женские;
в третьем случае может быть придется усиливать и те и другие в определенном соотношении их друг к другу.
К этому мы вернемся ниже.
Вторым важным для нас выводом будет то, что произведенное гибридизацией нарушение механизма полоопределения резко усиливается от F
1 к F
2, но и после F
2 оно может еще долго давать себя знать, прежде чем снова окажется восстановленным необходимый механизм, обеспечивающий правильный половой диморфизм.
Вернемся к примерному случаю Гольдшмидта, к скрещиванию сильной и слабой рас.
1. В самых исходных расах половой диморфизм осуществляется правильно.
♂ F
120M
80M
80, F — M = -40
♀ F
120M
80m, F — M = +40
2. У гибридов F
2 слабой и сильной расы пол чуть-чуть нарушен — самцы на границе интерсексуальности, самки, наоборот, подходят к экстрасенсам:
F
120M
80M
60, F — М = —20
F
120M
60m, F — M = +60
3. В F
2 появляются уже настоящие интерсексы вместо F, самцов:
F
120M
80M
60, F — М = —20
F
120M
60M
60, F — М = 0 интерсексы
F
120M
80m, F — М = +40
F
120M
60m, F — М = +60
4. Скрестим самцов из F
2 с сестрами. Так как здесь сестер две категории, то возможны два случая. Один из них и с самками + GO повторит собой F
2 и опять даст интерсексов; второй случай — с самками + 40 — даст:
F
120M
80M
60, F — М = —40
F
120M
80M
60, F — М = —20
F
120M
80m, F — М = +40.
F
120M
60m, F — М = +60
Здесь интерсексов уже не получалось.
5. Если скрестить самцов — 40 с самками + 40, получим правильный половой диморфизм как в чистой расе, а скрестив самцов — 40 с самками + 60, будем иметь:
F
120M
60M
80, F — М = —20
F
120M
80m, F — М = + 40
т. е. тоже правильный диморфизм полов.
Но если мы дальше скрестим самцов —20 с самками + 60, то опять получим картину F
2 с появлением интерсексов.
Таким образом механизм хромосом Х (= М) и Y (= F или плазме) должен будет давать нарушение, пока различные комбинации аутосомных генов не создадут нового баланса или пока не восстановится исходный хромосомный аппарат F
120 М
80М
80 и F
120 М
80.
Проблема преодоления бесплодии гибридов
Прежде всего необходимо констатировать, что до настоящего времени эта проблема не только не разрешена хоть в какой-либо степени, но для ее разрешения почти не предпринималось каких-либо специальных исследований. Поэтому настоящий раздел нашего изложения должен явиться своего рода программой исследования и мобилизацией некоторых материалов, которые так или иначе могут иметь отношение к этой проблеме.
Поэтому нам придется разобрать здесь фрагментарно ряд вопросов.
1. Необходимо помнить о различиях реципрокных скрещиваний в смысле плодовитости полов. Так, если в скрещиваний Гольдшмидта
♀-европейской с ♂-японским все женское потомство оказывалось бесплодным, интерсексуальным (0), то в реципрокном скрещивании
♀-японской с ♂-европейским самки (+60) оказывались вполне плодовитыми. Разница эта особенно сильна дли гетерозиготного пола: самок у птиц и бабочек, самцов — у остальных (в данном примере +60 — 0 = 60). Но и гомозиготный пол в случае участия цитоплазмы в определении пола тоже будет различным (в этом же примере — 20 + 60 = 40). При скрещивании домашних уток с мускусными в случае Cairina ♂ × ♀ Anas дочери получаются совершенно бесплодными, реципрокное скрещивание, как раньше упоминалось, дает дочерей, откладывающих яйца. Хотя и здесь налицо бесплодие, но можно ожидать, что. работая с этими самками, все же легче будет восстановить плодовитость, чем с их реципрокными кузинами. Таким образом,
удачно используя реципрокные скрещивания, можно подобрать более выгодную базу для дальнейших попыток преодоления бесплодия.
2. Этой же цели могут служить гибридагогические мосты, т. е. третьи виды, с которыми может быть скрещен каждый из интересующих нас. Если мы стремились скрестить виды
А и
В, дающие бесплодных гибридов, и каждый из них скрещивается с третьим видом
С, особенно если этот вид располагается систематически между
А и
В, то можно получить предварительно гибридов
АС и
ВС и затем скрестить либо
АС ×
ВС, либо
А ×
ВС, либо
В ×
АС, используя и здесь реципрокные сочетания
АС и
С А, ВС и
СВ и т. д. Такие мосты между видами, даже не скрещивающимися между собой, удавалось перебрасывать ботаникам.
Нужно иметь в виду и то, что, скрещивая не непосредственно гомозиготные виды
А и
В друг с другом, а заменяя их гетерозиготами АС и
ВС, мы вводим в скрещивание сразу весьма разнообразные гаметы, среди которых несомненно гораздо легче можно будет встретить комбинации, дающие плодовитые зиготы. Этот прием конечно будет иметь одинаковое значение и для проблемы преодоления как нескрещиваемости видов, так и нежизнеспособности гибридов и их бесплодия. Поэтому разработка теории и практики гибридагогических мостов и отыскание их должны несомненно занять видное место в гибридагогии. Виды, которые сами по себе могут и не представлять какого-либо интереса, могут оказаться чрезвычайно важными для нашей работы именно под углом зрения использования их и качестве мостов. Нужно иметь в виду даже возможность перекидывания не только одночленных, но и дву-, трех- и многочленных мостов, т. е. через два-три промежуточных вида. Так, если овцы и козы окажутся неспособными дать между собой гибридов прямо, можно будет перепробовать такие более или менее промежуточные формы, как кавказские козлы, тар, гривистый баран, нахур и др. Более детальное изучение систематики их приобретает с этой точки зрения особый интерес. То же можно сказать о песце или некоторых американских собаках и лисицах как о возможных мостах между собакой и лисицей (Vulpes vulpes), об использовании гибридов Anser × Cygnopsis для скрещивания с другими видами гусей и лебедей, об Anoa, как возможном мосте между буйволами, коровами и антилопами и т. д.
Обсуждая вопрос о мостах, следует иметь в виду исключительную способность некоторых видов к гибридизации. В свое время мы упоминали о двух видах рыб рода Fundulus — F. majalis и F. heteroclitus. Первый из них мало способен давать гибридов с другими видами. Наоборот, F. heteroclitus в опытах Ньюмана и др. показал исключительную способность к гибридизации. Несомненно, что именно такие виды могут оказаться чрезвычайно ценными для использования их в качестве- мостов.
Такую же роль, по по другим причинам смогут играть наши домашние животные, так как они 1) сами уже являются продуктами гибридизации и имеют родство к нескольким видам, 2) чрезвычайно разнообразны по размерам и иным признакам и 3) имеют «расшатанный» рефлексологический аппарат и поэтому легче скрещиваются и 4) имеют как правило течку в разные времена года. Они могут служить мостами между дикими видами даже тогда, когда мы. можем стремиться скрещивать именно дикие и полудикие виды друг с другом, например яка с зубром.
Придавая таким образом большое значение внедрению в гибридагогическую работу мостов, мы с тем большим сожалением должны отметить, что и настоящее время мы не имеем почти никакого фактического материала из зоологической гибридизации, заслуживающего детального рассмотрения.
Прекрасный пример таких мостов мы: находим лишь по данным Поля у уток (Poll, 1910). Гибриды свиязи (Mareca penelope) с шилохвостью (Dafila acuta) бесплодны, и их гонады сильно редуцированы. Однако как свиязь, так и шилохвость дают плодовитых, гибридов с серой уткой (Cliaulelasmus streperus). Весьма вероятно, что скрещивание (М. × Ch.) × (Ch. × D.) даст плодовитых гибридов, и будет перекинут мост от Mareca к Dafila.
Здесь же уместно указать, что в некоторых случаях бесплодие может оказаться связанным с причинами, допускающими их преодоление генетическими методами, мутагенным воздействием рентгена и т. п. Рядом интересных работ Ленсфильда (Lancefield, 1925, 1929), Келлера (Koller, 1932). было показано, что у гибридов «рас» Dr. pseudoobscura бесплодие вызывается комбинацией генов, локализованных по концам Х-хромосомы. Удаление этих концов в результате воздействия рентгеном или перекрестом должно будет устранить бесплодие (ср. Шапиро, 1934).
3. Из предыдущего изложения мы видели, что непосредственными причинами бесплодия являются две: 1)неправильное в половом отношении развитие фенотипа гибридов, неправильное развитие или даже недоразвитие половых органов, приводящее к невозможности для гибрида нормального развития половых продуктов, яиц и сперматозоидов, и 2) неправильности деления созревания, отсутствие конъюгации между хромосомами, внесенными от различных видов в гибридную клетку. Последняя причина, как это показывают наблюдения над растениями, может действовать, тогда, когда в других отношениях половой фенотип развит вполне нормально. Поэтому можно думать, что обе эти причины бесплодия существенно различны по своей природе, и борьба с бесплодием должна быть специфической и обоих этих случаях, а большей частью — комплексной в обоих направлениях.
Можно впрочем указать и на третью причину уже кажущегося бесплодия — на то, что в результате сильного расщепления может возникать большое количество нежизнеспособных, летальных — и сублетальных комбинаций генов и хромосом, приводя к отмиранию значительного процента гамет и зигот. Но этот вопрос относится уже не столько непосредственно к бесплодию, сколько к проблеме нежизнеспособности гибридов, почему мы его и рассмотрели уже в предыдущей главе.
Отсюда следует, что при подходе к вопросу о борьбе с бесплодием гибрида необходимо прежде всего подвергнуть тщательному изучению развитие половой системы гибрида и ход у него гаметогенеза для того, чтобы определить, с каким типом бесплодия, «фенотипическим» или «мейотическим», мы имеем дело. Впрочем нужно иметь в виду, что во многих случаях будут иметь место одновременно оба типа.
4. Фенотипическое бесплодие, на преодоление которого мы в настоящее время имеем больше всего шансов, требует, как уже было указано, дальнейшей диагностики, так как здесь возможны недоразвитие (инфрасексуальность) пола и уклонение в сторону другого пола (интерсексуальность) или в противоположную (экстрасексуальность, суперсексуалыюсть). Проблема такой диагностики однако достаточно трудна. Здесь можно наметить такие пути:
a. Тщательное морфо-физиологическое изучение полового диморфизма у обоих половых типов — у исходных видов и у F
1 между ними. Если эти исследовании обнаружат, что у гибридов половой диморфизм будет выражен слабее, чем у каждого из исходных видов, то это. можно будет рассматривать как указание на известную интерсексуальность гибридов. Если, наоборот, половой диморфизм F
1 окажется еще резче выраженным, чем у исходных видов, можно будет предполагать их экстрасексуальность.
b. Большой интерес представляет перспектива биохимической диагностики сексуальности. Здесь можно указать на получившую широкую известность и подвергшуюся сильной критике реакцию Манойлопа, при помощи которой автор пытался биохимически различать кровь самца и самки. Работами ряда авторов было показано, что это различие может быть в значительной степени сведено к различной силе окислительных процессов, и например Бухгольц показал, что у растений достаточно систематическое различие между разными полами в случае двудомности дает реакция с перманганатом. В настоящее, время может быть выдвинута на разрешение задача так усовершенствовать подобный метод, чтобы иметь возможность давать
количественную оценку пола и тем самым иметь возможность различить интерсексуальность от экстрасексуальности.
c. Для той же цели может быть конечно разработан и эндокринологический метод, поскольку эндокринологи в состоянии давать количественную характеристику наличия гормонов в организме, подобно тому как Б. Завадовским и другими исследовались количества гормона щитовидной железы в разных тканях скармливанием их метаморфизирующим головастикам и оценкой силы действия гормона по степени стимуляции метаморфоза.
При наличии тем или иным путем поставленного диагноза может быть начата уже терапия. Здесь руководящая роль должна быть отведена эндокринной терапии, хотя методы ее могут быть различны. Можно предположительно наметить такие пути:
1. Гормонные инъекции гибриду.
2. Имплантации гибриду развитых желез от полноценных половых особей.
3. Трасплаптация гибридной гонады в полноценные половые особи.
Первый путь вероятно технически самый простой, однако для его успешности необходимо достаточно ясное представление об эндокринных недостатках бесплодного гибрида, т. е. весьма точная эндокринная диагностика причин бесплодия. Гипофункция половой железы может явиться результатом ее собственной аномалии, но может явиться результатом, наоборот, и гипо- или гиперфункции какой-либо антагонистической железы, например гипофиза, или еще какого-либо более сложного расстройства эндокринной системы. Поэтому может оказаться вовсе не обязательным введение в гибрида именно полового гормона. Возможно, что лучшие результаты может дать инъекция гормонных комплексов вроде гравидана (моча беременных женщин), содержащего повидимому набор различных гормонов, способный оказывать, как показал опыт, яркий возбуждающий эфект на половую систему.
Те же соображения могут быть высказаны и но второму пункту. Действительно трудно сказать, какие именно железы придется имплантировать бесплодному гибриду для возбуждения его половой деятельности. На первый взгляд может показаться наиболее целесообразной имплантация нормальной активной половой железы того же пола.
Однако, как показывают некоторые опыты, сделанные, правда, по с целью восстановления плодовитости, а с противоположной целью изменения направления развития половой железы, — успех здесь нисколько не обеспечен. Данные разных экспериментов дают противоречивые результаты.
Гринвуд (Greenvood, 1925) имплантировал семенники и яичники в эмбрионов цыплят на 7-м дне их инкубации. Но несмотря на удачу пересадок эмбрионы с обеими противоположными гонадами развивались в типичных самцов и самок. Хотя женский гормон и является «доминирующим), но в данном случае подобного влияния на развивающуюся гонаду цыпленка он оказать не сумел.
Витши (Witschi, 1927) проделал аналогичный опыт с лягушками, пересаживая головастикам задолго до начала половой дифференцировки кусочки семенников от взрослой лягушки. Яичники головастиков несмотря на наличие имплантата развились нормально.
Такие же отрицательные результаты получали Витши (1927) и Виллнер (Willier, 1927) с парабиозом (сращиванием) у головастиков и у цыплят. Срастив в ранней стадии — приблизительно на третий лень развития — головастиков лягушек попарно, Витши получил после дифференциации полов такие сращенные пары: 16 ♂♂; 17 ♂♀; 10 ♀♂; 13 ♀♀. — т. е. распределение полов, ясно подчинилось заколам вероятности и следовательно никакого влияния на пол не оказалось — в полную противоположность данным Буржа (1925), утверждавшего, что при таком же опыте с аксолотлями получались будто бы близнецы, всегда однополые: 44 ♂♂ + 36 ♀♀.
С другой стороны, общеизвестно антагонизирующее действие гормонов у млекопитающих, проявляющееся в так называемом фримартинзме, когда раздельнополые двойни у коров при установлении сообщающегося кровообращения дают постоянно нормального самца и физиологически кастрированную бесплодную самку. В слабой степени подобные явления наблюдал Витши и у сращенных головастиков.
Конечно вопрос о подавлении нормальной гонады нормальной же гонадой другого пола и возбуждение недоразвитой гонады нормальной гонадой того же пола — различные проблемы. К сожалению прямых опытов восстановления недоразвитой гонады гибрида нормальной гонадой поставлено еще не было. По М. Завадовскому, правда, в атом случае можно ожидать и прямо противоположного эфекта.
Более обещающим нам представляется обратный метод пересадок молодых гонад гибридов в нормальные в половом отношении организмы. Здесь они могут встретить совершенно нормальную для полового развития среду, и если только эта среда в состоянии определить нормальное развитие гибридной гонады, то это и произойдет. Здесь однако встречаются другие трудности: 1) разработка такого метода трансплантации, чтобы в случае удачи можно было пользоваться вырабатывающейся спермой или яйцами, и 2) затруднения с приращением трансплантата, так как здесь речь будет итти уже о гетеротрансплантации, кончающейся часто — особенно у млекопитающих— гибелью трансплантата. Возможность получения путем трансплантации дальнейшего развития гибридных гонад, естественно не развивающихся, была впервые недавно показана экспериментально Битинским-Зальц (Bytinski-Salz, 1933). Несмотря на то, что эти опыты относится к несколько своеобразному случаю, они представляют для нас значительный интерес.
При скрещивании двух видов бражника — помаренникового и молочайного (Deilephila galii × D.euphorbiae.) реципрокные скрещивания дают различный результат. Скрещивание euphorbiae
♂ × ♀ galii дает потомство обоего пола вполне жизнеспособное и плодовитое. Реципрокное скрещивание дает однако лишь самцов. Самки остаются в стадии куколок неспособными преодолеть диапаузу и погибают после нескольких перезимовок. Генетика этой сублетальности, изученной детально Федерлеем, была уже нами изложена. Она стоит в связи с «дисгармонией» Х-хромосомы euphorbiae и Y-хромосомы или плазмы galii.
Битинский-Зальц произвел трансплантации эмбриональных яичников от таких гибридных самок, неспособных к развитию (рис. 66), к нормально развивающимся самцам как обоих чистых видов, так и гибридов. В целом ряде случаев эти трансплантации удались, и яичник, попав в тело нормально развивающейся куколки, начинал и сам развиваться. Прилагаемый рисунок (рис. 67) иллюстрирует степень этого развития. Легко видеть, что оно может пойти так далеко, что в трансплантированном яичнике развиваются нормальные крупные, характерной для бражников зеленоватой окраски яйца, по мнению автора способные и к оплодотворению.
 Рис. 66. Одновозрастные яичники реципрокных F1 Deilephila euphorbiae ♂ × ♀ galii (A) и galii ♂ × ♀ euphorbiae (В) (по Битинскому-Зальц.)
Рис. 66. Одновозрастные яичники реципрокных F1 Deilephila euphorbiae ♂ × ♀ galii (A) и galii ♂ × ♀ euphorbiae (В) (по Битинскому-Зальц.)
Развитие этих трансплантированных эмбриональных яичников происходило несомненно под воздействием организма нового хозяина и притом под воздействием очевидно гуморального характера. Оно происходило синхронно с развитием хозяина. Если хозяин развивался без зимовки, развивался без зимовки и имплантат. (Нормально усиленное развитие яичников падает на последний десяток дней куколочной стадии.) Если новый хозяин оставался в стадии куколки зимовать, то без развития оставался лежать и имплантат (как показало например гистологическое изучение в январе) и пробуждался к развитию уже летом. Наконец контрольные опыты пересадок яичников чистых euphorbiae в куколки самок гибридов galii ♂ × ♀ euphorbiae показали, что эти нормальные яичники теряют способность развиваться дальше, попав в спящую куколку гибрида.
Своеобразие этого случая состоит в том, что мы в нем не имеем специфических явлений бесплодия. Недоразвитие органов размножения является результатом общей остановки развития самки. Тем не менее мы видим, что остановка развития яичника но связана со свойствами самого яичника (аналогичные опыты были поставлены и С крыльями), а зависит от каких-то иных частей организма куколки. От самки, неспособной дать потомство, путем трансплантации ее яичников в другой организм удалось таким образом получить яйца, вполне развитые и повидимому способные к оплодотворению. Совсем неплохое начало.
Надо думать, что та или иная эндокринная «починка» гибрида будет наиболее плодотворным путем преодоления бесплодия гибридов. Несмотря на то, что например по приведенным выше данным Поли о типах бесплодия гибридов птиц в большинстве случаев стейронотии гаметогенез останавливается на том или другом, делении созревания, а в частности на стадии синапсиса, — мы полагаем, что не неспособность гетерогенных хромосом вступить в конъюгацию дает бесплодие, но бесплодие в основном вызвано нарушением сексуальности фенотипа гибрида. Основным нашим аргументом является то, что по тем же данным Поля имеется весь постепенный переход от полной плодовитости к таким стадиям бесплодия (апомитозные стейроноты), при которых совершенно не развиваются гонады. У гибридов фазан × курица (Poll, 1912) и некоторых случаях недоразвитие яичников шло так далеко, что «на анатомическом место яичника» простым глазом ничего рассмотреть было невозможно, и лишь гистологический анализ показал совершенно редуцированную ткань. Здесь следовательно развитие яичника остановилось задолго до того момента, когда невозможость конъюгации хромосом могла бы дать себя знать. Поль отмечает черты сходства гибридов фазан × курица с кастратами.
 Рис. 67. Развитие трансплантированных яичников galii × euphorbiae (по Битинскому-Зальц)
Рис. 67. Развитие трансплантированных яичников galii × euphorbiae (по Битинскому-Зальц)
Такое же полное недоразвитие гонад установлено у гибридов Mareca penelope ♂ × ♀ Lampronessa sponsa (см. выше). Важно также иметь в виду бесплодность жеребцов и баранов крипторхов несмотря на то, что у них хромосомный аппарат в порядке.
Возникает однако вопрос: если дело в нарушении фенотипа, то почему гибридизация расстраивает столь сильно лишь половую железу, а не какую-либо иную? На этот законный вопрос мы считаем вероятным такой ответ.
Во-первых, как показано выше, эволюция половых признаков находится под давлением и ведущим руководством одного чрезвычайно мощного противоречия — противоречия полового подбора. Конечно и эволюция других сторон тоже противоречива — мы насчитываем не менее пяти таких противоречии. Тем не менее половые признаки в этом отношении находятся в специфических условиях, и можно ожидать гораздо более быстрой эволюции их генотипа, что подтверждают например те же опыты Гольдшмидта с географическими расами Limantria.
Однако возможно, что при гибридизации не менее часто или лишь немного реже расстраиваются и другие эндокринные железы. В частности в результате этих нарушений будет в свою очередь возникать или усиливаться бесплодие. Но мы можем иметь дело у «бесплодных гибридов» лишь с очень слабыми степенями расстройств таких эндокринных желез, как гипофиз, щитовидная и пр., просто потому, что при сильном расстройстве гибриды будут попадать в категорию нежизнеспособных. Действительно мы видим, что даже сильнейшее недоразвитие гонад, как у гибридов фазан × курица, не только не вредит организму, но даже повидимому делает его более сильным. А что сталось бы с животным, если бы у него в такой степени недоразвились гипофиз, или щитовидная железа, или надпочечник? Оно было бы совершенно нежизнеспособным уродом вроде известных телят-бульдогов, у которых на почве мутации несколько недоразвивается гипофиз.
Глава IX. Явления наследственности при гибридизации
После того как нами были рассмотрены пути получения гибридов, препятствия, стоящие на этих путях, и возможные методы преодоления этих препятствий, — мы должны более подробно ознакомиться со свойствами самих гибридов. Часть этих свойств была уже попутно обсуждена в предыдущих главах. Настоящая глава будет рассматривать вопросы о характере первого поколения гибридов и следующих поколений, об их фенотипе, об основных явлениях наследственности гибридов, об их цитологии, особенностях развития, о гетерозисе гибридов и т. д.
Особенности первого поколения гибридов
В отличие от первого поколения при скрещивании близких рас, в котором обычно мы можем наблюдать типичную картину мозаики доминантных признаков обоих родителей, для гибридов более характерно то, что они как правило получают фенотип, промежуточный между фенотипом скрещиваемых видов (рис. 68), в котором отдельные признаки родителей в чистом виде либо вовсе не удается уловить либо удается уловить лишь с большими трудностями. Известен промежуточный характер мулов и лошаков, гибридов зебу и яка, утки и шилохвости, сатурний, дрозофил и пр. Происхождение этой промежуточности в настоящее время совершенно понятно — она является не чем иным, как результатом высокой степени полигибридности гибридов и в этом отношения не отличается принципиально от межрасовых скрещиваний, так как и при скрещивании достаточно сильно различающихся рас — например далеких пород кур или собак — гибриды тоже оказываются более или менее промежуточными, по крайней мере по ряду признаков.
 Рис. 68. Промежуточная форма крыла Deilephila galii × euphorbiae.
a — galii, b — euphorbiae; пунктир — гибрид (по Битинскому-Зальц и Гюнтеру).
Рис. 68. Промежуточная форма крыла Deilephila galii × euphorbiae.
a — galii, b — euphorbiae; пунктир — гибрид (по Битинскому-Зальц и Гюнтеру).
Однако есть и еще один элемент, создающий промежуточный характер гибридов, — это то, что в случае расовых отличий мы имеем дело часто с различиями, вызванными проявляющимися мутациями, сознательно или бессознательно сохраненными человеком. Таких резких мутационных
различий между видами как правило не существует, так как повидимому естественный отбор сохраняет резкие мутационные изменения лишь в исключительных случаях. Соответственно с этим межвидовые и межродовые отличия связаны несомненно с большим числом «малых генов», т. е. таких генов, изолированно рассматриваемое действие каждого из которых незначительно или даже ничтожно.
Однако
генетика видовых различий в настоящее время еще совершенно не достаточно изучена, особенно у животных, и будет изучена только в процессе дальнейших гибридологических работ. Виды могут отличаться друг от друга весьма многообразно. Различия могут быть генными, при которых один вид имеет аллеломорф
А, а другой — аллеломорф А или
а, или вовсе не иметь реального аллеломорфа. Последнее неизбежно постулируется для далеко отстоящих видов, хотя пока и не может быть доказано экспериментально. Различия могут быть в распределении генов по хромосомам, в различном числе и относительной длине отдельных групп сцеплений, в порядке расположения генов в этих группах, что пока хорошо показано лишь для различных видов дрозофил. Сам хромосомный аппарат может быть устроен различно но общему характеру хромосом (толстые, тонкие, длинные, короткие), но особенностям полоопределяющего аппарата (XY, ZW, ХО). В некоторых случаях один вид может иметь кратное число хромосом другому виду. Причина этого уже более или менее изучена у растений, являясь результатом гибридизации. Помимо общего сложения хромосомных аппаратов двух видов в результате гибридизации имеет повидимому место и процесс удвоения отдельных частей хромосом (дупликации) без участия гибридизации. Наконец межвидовые различия могут быть связаны и с внеядерными элементами, что несомненно для растительных пластид. В последнее время накапливаются данные и о различии видовых плазм, плазмонов, хотя толкование роли плазмы весьма различно. Этот вопрос недостаточно четко ставится и методологически. В большинстве случаев вместо противопоставления плазмы ядру как двух частей клетки ядру противопоставляется под именем плазмы строение яйца как целого. Совершенно несомненно, что строение яйца, с которого начинается развитие эмбриона, должно оказывать и действительно оказывает влияние на это развитие, особенно на ранних стадиях. Но здесь идет речь именно о строении яйца как целого, включая сюда например и местоположение ядра в яйце, соотношение размеров ядра и всего яйца и т. д. Этот вопрос очевидно совсем другой, чем вопрос о том, является ли цитоплазма носителем наследственных элементов аналогично ядру или нет. В ряде случаев показано с большой вероятностью, что цитоплазма может явиться передатчиком деятельности хромосом, находившихся в яйце до редукционного деления и потом ушедших из яйца. Некоторые случаи пестролистности растений позволяют усматривать в цитоплазме (помимо пластид) и элементы, сопоставимые с хромосомными генами, хотя вопрос этот и очень далек от ясности. Межвидовая гибридизация и должна осветить подобные вопросы.
Формула, что гибриды имеют фенотип, промежуточный между родителями, слишком обща и поэтому неудовлетворительна. Более правильным будет сказать, что при сравнении признаков гибридов с признаками родителей можно различить пять различные-случаен, а именно:
1. Одни признаки оказываются более или менее хорошо повторяющими признаки одного из родителей.
2. Другие признаки оказываются более или менее хорошо повторяющими признаки второго родителя.
3. Третьи признаки оказываются более или менее промежуточными между признаками обоих родителей.
4. Четвертые признаки оказываются развитыми сильнее, чем у какого-либо из родителей, или даже новыми.
5. Пятые признаки оказываются развитыми слабее, чем у кого-либо. из родителей, вплоть до полного исчезновения.
Из более или менее одновременного присутствия всех: этих пяти групп и слагается суммарное впечатление «промежуточности» гибридов.
В качестве примера приведем полученных и детально изученных Лангом (Lang, 1909) и Клейпером (Kleiner, 1913) гибридов молюсков Helix hortensis и Helix nemoralis.
По большинству признаков, различающих родителей, гибриды носят мозаичный характер: они обнаруживают, одни признаки чисто отцовские, другие — чисто материнские. При этом видовые различия (к которым Ланг относит форму отверстия, индекс отверстия и выпуклости, в сильной степени пигментацию перистома, форму и ветвление слизистых желез и в первую очередь особенности любовной стрелы — рис. 69) и расовые (окраска раковины, комбинации полосок на ней и т. п.) ведут себя несколько различно.
 Рис. 69. Любовные стрелы улиток: сбоку (А) и в разрезе (В).
1 — Helix hortensis; 2 — гибрид; 3 — Helix nemoralis.
Рис. 69. Любовные стрелы улиток: сбоку (А) и в разрезе (В).
1 — Helix hortensis; 2 — гибрид; 3 — Helix nemoralis.
Промежуточными оказываются абсолютные размеры, длина завитка, размер последнего оборота, общая величина раковины, длина мешочка стрелы, длина стрелы, длина flagellum.
В одном признаке — в необыкновенной вздутости раковины (forma соnoidea) — большинство гибридов заметно превосходит даже более вздутую раковину H. nemoralis.
Редко оказываются промежуточными расовые признаки — полосы.
Стрелка, по которым оба вида имеют не трансгрессирующие различия, развивается у гибридов в форме, частью промежуточной, частью мозаичной.
В качестве второго хорошего и детально описанного примера приведем исследованную Гайером (Guyer, 1909) группу из 5 гибридов цесарки × лангшан (порода кур).
По весу и размерам гибриды приближались к курам, веся в возрасте трех лет в среднем 8 англ. фунтов (цесарка — 3–5 фунтов, петух-лангшан — около 10 фунтов). По общим контурам тела они были промежуточными (рис. 70). Оперение и рисунок были однако более генерализованы (атавизированы, см. ниже), чем каждый из родительских видов. Голова была одета перьями до клюва и без всяких украшений — не было ни гребня, ни сережек, ни шлема, имелись лишь слабые следы голубой окраски лица цесарок возле самых глаз, где сохранилась небольшая голая зона, если не считать прорастающих ее нежных волосоподобных перышек. Голова длинная, несколько цилиндрическая, переходящая в длинную шею, создавая несколько змеиное впечатление. Клюв по форме и окраске приближался к цесарке, но с большей кривизной. Оперение на шее и голове несколько уже и нежнее, чем у кур (у цесарок голова и шея частично голые, частично покрыты редкими щетинообразно измененными перьями). Под горлом наподобие цесарок имелись продольные висячие складки кожи с маленькими белыми перышками. Хотя все о гибридов оказались самцами, ни один не имел шпор. Крепкие ноги имели по внешней стороне слабое оперение подобно, но значительно слабее, чем у отца-лангшана (мутационный признак, см. ниже).
 Рис. 70. Гибриды цесарка × петух. (По Гайеру.)
Рис. 70. Гибриды цесарка × петух. (По Гайеру.)
Воспитавший гибридов заводчик уверял, что в молодости гибриды походили на цесарок и лишь с возрастом приобрели промежуточный характер. Сохранившиеся дольше в зоосаду в Цинциннати два гибрида и пятилетием возрасте приобрели еще более выраженное сходство с петухами, развив по паре типичных для петухов серповидных перьев в «хвосте» (кроющие хвоста).
Голос гибридов более громок и еще более негармоничен, чем у цесарок, походя в общем более на голос последних. Гибриды были все время чрезвычайно дикими и не сделались ручными несмотря на частое общение с ними и взятие на руки. Пойманные, они бились и кусались и время от времени выбрасывали из клоаки большие количества густой жидкости — явно в качестве метода защиты (ср. чаек). В связи с этим следует отметить, что вскрытие обнаружило у них сильное развитие слепых отростков кишки — более сильное, чем у обоих родительских видов.
Общий характер оперения гибридов был более или менее промежуточным, приближаясь более к цесарке; хвост хотя и не приподнимался круто вверх, как у лангшанов, но все же был восходящим и никогда не опускался вниз, как у цесарок. Некоторые перья хвоста имели одно опахало типичное для всего оперения, а другое — черное, причем на-открытых частях крупных перьев имелась тенденция развивать рисунок, а на закрытых — быть черными. Первые махи у всех гибридов были белыми (1–2–3), как обычно у цесарок. Характерная белая пятнистость цесарок исчезла, и перья имели более или менее подковообразный рисунок из черных и узких беловатых полос. Нередко проявлялся, красноватый оттенок, особенно в гриве (при скрещивании лангшанов с дикоокрашенными породами, например бурыми леггорнами, имеющими черногрудо-красную окраску петухов, красный цвет с некоторыми видоизменениями в распределении очень сильно проявляется уже у первого поколения).
Эта подковообразная разрисовка перьев по исследованию Гайера оказалась широко распространенной в группе куриных (см. ниже)
Детальный анализ более близких гибридов кряквах шилохвость, выясняющий природу «промежуточности» и сводящий ее к мозаике признаков родителей, находим в книге Науманна (Naumann).
Поведение гибридов представляет безусловно большой интерес, так как и здесь должно находить свое специфическое-выражение соединение двух разных генотипов. Здесь часто можно-отметить большую дикость гибридов и их интерсексуальное, кастратное поведение.
Дикость отмечает Гайер, как мы только что видели, для гибридов, цесарка × петух (хотя для близких гибридов цесарка × павлин Поль, наоборот, отмечает мирный характер. — Poll, 1920, 379). Чрезвычайную дикость гибридов цесарки и бойцового петуха отмечает Жуан Виларо (Guan Vilaro, 1897). Очень злыми являются также некастрированные мулы. Наоборот, гибриды льва и тигрицы были вялыми и смирными, что Ноак (Noack, 1908) ставит в связь с редукцией у них мозга (см. ниже).
Гибриды цесарках павлин необыкновенно долго ходили за своей наседкой, гораздо дольше, чем одновозрастные цесарята. В первый год они подвергались жестоким преследованиям всего птичьего двора — и павлины и цесарки одинаково гнали и били их. Сами гибриды сначала больше присоединялись к цесаркам, после — к павлинам. Изменилось и их положение на птичнике — они заняли господствующее положение. С утра до ночи бродили но лужайке и, вмешиваясь в птичьи драки, распугивали дерущихся. В длинных полетах павлинов они принимали участие неохотно. Когда цесарки подросли, в 1913 г. гибриды соединились с ними, как описывают и Гайер (1912) и Гиджи (1900), так что «самосознание» гибридов роднило их больше с цесарками несмотря на разницу в размерах.
Интересно, что на ночь зимой, гибриды забирались в сарай, куда родительские формы никогда не заходили.
По крику гибриды цесарка × павлин непохожи ни на кого из родителей. Они кричат в продолжение минуты громко, двусложно, один слог с ударением, другой без ударения — «вроде скрипения двери».
Один из гибридов «пытался подражать» павлину, распускающему хвост, и оригинально ухаживал (в возрасте лишь 4 лет) за цесаркой, распуская на короткий момент кроющие хвоста и снова складывая их. При этом он стрекотал по-павлиньи стержнями перьев, кричал на цесарку, которая убегала и взлетала на дерево. (Движения шеи, характерного для павлина, у него не наблюдали.) Так как цесарка не реагировала на ухаживание и убегала, то попыток крыть ее не было.
У гибрида чилийской свиязи с домашней уткой Гейнрот (Heinroth, 1930, 664) описывает своеобразное поведение, характерное для мускусных уток — гибриды задирают голову кверху и как бы хватают клювом воздух или «пытаются проглотить собственный язык».
Штандфусс наблюдал поведение гибридных самцов сатурнии pavonia × pyri. Им были предложены самки гибридные, обоих родителей и третьего вида spini. Удивительно, что гибридные самцы чувствовали себя вполне подходящими к самкам чистой обыкновенной сатурнии (pavonia), а эти последние принимали их как своих без малейшего сопротивления. Скрещивание происходило в нормальное для этого вида время — 3–5 час. дня. В то же время дня самцы помеси ухаживали, и за гибридными самками, но со, значительно уже меньшей охотой, хотя эти самки относились к ним Столь же благосклонно. Спаривание с самками другого родительского вида, плодовой, удавалось уже с трудом и лишь с наступлением темноты. Гибриды также «назойливо приставали» и к самкам третьего вида, spini, терновой сатурнии, но эти самки тотчас же сбрасывали гибридных самцов или убегали,
Интересные наблюдения над питанием гибридов произведены над бабочками, у которых питание часто очень специализировано на определенных видах растений,
При гибридизации шелкопрядов дуболистный (Eutricha quercifolia ♂ × ♀ тополевый (Eutricha populifolia) гибриды, так называемые Е. jolini в стадии гусениц по внешности приближаются сначала к тополевому, а позднее к дубоволистному шелкопряду. Замечательно, что в смысле пищи они, наоборот, сначала ели корм дубоволистного шелкопряда (боярышник), а затем тополевого (тополь).
Наоборот, при различных скрещиваниях бражников рядом исследователей отмечено, что они едят корм материнского вида (Deilephila euphorbiae, молочайный × D. vespertilio, D. euphorbiae × galii. - Kysela 1907, 1908). Естественных гибридов galii ♂ × ♀ euphorbiae два года подряд находил на молочае уже Mutzel (1840). Наконец гибриды gaii ♂ × ♀ vespertilio тоже выкармливались на иван-чае (Kysela, 1907). Это является естественным результатом того, что матери откладывали яйца на типичные для них растения. Если бы гибридные гусеницы оказались неспособными есть эти растения, они бы погибли и не были бы «найдены в природе».
К этому необходимо добавить
кастратный характер многих гибридов, например курица × фазан, связанный может быть с сильным недоразвитием половых органов, отсутствием проявлений половых инстинктов и склонностью в то же время к выхаживанию цыплят, что характерно и для каплунов. Этот кастратный характер при отсутствии однако характерного для кастратов петухоподобного оперения (см. исследования М. М. Завадовского с кастрацией петухов и фазанов, 1922) придает описываемым гибридам несколько более своеобразный характер — «промежуточный» сразу между четырьмя формами: обоими видами и обоими полами.
Интерсексуальный (или инфрасексуальный) характер свойственен, как говорилось в предыдущей главе, многим гибридам, почему половые признаки часто попадают в 5-ю группу признаков гибридов, накладывая на них своеобразный характер. Хорошим примером такого фенотипа может служить гибрид гаур × шортгорн, изображенный на рис. 71. Большой интерес представляет в связи с этим вопрос, не стоит ли в непосредственной связи с этим возникновение гетерозиса у гибридов, о чем будет итти речь ниже.
 Рис. 71. Гибрид гаур × шортгорн (из Плате).
Проявление мутантных признаков у гибридов
Рис. 71. Гибрид гаур × шортгорн (из Плате).
Проявление мутантных признаков у гибридов. При гибридизации диких форм с домашними со стороны последних в скрещивание обычно вводятся те или иные мутантные признаки, притом такие, которые в пределах домашних видов дают типичные менделевские явления и в первую очередь доминирование. Имеется уже довольно много указаний на то, что доминирование таких мутантных признаков достаточно хорошо сохраняется и в процессе гибридизации. Так, у крупного рогатого скота отмечалось, что доминантный ген белоголовости доминирует и при скрещивании с бизонами и- с яками. Более или менее доминируют комолость и белохребетность. Прилагаемый рис. 72 изображает мула, унаследовавшего доминантную пегость чубарой лошади.
 Рис. 72. Чубарый мул (по Лангу).
Рис. 72. Чубарый мул (по Лангу).
Подробно в этом отношении были изучены гибриды куриных (Серебровский, Гайер). Так, при скрещивании цесарок с лохмоногими лангшанами или фазанов с лохмоногими курами гибриды получают «гетерозиготную» оперенность ног, совершенно типичную и для скрещиваний внутри пород кур. При скрещивании фазанов с курами-голошейками (прекрасный доминантный признак кур) типично голошеим оказался и гибрид. Гибрид цесарки с курицей в Парижском: музее имеет типичные для кур баки, вызываемые доминантным и мутантным геном (Guyer, 1909). Гибриды цесарки с белым леггорном— петухом (Подвязная, неопубликовано) оказались белыми благодаря гену эпистатической белой окраски.
Очень поучительным оказалось скрещивание монгольского фазана с курицей плимутрок. Окраска последней по сравнению с «дикой» характеризуется тремя мутантными генами: «меланистическим» геном tifa, локализованным в аутосоме, и двумя генами, локализованными в половой хромосоме: «серебристый» ген tuge, замещающий золотистый пигмент белым цветом, и «поперечнополосатый» ген trage, вызывающий возникновение 4–5 белых полос поперек опахала.
Так как tuge и trage локализованы в Х-хромосоме, то курицей они могут быть переданы только своим сыновьям, a tifa как аутосомный ген — и сыновьям и дочерям, а в случае гетерозиготности — половине сыновей и половине дочерей. Это и произошло в действительности. Одна самка-гибрид оказалась чисто черной, другая самка — черной с коричневым крапчатым рисунком. Оба эти типа — чисто черный и крапчатый — возникают и при скрещиваниях с участием tifa в пределах пород кур.
Сыновья оказались, как ожидалось, «серебристыми» со слабо заметной поперечной полосатостью или, наоборот, поперечнополосатыми со слабо заметной серебристостью. Это тоже точно совпадает с тем, что ожидалось, и объясняется тем, что сыновья, получившие tifa, должны были дать типичную для tifa-trage окраску плимутрок, на которой tuge обычно не проявляется. Наоборот, не получившие tifa-гена имеют возможность сильно развить «серебристую» окраску по всему опахалу, почему беловатые поперечные полосы trage обычно становятся незаметными и у чистых кур.
Помимо описанного свойства ген trage имеет и еще одно, очень своеобразное, вызывая повидимому нарушение соматических митозов и частую элиминацию половой хромосомы в перьях. Обычно происходит элиминация половой хромосомы, несущей сам этот trage-ген, а вместе с ним и tuge, если он сидит в той же самой хромосоме. В результате этого процесса возникают так называемые «исключительные перья», лишенные признаков поперечной полосатости и серебристости и соответственно оказывающиеся «золотистыми» с различным рисунком, определяемым остальным генотипом.
Совершенно такие же исключительные перья были обнаружены нами и у всех самцов-гибридов фазан × плимутрок с одним возможным отличием, что число исключительных перьев было несколько больше, чем то бывает при скрещиваниях в пределах пород кур.
Рассмотренные примеры относятся к числу доминантных мутаций (по сравнению с диким Gallus bankiva). Однако и по отношению к изученным рецессивным. мутациям картина получилась та же. Так, у кур имеется рецессивная мутация «ситцевости» — atrase, выражающаяся в возникновении хорошо очерченных белых пятен на концах перьев. При скрещивании с фазанами этот признак исчез у гибридов, т. е. повел себя как рецессив и в гибридном скрещивании, указывая на наличие у фазанов соответствующего доминантного аллеломорфа. В скрещивании селезней породы хаки, имеющих рецессивную мутантную коричневатую окраску, сцепленную с полом, с черными мускусными утками ♀♀, сыновья оказались интенсивно окрашенными, а дочери более светлыми, как и следовало ожидать по схеме наследования рецессивных сцепленных с полом признаков (Соколовская, 1934).
Гибрид цесарка × белый виандот оказался окрашенным в соответствии с рецессивной белой окраской виандотов, использованных Огородним.
Совершенно такую же картину дали гибридизационные опыты с дрозофилами. В скрещивание D. melanogaster × D. simulans Стертевантом вводились самые разнообразные рецессивные и доминантные мутации, числом до нескольких десятков. Всюду оказывалось, что мутации, наследовавшиеся в качестве рецессивных в пределах чистого вида вели себя также в качестве рецессивов, т. е. не проявлялись в F
1 при гибридизации. И наоборот. Только полудоминантный ген Ваг, суживающий глаза у D. melanogaster, у гибридов проявлял себя слабее, может быть в связи с большими размерами глаз у D. simulans.
Эти факты, которые можно было бы еще пополнить, показывают, что гены, возникающие в качестве доминантных или рецессивных трансгенаций в пределах вида, ведут себя таким же образом и при межвидовых скрещиваниях. Это позволяет нам считать, что наблюдающееся иногда у гибридов неполное проявление доминантных генов зависит не от смены доминирования, но от явлений взаимодействия генов, от эпистаза и новообразований при комбинациях. Примером этого может служить наблюдавшееся нами очень слабое проявление гена trage у гибрида Gallus varius × домашняя курица. Несмотря на то, что наличие этого гена у гибрида, судя по некоторым местам оперения, было несомненно, на большинстве перьев можно было уловить лишь намеки на поперечную полосатость. Надо думать, что в генотипе G. varius имеется особый ген (гены), действующий антагонистически с геном trage, и из совместного действия этих генов-антагонистов и получился этот фенотипический признак, едва заметная полосатость. Об ослаблении проявления гена Ваг у гибридов дрозофил только что упомянуто.
Наоборот, в главе VII мы разбирали случай резкого усиления действия гена Sp. у гибридов Xiphophorus × Platypoecilus, приводящего к летальному меланозису. Усиление отмечено также для некоторых других мутантных генов у Platypoecilus. Ген R вызывает у Platy развитие красной окраски после 10-го дня. У гибридов с Xiphophorus этот же ген окрашивает мальков уже при рождении. Ген Dr. проявляется у чистых Platy лишь на 20-й день, а у гибридов F
1 и F
b × на Xiphophorus уже на 1—2-й день (Kosswig, 1933).
Если в описанных выше случаях курица × фазан, кряква × мускусная утка наследственность, сцепленная с полом, наблюдается относительно нормального и мутантного аллеломорфа, то у тех же уток и у бабочек мы находим хорошие примеры подобной наследственности и относительно двух нормальных генов. В скрещивании домашней утки с мускусной сцепленным с полом повидимому оказывается хохол мускусной утки (Соколовская). Гусеницы шелкопряда-отшельника Pygaera anachoreta темнее, чем гусеницы P. curtula. При скрещивании anachoreta ♂ × ♀ curtula гибридные гусеницы оказываются тоже темно окрашенными. Здесь таким образом темная окраска оказывается доминирующей (Federley, 1922). При реципрокном скрещивании однако возникают гусеницы двух типов — темные, дающие самцов, и светлые, дающие самок: темная окраска матери переходит к сыновьям, что отвечает XY-строению самок бабочек:
ХХ (светлый отец) → X
DY (темная мать) → XX
D (темный сын) + XY (светлая дочь)
Является ли данный ген свойственным обоим видам, в форме
D у anachoreta и рецессивной в форме
d у curtula или у curtula его совсем нет, так что гибридные самки должны быть обозначены как
DО и ген
D является в полной мере неоморфом — требует очевидно специальных исследований.
Аналогичное явление наблюдалось в скрещиваниях красноголовой и черноголовой гульдовой амадины.
Закономерности образования признаков гибридов
Атавизация
Изучение признаков гибридов позволяет установить одну очень важную закономерность, важную в теоретическом отношении, так как она дает указание на закономерность эволюционной диференциации видов, и важную с точки зрения практической, так как она позволяет до некоторой степени предсказывать свойства гибридов первого поколения.
Эта закономерность состоит в том, что гибриды получают известный
атавистический характер, выражающийся прежде всего тем, что у них обыкновенно исчезают или резко ослабляются все более специфические признаки скрещиваемых видов, т. е. признаки, в эволюционном отношении более молодые. Хотя эта закономерность не строгая, тем не менее она достаточно широка дли того, чтобы с нею надо было считаться. Является ли она свойственной всем группам животных или в особой степени некоторым, пока судить трудно, но в некоторых группах (в частности у куриных) она выражена очень резко.
Относящиеся сюда факты следующие. Различные куриные имеют своеобразные и специфические головные украшения. Так, петух Gallus имеет на голове мясистый, более или менее зубчатый гребень и сережки; цесарка имеет вместо мясистого гребня костнокожный шлем, сережки иного строения, широкие ноздри; павлин имеет оперенную голову с оригинальным хохлом из перьев, снабженных маленьким опахалом на конце более или менее голого стержня; индюк имеет голову полуголую, как и цесарка, но над клювом свисающий, мягкий, способный раздуваться отросток и такие же кавернозные шишки на шее.
Все гибриды между названными формами имеют совершенно гладкую голову, более или менее нормально оперенную (см. рис. 70 и фронтиспис). Все перечисленные выше родовые признаки, являющиеся эволюционно молодыми, так как они свойственны только каждому из данных родов и несвойственны массе куриных, у гибридов исчезают. В силу этого голова гибрида с полным правом может быть названа не «промежуточной», а атавистической. Если даже у предков куриных голова и не была именно такой, то во всяком случае несомненно, что она не имела ни гребня Gallus, ни шлема Numida, ни, хохолка Pavo, ни шишек Gallopavo.
Гребень, свойственный Gallus, исчезает (точнее крайне недоразвивается) во всех изученных до сего времени гибридизациях с цесаркой, с павлином, с фазаном, с тетеревом, и у нас поэтому имеются все основания ожидать, что и при всяких других скрещиваниях Gallus он будет также исчезать. Исключением из этого естественно должны быть лишь скрещивания с другими Gallus, у которых гребень также имеется, как и у домашнего петуха. Но здесь уже следует ожидать исчезновения тех специфических отличий в гребнях, которые еще моложе, будучи свойственными лишь отдельным видам, или имеют более специфический характер. Действительно гибриды Gallus bankiva × G. varius имеют гребень, но, насколько позволяют судить недостаточно точные описания, утрачивают видовые специфические признаки: глубокие вырезки G. bankiva, желтые пятна G. varius и пр.
Из всех куриных цесарка имеет особый и редкий серебристый тон оперения благодаря наличию лишь черного пигмента и отсутствию широко распространенного пигмента желто-коричневого. Так же своеобразен и рисунок цесарки в виде рядов округлых белых пятен. Можно было бы ожидать поэтому исчезновения этих специфических особенностей окраски у гибридов.
Действительно известные до сего времени гибриды цесарки с павлином, индюком и курицей имеют нормально развитый желтокоричневый пигмент и рисунок, в котором ряды округлых пятен заменены подковообразными рыжими полосами. Этот рисунок, подробно изученный Гайером, является характерно «атавистическим», так как он чрезвычайно широко распространен в различных группах куриных птиц и даже у более примитивных видов подсемейства цесарок-
Серебристый и золотистый фазаны оба имеют хохлы, однако совершенно различного строения. У серебристого фазана перья состоят из отдельных пушистых нитей черного цвета (при общем серебристом окрасе оперения), у золотистого — из крышеобразно сложенного опахала из крепкого стержня и рассученных бородок первого порядка золотистого цвета. Хохол гибрида состоит из нормальных перьев серебристой окраски, как преимущественно и все оперение, все специфические особенности строения перьев обоих родов (Gaenneus и Chrysolophus) нацело исчезли.
Подобные же факты можно найти и в других группах животных.
Так например в сем. лошадей зебры обладают очень своеобразной окраской из черных и белых полос. Если наклонность к полосам распространена довольно широко (черные ремни лошадей и ослов, крестообразная полоса на холке ослов, поперечная полосатость ног лошадей и ослов), то белая окраска взамен рыжей является у млекопитающих редким исключением. В соответствии с этим гибриды зебр с другими Equidae нацело теряют белую окраску и сохраняют лишь — иногда по всему телу, иногда местами — темные полосы. Возможно при этом, что древние лошади имели полосы более резко выраженные, чем сейчас, и их нынешняя окраска явилась уже приобретением последних полупустынных эпох. Расположение полос у зеброидов обычно напоминает наиболее примитивный тип зебры Греви (Dolichohippus grevii).
У гибридов як × зебу исчезают специфические видовые признаки — бахрома по бокам тела яка и горб зебу.
Подобных фактов можно было бы привести еще много для того, чтобы показать, что они превалируют над противоположными. Однако и наличие этих противоположных фактов несомненно. Так например у цесарок несколько первых маховых перьев обычно бывают белыми (как и у павлина). Гибриды цесарки с курами тоже имеют белые первые маховые (Guyer, 1909). Гибриды цесарки с павлином имеют очень удлиненные кроющие хвоста (как у павлина, но может быть и у некоторых других куриных, в том числе у Gallus). Очень эфектно появление резкого, хотя и маленького белого пятна на плече у чрезвычайно далекого гибрида тетерев × фазан (рис. 73). Совершенно такое же белое пятнышко имеется у тетерева, у глухаря и у межняка.
Блеск оперения вызывается, как известно, не пигментом, а структурными особенностями оперения. Так как эти особенности не одинаковы у различных видов и весьма специфичны, то неудивительно, что даже в том случае, когда оба родителя гибрида имеют яркий блеск, гибрид может оказаться матовым. В качестве такового Гайер приводит гибрида павлин × курица (Guyer, 1909, 744) и различных других, произошедших от блестящих, но далеких форм, например фазан × курица.
Несмотря на наличие подобных исключений — на наш взгляд они в самом деле являются более или менее исключениями — мы можем считать атавизацию гибридов действительной закономерностью, определяющей характер признаков гибридов. Какова природа этой закономерности? Действительно ли играют здесь роль «молодость» и «старость». признаков? Толкование этих явлений, как делают некоторые авторы в терминах «молодости» и «старости», граничит с явной мистикой. На самом деле, как мы думаем, вопрос здесь в том, что большинство специфических, вновь возникающих признаков оказывается связанным в большинстве случаев с накоплением рецессивных мутаций. В такой форме эта закономерность, наоборот, совершенно естественно освещается нашими современными генетическими знаниями. Действительно сейчас установлено колоссальным количеством фактов из разных областей животного и растительного мира, что рецессивные мутации возникают во много раз чаще настоящих доминантных. В любой популяции в скрытом виде, «как в губке», оказываются сохраняющимися разнообразные рецессивные мутации до тех пор, пока по условиям жизни вида та или иная из этих мутаций не окажется полезной. Тогда, подхваченная естественным отбором, она может начать быстро накапливаться и войдет в состав характерного генотипа вида. Конечно для естественного отбора, для качества приспособительного признака не имеет значения, обусловливается ли он действием доминантного или рецессивного гена, и поэтому решающим должен явиться вопрос, какая мутация будет чаще подпадать под действие отбора. Поскольку рецессивные мутации значительно. чаще и могут длительно сохраняться в скрытом виде до подходящего момента, постольку они чаще и будут подпадать под действие положительного отбора (некоторые имеющиеся здесь отличия между доминантами и рецессивами, например то, что доминанты могут подпадать под действие отбора непосредственно по их возникновении, а рецессивы — лишь после некоторого их предварительного стохастического размножения
[37], — для данного вопроса решающего значения не имеют).
 Рис. 73. Гибриды тетерев × фазан (слева) и тетерев × глухарь (справа) (ориг. с экз. Ганноверского музеи).
Рис. 73. Гибриды тетерев × фазан (слева) и тетерев × глухарь (справа) (ориг. с экз. Ганноверского музеи).
Поэтому для всякого вновь вырабатываемого признака больше шансов оказаться связанным с рецессивными мутациями, чем с доминантными. Вместе с тем понятно и то, что в определенном, различном для каждой группы форм проценте в образование новых признаков должны вовлекаться и доминантные мутации
[38]. Тем не. менее роль рецессивных мутаций остается преобладающей, почему и атавизация при гибридизации является действительно закономерностью, отнюдь не устраняемой тем, что она захватывает фенотип не нацело, не на 100 %, а лишь в меру преобладания среди специфических признаков рецессивных над доминантными.
Закономерность атавизации гибридов таким образом является производной от закономерности преобладания в процессе эволюционной дифференциации рецессивных признаков, а эта последняя — производной от преобладания рецессивных мутаций над доминантными. Несомненно, что эта последняя является производной от еще более широких эволюционных закономерностей.
К сожалению специальных исследований, посвященных атавизации гибридов и теоретическому анализу ее, мы имеем очень мало. Помимо Гайера (Guyer, 1909) атавизацию исследовал Битинский-Зальц (1933) у гибридов-бражников, где например нередко возникает на переднем крыле особая линия, считаемая атавистической. Наследование ее прослеживается и в дальнейших поколениях, однако по. неясной схеме. Для объяснения этого наследования автор предлагает ряд соображений в духе Гольдшмидта, гипотетически допуская различные изменения порога раздражения для различных скрещиваний, в результате чего, во-первых, рецессивные гены усиливают свое действие до степени доминантных, в связи с чем, во-вторых, может ослабиться разница между доминантными и рецессивными аллеломорфами и т. и. Подробнее на этих теоретических построениях мы здесь однако останавливаться не будем, так как подкрепляющий их экспериментальный материал еще слишком слаб и к тому же вовсе не исчерпано самое естественное объяснение атавизации комбинаторикой генов по типу явлений атавизации в скрещиваниях пород домашних животных.
Чрезвычайно интересный пример атавизации гибридов мы находим в изученном Крыжановским (неопубликованно) скрещивании леща (Abramis brama) с карасем (Carassius carassius) — представителей подсемейств Abramidinae и Cyprininae. Кровеносная система в области жаберных дуг у гибридов оказывается резко отличной от обоих видов и несвойственной и другим костистым рыбам, в то же время она напоминает кровеносную систему других, более примитивных групп рыб — осетровых.
 Рис. 74. Атавизация кровеносной системы у гибридов леща с карасем.
Правее и ниже глаза видны два сосуда — мандибулярная и гноидная дуги, соединенные комиссурой. По Крыжановскому, как и три следующие пояснительные схемы).
Рис. 74. Атавизация кровеносной системы у гибридов леща с карасем.
Правее и ниже глаза видны два сосуда — мандибулярная и гноидная дуги, соединенные комиссурой. По Крыжановскому, как и три следующие пояснительные схемы).
Рисунок 74 и схемы (рис. 75, 76, 77) поясняют этот поучительный случай. Более примитивный тип развития висцеральных кровеносных сосудов наблюдается у осетровых (Aciponser) (рис. 77). Сперва у них закладывается и функционирует лишь одна мандибулярная дуга аорты:
Md(d) +
Md(v). Затем появляются дорзальная половина гноидной дуги аорты
Hy(d) и комиссура
(сот), соединяющая ее с мандибулярной дугой аорты. После этого развивается вентральная половина гноидной дуги аорты
Hy(v). У костистых рыб сходная картина развития наблюдается лишь как
редкая индивидуальная вариация, отличаясь от типичной для Acipenser картины тем, что закладывающаяся дорзальная половина гноидной дуги аорты всегда более или менее рудиментарна (рис. 76). Обычно же у костистых рыб после мандибулярной дуги аорты появляются, только вентральная половина гноидной дуги аорты
Ну(с) и комиссура
(сот), соединяющая ее с мандибулярной дугой; дорзальная же половина гноидной дуги аорты совсем не развивается (рис. 75). Несомненно, что отсутствие
Hy(d) у костистых рыб есть явление новое. У гибридов Abramis brama × Carassius carassius
всегда появляется
Hy(d), нормально функционирующая вместе с вентральной половиной
Hy(v), т. е. у них наблюдается более примитивное состояние, приближающее их к ганоидам (Крыжановский С. Г. ex letr.).
 Рис. 75. Схема нормального развития сосудов костистых рыб.
Md (d) — дорзальная часть мандибулярной дуги; Md(u) — вентральная часть; com — комиссура, соединяющая мандибулярную дугу с гноидной, от которой осталась лишь вентральная часть [Hy(v)].
Рис. 75. Схема нормального развития сосудов костистых рыб.
Md (d) — дорзальная часть мандибулярной дуги; Md(u) — вентральная часть; com — комиссура, соединяющая мандибулярную дугу с гноидной, от которой осталась лишь вентральная часть [Hy(v)].
 Рис. 76. Схема сосудов, встречающаяся у костистых рыб в качество редкой индивидуальной вариации; сохранилась слабо развитая дорзальная часть гноидной дуги [Hy(d)].
Рис. 76. Схема сосудов, встречающаяся у костистых рыб в качество редкой индивидуальной вариации; сохранилась слабо развитая дорзальная часть гноидной дуги [Hy(d)].
 Рис. 77. Схема нормального развития сосудов у ганоидов (Acipenser): дорзальная часть гноидной дуги [Hy(d)] развита лучше вентральной [Hy(v)].
Рис. 77. Схема нормального развития сосудов у ганоидов (Acipenser): дорзальная часть гноидной дуги [Hy(d)] развита лучше вентральной [Hy(v)].
Этот случай является особенно интересным потому, что в свете генетического толкования атавизации он как бы указывает на большую древность расхождения названных подсемейств в ту эпоху, когда указанное примитивное строение должно было быть еще свойственно тем общим предкам, от которых развились и лещ и карась.
Дальнейшие исследования в этом направлении необходимы для окончательного разъяснения явления атавизации гибридов.
Очень интересен в этом же отношении изученный Ноаком (Noack, — 1908) череп гибрида льва и тигрицы, полученного Гагенбеком (Hagenbeck u. Aacken, 1897). Явление атавизации оказалось выраженным очень ясно в сильной редукции хищных зубов (гибрид оказывается менее хищником, чем оба родительские вида) и особенно в редукции мозговой капсулы. «Если бы, — замечает автор, — череп гибрида был найден в ископаемом состоянии…, то он был бы вероятно описан как новый вид и как более низко стоящая прародительская форма, из которой развились как львы, так и тигры и которая благодаря сходству с черепом ягуара являлась связующим звеном между крупными кошками Старого и Нового света» (Noack, 1908, s. 685).
Дальнейшую деталь вносят гибриды бражников Deilephila euphorbiae ♂ × ♀ D. hippophaes. Окраска нижней поверхности крыльев у этих гибридов оказывается несвойственной обоим родительским видам, но зато очень напоминает окраску далеко живущих островных видов D. dahlii (Корсика и Сардиния) и D. tithymali (Канарские о-ва). Мори (Могу, 1901, s. 352) толкует этот случай как атавизацию, считая, что на островах сохранились более древние формы, континентальные же euphorbiae и hippophaes ушли в своей эволюции дальше. Именно в этом смысле следует оценивать и собранный Штандфуссом многочисленный ряд примеров доминирования у гибридов признаков «филогенетически более древних видов» (ср. Standfuss, 1898, Ex. Zool. st., S. 43 и др.).
Новитация
Противоположностью атавизацин гибридов является возникновение новых признаков, не свойственных ни одному из родителей, так сказать,
новитация. Однако явлений этой категории мы знаем очень мало. У гибридов сатурний S. pavonia × pyri, в общем похожих на pavonia, но более крупных, поверхность крыльев между жилками оказывается волнистой, что сопровождается еще возникновением большего или меньшего количества разветвленных жилок, отсутствующих у чистых видов (Штандфусс). Эти жилки (рис. 78) не удается истолковать как атавизацию. Однако, чтобы отнести новообразование и категорию новитаций, надо доказать все же, что оно не является атавизацией. Возникновение гладкой головы у гибридов цесарка × петух является новостью по сравнению с обоими видами, но несомненным примером атавизации. Гибриды бражников Smerinthus ocellatus × S. populi вылупляются из куколок через три недели, не перезимовывая, в то время как оба родительские вида зимуют в стадии куколки (Kirk, 1890). Еще эфектнее гибрид психей Fumea nitidella × F. affinis. Они вылупляются без зимовки, в то время как F. nitidella как правило перезимовывает один раз, a F. affinis даже два раза (Pungeler, по Штандфуссу, 1896). Надо детально изучить всю эволюцию этих видов и при том с третичной эпохи, чтобы доказать, что это развитие без зимовки не является атавизацией, восстановлением признаков предков, живших в более теплом климате.
 Рис. 78. Добавочные жилки (а и b) у гибрида сатурний (по Штандфуссу).
Рис. 78. Добавочные жилки (а и b) у гибрида сатурний (по Штандфуссу).
У гибридов птиц нередко возникает красно-фиолетовый блеск вместо синего или зеленого у родителей [например тетерев-межняк (см. Огнев, 1929) и райские птицы (см. Штреземан, 1931, S. 15)] или вместо матовой черной окраски (Штреземан). Так как передвигание блеска в красную сторону связано с утолщением блестящих пластинок (Elsasser, 1925), т. е. с некоторых! огрубением структуры пера, то и это явление может оказаться атавизацией, а не новитацией.
Подозрение об атавизации отпадает лишь в случае новообразований патологического характера вроде случая с меланозисом у гибридов Xiphophorus. Это новообразование, как мы видели, объясняется встречей генов, никогда ранее не встречавшихся (и очевидно возникших после дивергенции обоих видов).
К новитации можно отнести и широко распространенное у гибридов явление гетерозиса.
Гетерозис
Под именем гетерозиса разумеется общее повышение жизнеспособности гибридов по сравнению с обоими родительскими формами, выражающееся наиболее наглядно в усилении роста, увеличении общих размеров и далее в понижении заболеваемости и процента отхода (смертности) на всех стадиях развития, в усилении плодовитости и различных сторон продуктивности. В англо-американской литературе говорят о «гибридной мощи» (hybrid vigor), в немецкой образован специальный глагол «роскошествовать» (luxuriiren), — говорят о «роскошествующих гибридах» (luxuriirenden Bastarden). Гетерозис не является специфическим свойством межвидовых гибридов. Столь же часто наблюдается он и при субгибридизации (скрещивании пород и сортов) и даже при скрещивании вообще неродственных популяций II линий, на чем в зоотехнии основано излишне популярное мероприятие «освежения крови» и имеющее крупное значение «промышленное скрещивание». Однако, как доказывает более тщательное изучение вопроса, гетерозис возникает не при всяком скрещивании, а степень его может быть весьма различной — от чуть заметной до исключительной, когда гибриды обнаруживают рост например в два раза больший, чем у исходных родителей.
 Рис. 79. Гетерозис у верблюдов (ориг. по фото Лакозы).
1— бактриан, 2 — дромедар, 3 — F1, 4 и 5 — два обратных скрещивания.
Рис. 79. Гетерозис у верблюдов (ориг. по фото Лакозы).
1— бактриан, 2 — дромедар, 3 — F1, 4 и 5 — два обратных скрещивания.
Для межвидовых скрещиваний возникновение гетерозиса является весьма характерным. Гибриды льва с тигром оказываются крупнее чистых видов так же, как гибриды одно- и двугорбого верблюдов (рис. 79). Гибриды яка с алтайским домашним скотом оказываются почти на центнер тяжелее, чем каждый из родителей, как показывают следующие цифры (Власов, Гершензон, Поляков, 1932) и кривая рис. 80.
 Рис. 80. Кривая роста гибридов як × тур (по Власову, Гершензону и Полякову).
Рис. 80. Кривая роста гибридов як × тур (по Власову, Гершензону и Полякову).
Вес (в килограммах) — 2 года, 6 лет
Яки — 153, 231
Турицы — 148, 208
Гибриды — 220, 335
Гиц (1933) исследовал гибридов яка со швицкой породой, более крупной, чем яки. С точки зрения промежуточности наследования таких количественных признаков, как рост, можно было бы ожидать у гибридов промежуточных размеров между яками и швицами. Фактически гибриды в годовалом возрасте оказались по величине равными взрослым ячкам и ничуть не уступающими годовалым швицам.
Высота в холке / Обхват груди / Косая длина туловища (лентой)
Ячки взрослые — 111,3 / 165,3 / 129,8
Гибриды годовалые — 112,3 / 159,3 / 129,5
Швицы годовалые — 113 / 141,3 / 133,3
Если по отношению к гибридам, полученным в зоопарках и и хозяйстве, можно иногда заподозреть лучший За ними уход, то эта возможность исключается по отношению к диким гибридам. Леннберг (Lönnberg, 1905) изучил зайцев-тумаков, гибридов беляки о русаком. Они оказались физически очень сильно развитыми. Изображенный на рис. 81 премолярный зуб гибрида сравнительна с родительскими видами хорошо иллюстрирует гетерозисное развитие жевательного аппарата гибрида.
 Рис. 81. Гетерозис у зайца-тумака.
Зуб тумака (посредине) и родительских видов (по Леннбергу).
Рис. 81. Гетерозис у зайца-тумака.
Зуб тумака (посредине) и родительских видов (по Леннбергу).
Подобный же гетерозис наблюдался у гибридов цесарки с петухами, которые по размерам почти не отличаются от петухов несмотря на значительно более мелкие размеры матерей — цесарок (Гайер, 1909), и у гибридов кур с фазанами. Гибриды домашних уток с мускусными по данным Соколовской крупнее обоих родителей или по крайней мере равны более крупным мускусным. Вылупляемость их из яиц оказалась также выше, чем у чистых уток.
У рыб резко выраженный гетерозис наблюдал Гершлер у гибридов Xiphophorus × Platypoecilus (рис. 82), Каммерер — у гибридов окуня с ершом, рост которых развивался быстрее, чем у обоих родителей. То же самое наблюдал Фишер у гибридов прудового карпа с карасем. Как упоминалось в главе II, гибриды обнаружили повышенную морозоустойчивость. Возможно, что на счет гетерозиса можно отнести частично и более быстрое развитие гибридов бабочек Fumea и Smerinthus, о котором мы упоминали в предыдущем параграфе, и повышение количества шелка вдвое гибридами Platysamia (Ватсон, 1893).
 Рис. 82. Гетерозис у гибридов рыб Xiphophorus × Platypoecilus.
Вверху слева — ♀ и ♂ Xiphophorus; справа — ♀ и ♂ Platypoecilus; внизу ♀ × ♂ F1 (по Гершлеру из Баура).
Рис. 82. Гетерозис у гибридов рыб Xiphophorus × Platypoecilus.
Вверху слева — ♀ и ♂ Xiphophorus; справа — ♀ и ♂ Platypoecilus; внизу ♀ × ♂ F1 (по Гершлеру из Баура).
Гетерозис может обнаруживаться очень рано, еще у гибридных эмбрионов, понижая процент гибели на ранних стадиях, повышая процент вылупления. Как увидим более подробно ниже, Funrhilus heteroclitus развивается быстрее, чем F. majalis. Гибриды heteroclitus ♂ × ♀ majalis развиваются быстрее, чем majalis, с первых же часов эмбриогенеза. Но что дело здесь идет не просто о доминировании быстрого над медленным, видно и из того, что при оплодотворении «быстрых» яиц heteroclitus «медленной» спермой majalis тоже получается некоторое убыстрение дробления.
Физиология гетерозиса изучена совершенно недостаточно. Вермель (неопубл.) констатировал у гибридов як × тур более крупные размеры клеток, чем у обоих родительских видов. У таких же гибридов Самохвалова и Кожарин нашли повышенное количество гемоглобина. Некоторые зоологи ставят вопрос о том, не стоит ли гетерозис в связи с нарушением половой деятельности гибридов, столь свойственным гибридам. Наличие гетерозиса у субгибридов, обнаруживающих нередко даже повышенную плодовитость, этому взгляду на природу гетерозиса противоречит, однако здесь необходимо более тщательное изучение эндокринологии гибридов.
Исчерпывающей теории гетерозиса мы еще не имеем.
Сторонники биохимической теории гибридизации (см. главу VII) пытались свести гетерозис к слабым степеням взаимного «отравления» соединенных в гибриде хромосом (и плазмы различного происхождения). Критику этой точки зрения мы уже дали. Она совершенно не учитывает несомненных менделестических элементов, обусловливающих гетерозис. Хотя и возможно, что менделистическое толкование не исчерпывает всего явления, но это нужно еще доказать.
Согласно этой теории гетерозис возникает благодаря тому, что гибрид оказывается богаче доминантными генами, чем каждый из родителей. Пусть мы скрещиваем два вида, имеющие одинаковый рост, например яка с ойротским скотом. Нет сомнения в том, что те гены, которыми в каждом виде обусловлен этот рост, не тождественны, по крайней мере в некоторой части, например:
вид
А, гены роста АА, ВВ, ЕЕ, FF, GG, KK, LL
вид
В, гены роста АА, СС, DD, ЕЕ, II, ММ, NN
Гибриды А × В будут иметь генотип:
АА Bb, Сс, Dd, ЕЕ, Ff, Gg, Ii, Kk, LI, Mm, Nn.
В то время как у родителей было по 7 генов, усиливающих рост, у гибрида их 11. Правда, большинство их находится в гетерозиготном количестве. Но если гены BCDF… более или менее полно доминируют, то их эфект несмотря на гетерозиготность будет значительно выше, чем эфект у гомозиготных генов.
Эта гипотеза объясняет, почему при скрещивании различных видов гетерозис возникает в различной степени, почему он связан особенно с F
1 и т. д.
Более подробно в обсуждение вопроса о гетерозисе, трактуемого во всех современных курсах генетики и разведения, мы здесь входить не будем и остановимся лишь на следующих, важных для гибридизации соображениях.
Если явления атавизации гибридов (в некоторых группах) мы связали с эволюционной закономерностью — с преимущественным участием рецессивных признаков в диференциации видов, то и явление гетерозиса нам кажется возможным связать с другой закономерностью эволюции —
эволюционным параллелизмом. В самом деле, если мы имеем два родственных вида
А и
В, то с большой долей вероятности можно ожидать, что то, что благоприятно для одного вида, будет более или м нее благоприятно и для другого, в меру степени их сходства. Если для процветания вида
А в борьбе за существование полезно увеличение роста, то более или менее в такой же степени увеличение роста будет полезно и для вида
В. Поэтому, если естественный отбор будет накапливать в виде
А гены, усиливающие рост, то при общем сходстве, мутационного процесса и условий борьбы за существование и в генофонде вида
В будет итти подобное же накопление генов роста. То же самое можно будет сказать по отношению к любому, другому признаку обоих видов, так как даже при некотором различии видов
А и
В сходство их всегда в тысячи раз сильнее различия, а следовательно и движущие силы их эволюции имеют значительно больше сходных черт, чем различий. Таким образом и эволюция даже уже разошедшихся родственных видов
А и
В наряду со специфическими особенностями для каждого отдельного вида (в меру их различия) будет иметь и общие черты — в меру их сходства друг с другом. Отсюда совершенно неизбежны и черты параллелизма в эволюции родственных видов, дающие соблазнительную пищу для эволюционистов-идеалистов, видящих в явлениях параллелизма проявление не сходной диалектики развития сходных видов, но различных мистических сил.
Наличие параллелизма эволюций должно приводить к сходным изменениям генотипов видов
А и
В. Но столь же естественно, что это сходство не будет полным, и если например увеличение роста в виде
А будет достигнуто накоплением генов Х
1, Х
2, Х
3 … то в виде
В могут накопиться частично те же самые гены, а частично другие, например Х
1, Х
2, Х
4 … Так как несомненно, что хотя ныне живущие организмы и являются-достаточно приспособленными к жизни, но ни в коем случае не предельно приспособленными, то надо ожидать, что в большинстве случаев гибрид, возникший от скрещивания
Х
1Х
2Х
3 × Х
1Х
2Х
4
|
Х
1Х
2Х
3Х
4
и соединивший все накопленные обоими видами гены, окажется еще более жизнеспособным, чем каждый из, родителей. Здесь произойдет как бы «обмен эволюционным опытом») обоих видов, суммирование их эволюционных достижений —
ададаптация.
Поскольку каждый из видов имеет свою специфику и в процессе эволюции приобрел свою специальную адаптацию, не отвечающую потребностям другого вида, гибридизация неизбежно должна сопровождаться и дезадаптацией, многочисленные примеры чего мы видели в, главе о нежизнеспособности гибридов и их бесплодии. Таким образом общая жизнеспособность гибрида должна явиться результатом одновременной ададаптации и дезадаптации. В зависимости от удельного веса, соотношения обоих этих элементов может получиться или итоговое повышение жизнеспособности или понижение ее.
При очень близком родстве форм оба элемента, а особенно дезадаптация, будут слабы, и мы будем иметь слабый гетерозис. При более сильном удалении оба элемента возрастут, но поскольку сходство скрещиваемых видов будет значительно больше их различия, элемент ададаптации будет значительно преобладать, что приведет к усилению гетерозиса. Однако далее начнут усиливаться элементы дезадаптации, на что будет реагировать в первую очередь locus minoris resistentiae, пока гибриды не будут получаться вовсе нежизнеспособными.
Толкование гетерозиса как «объединения эволюционных достижений)» обоих скрещенных видов объясняет одновременно и то, почему в F
2 и т. д. гетерозис как правило исчезает. Это исчезновение идет по двум линиям. Во-первых, только в F
1 имеются одновременно оба набора хромосом скрещенных видов в полном комплексе с их «эволюционными достижениями», в F
2 таких животных остается лишь 1:2
n — доля, где n равно числу хромосом, если хромосомы обоих видов не дают перекреста, или равно числу генов, различающих оба вида. Так как это число не ниже 20, то доля особей F
2, повторяющих свойства F
1 оказывается порядка 1 миллионной, т. е. практически равна нулю. С другой стороны, распре-деление генов между хромосомами. у разных видов тоже различно, и адаптивный характер носит лишь весь набор хромосом видов в целом. При свободном перекомбинировании хромосом, наступающем в F
2, лишь миллионные доли сохраняют адаптированную комбинацию хромосом, остальные же получают комбинацию хромосом с нарушением адаптации, либо вовсе летальную, почему F
2,
оцененное по средним показателям, естественно окажется более или менее значительно ниже, чем F
t и даже чем родительские виды.
Развитие признаков гибридов
Начало влияния сперматозоида
Вопрос о том, когда вступает в действие сперматозоид в качестве передатчика наследственных задатков отца гибриду, или иначе, когда вступает в действие генотип, привнесенный в гибрида через сперматозоид, и когда он начинает оказывать влияние на ход эмбриогенеза гибрида — принадлежит к числу чрезвычайно трудных и до сего времени еще спорных вопросов.
Изложенные в главе VI экспериментальные исследования по гетерогенному оплодотворению главным образом у иглокожих склонили большинство исследователей к взгляду, что на ранних стадиях эмбрионального развития роль генотипа, привносимого сперматозоидом, отступает на задний план, и это раннее развитие проходит вполне по материнскому типу. Таковы например были многочисленные случаи возникновения в результате гибридизации плютеусов чисто материнского типа. Более тщательное изучение показало однако: 1) что в ряде случаев происходит элиминация отцовских хромосом, в связи с чем конечно уже нельзя вообще говорить о сроке вступления их в действие, 2) в ряде случаев плютеусы получаются несомненно промежуточного типа (например Strong. purpuratus × S. franciscanus), где уже несомненно, что по крайней мере на стадии плютеуса хромосомы обоих родителей оказываются по силе своего действия равноправными. Тем не менее такие авторитеты, как Бовери, Конклин и др., высказывались за то, что развитие эмбриона, по крайней мере до стадии бластулы и гаструлы, определяется всецело структурой яйца. Так, Конклин писал: «факт остается фактом, что в момент оплодотворения наследственная потенция обеих половых клеток не одинакова; все раннее развитие, включающее возникновение полярности, симметрии, типа дробления и относительное расположение и пропорцию будущих органов, оказывается предетерминированным в цитоплазме яйца, тт только диференциация позднейшего развития оказывается под воздействием спермы. Вкратце, яйца устанавливают тип развития, и ядру сперматозоида принадлежат лишь детали».
 Рис. 83. Fundulus heteroclitus, majalis и diaphanus и Cyprinodon variegatus, самки, сверху вниз. (По Ньюману.)
Рис. 83. Fundulus heteroclitus, majalis и diaphanus и Cyprinodon variegatus, самки, сверху вниз. (По Ньюману.)
Против этой точки зрения выступил с данными своих экспериментов по гибридизации рыб Ньюман (1908, 1910, 1914); проделавший, правда, на протяжении своих исследований существенную эволюцию взглядов. Для опытов он воспользовался наличием ясной разницы в темпе эмбриогенеза у трех видов рыб рода Fundulus (рис. 83), из которых F. heteroclitus развивается наиболее быстро, F. diaphanus — средне, a F. majalis — наиболее медленно. В первой работе Ньюман обнаружил ясное влияние сперматозоида на развитие яйца через 14 часов по оплодотворении и предположил, что при более точной методике удастся установить еще более раннее влияние. В работе 1910 г. Ньюману удалось показать, что уже через 4 часа обнаруживается различие в темпе дробления яиц «чистых» и яиц гибридных, причем, что особенно важно, удалось показать, что темп дробления, яиц F. majalis ускоряется под влиянием оплодотворения их спермой F. heteroclitus. Дробление F. heteroclitus идет примерно на
1/
4—
1/3 быстрее, чем у F. majalis, а все развитие до вылупления мальков F. heteroclitus проходит в 2 недели, у F. majalis — в 3 недели. Второе деление дробления F. majalis наступает через 4 часа по оплодотворении, у F. heteroclitus — через 3 часа, что вероятно стоит в связи и с размерами рыб и их икринок.
 Рис. 84. Шкала дробления яиц Fundulus. (По Ньюману.)
Рис. 84. Шкала дробления яиц Fundulus. (По Ньюману.)
Приведем для иллюстрации 6-й опыт. Предварительно за несколькими яйцами было проведено индивидуальное наблюдение, и примерно через равные промежутки времени были сделаны зарисовки стадий (рис. 84). Затем икра крупной самки была разделена на две части, одна осеменена спермой того же вида F. majalis, а другая часть гибридизирована спермой F. heteroclitus. Приблизительно через 4 часа дробившиеся яйца были зафиксированы и расклассифицированы по стадиям (если стадия попадала между нарисованными, то классифицировалась дробно, например 3/
2), Таблица показывает результаты:

Среднее для чистых — 5,11, для гибридов —5,57, разница почти на
1/
2 стадии.
Более отчетливо разница выступила при таком сопоставлении:
Стадия Чистые Гибриды
1—5
1/
2 64,3 % 38,3%
6—7 35,7 % 60,7%
В 6 подобных опытах всюду оказалось ускорение у гибридов, хотя в одном опыте и очень незначительное. В первом эксперименте яйца были фиксированы уже черев 3 часа 45 мин. после оплодотворения и дали даже наибольшее превышение гибридов над чистыми. Отсюда можно было следовательно заключить, что ускоряющее влияние сперматозоидов «быстрого» вида на дробление F. majalis начинается чрезвычайно рано, очевидно уже при первом делении. Применив цитологический метод, можно было бы выяснить разницу в подготовке первого веретена. Вместе с тем Ньюману пришлось отметить влияние гетерогенного оплодотворения и на характер дробления, так как в гибридных сериях значительно чаще встречалась несимметричная стадия «трех бластомеров», т. е. стадия, на которой один из двух первых бластомеров уже успел разделиться на два дочерних, обогнав второй.
 Рис. 85. Развитие гибридов Fundulus.
d — diapliaiuis; md — majalis ♂ × ♀ diaphanus; hd — heteroclitus ♂ × ♀ diaphanus.
Стадии сверху вниз — 20 час., 2 дня, 4 дня, 7 дней. (По Ньюману.)
Рис. 85. Развитие гибридов Fundulus.
d — diapliaiuis; md — majalis ♂ × ♀ diaphanus; hd — heteroclitus ♂ × ♀ diaphanus.
Стадии сверху вниз — 20 час., 2 дня, 4 дня, 7 дней. (По Ньюману.)
В результате этих экспериментов Ньюман решительно стал на точку зрения Фишеля, что с самого начала роль сперматозоида является формативной, почему он может наложить печать своего вида уже на ранние стадии развития организма, в то время как цитоплазма яйца представляет собой, лишь материал для формирующей роли комбинированного ядра обоих родителей.
В работе 1914 г. Ньюман возвращается к тому же вопросу с новым материалом, произведя сравнительное изучение эмбриогенеза в скрещиваниях с третьим, видом F. diaphanus. Этот вид по скорости развития занимает промежуточное место между heteroclitus и majalis. Произведя оплодотворение икры diaphanus спермой всех трех видов, Ньюман обнаружил, что- гибридизация с heteroclitus ясно ускорила развитие, а гибридизация с majalis замедлила, его по сравнению с чистыми diaphanus.
Рисунок 85 показывает степень этого влияния.
В этих данных действительно бросается в глаза факт специфического влияния сперматозоида в сторону придании развитию гибрида свойств его отцовского вида. Тем не менее здесь возможны следующие сомнения: 1) не является ли замедление развития гибридов diaphanus × majalis простым результатом их отмеченной малой жизнеспособности и 2) соответственно не является ли ускорение в сочетании diaph. × heteroclitus результатом гетерозиса, а не тем, что гибрид унаследовал свойства F. heteroclitus.
На первый вопрос легко ответить отрицательно, так как яйца F. majalis, оплодотворенные спермой F. diaphanus, развиваются быстрее чистых majalis, хотя эта гибридная комбинация также нежизнеспособна.

Здесь отчетливо видно, что несмотря на слабость и нежизнеспособность гибридной комбинации гибриды с первых же. часов развеваются заметно быстрее, очевидным образом под влиянием свойств быстро развивающегося F. diaphanus, перенесенных сперматозоидом.
Что же касается ускоряющего действия F. heteroclitus, то здесь картина несколько сложнее, так как в этой комбинации несомненно развитие гетерозиса, что. выражено в том, что в обоих реципрокных скрещиваниях F. heteroclitus × F. diaphanus развитие гибридов идет быстрее. Впрочем это ускорение очень незначительно и заметно не во все моменты эмбрионального развития, так что можно вполне допустить, что развитие гибрида diaphanus × heteroclitus идет почти как у heteroclitus, потому что два противоположных влияния — замедляющее влияние генотипа diaphanus и гетерозисное ускорение — взаимно нейтрализуются. Это однако все же подтверждает раннее начало действия отцовских хромосом.
Наследование эмбриональных признаков у гибридов
Наследование эмбриональных признаков у гибридов изучено у морских ежей. Несмотря на то, что получение гибридных личинок у морских ежей началось с восьмидесятых годов XIX в. и проводилось рядом выдающихся исследователей, правильное истолкование. наследственных явлений при этом дано было впервые лишь в 1910 г. Лебом, Кинг и Муром, после того как получили достаточное распространение идеи менделизма. До этого же времени споры велись вокруг того, отцовские или материнские признаки перевешивают у гибридов, оказывает ли сперматозоид влияние на эмбриогенез гибридов или нет и пр.
 Рис. 86. Плютеусы Strongylocentrotus purpuratus (2 и 4) и F1 purpur. ♀ × ♂ franciscanus (1 и 3); 6-дневные (1 и 2) и 14-дневные (3 и 4). (По Лебу, Кинг и Мур.)
Рис. 86. Плютеусы Strongylocentrotus purpuratus (2 и 4) и F1 purpur. ♀ × ♂ franciscanus (1 и 3); 6-дневные (1 и 2) и 14-дневные (3 и 4). (По Лебу, Кинг и Мур.)
В 1909/10 г. почти одновременно названные авторы, с одной стороны, и Теннент (Tennent, 1909) — с другой, четко поставили вопрос о доминировании отдельных признаков у гибридов. Леб, Кинг и Мур поставили реципрокные скрещивания двух тихоокеанских ежей Strongylocentrotus franciscanus и St. purpuratus, получив большие количества как гибридов, так и контрольных чистых форм от тех же самок.
Взрослых морских ежей в лабораториях выращивать не удается, и опыт касается лишь развития до плютеуса включительно. Сравнивая чистые виды, авторы устанавливают такие различия между ними (рис. 86 и 88).

Гибридные плютеусы franciscanus ♂ × ♀ purpuratus, показанные на рис. 86, фиг. 1 (6-дневные) и на рис. 86, фиг. 3 (14-дневные), воспитывались в одинаковой температуре и одинакового возраста с изображенными рядом чистыми формами. На рис. 87 показан отдельно, по точным измерениям, скелет спереди и сбоку. Как видно, форма тела гибрида округла, как у franciscanus, а не пирамидальна, и руки развиты тоже, как у franciscanus, так что и форма тела и длина рук franciscanus обнаруживают доминирование. Наоборот, ветвь
a скелета имеет форму палкообразную, как у purpuratus, но однако не гладкую, а с ясными шипами и сучками, давая таким образом комбинацию двух доминирующих признаков, идущих от разных родителей. Далее средняя ветвь
f хорошо развита, как у franciscanus, как и ветвь
d. Две другие ветви с и
d развиты почти как у franciscanus, хотя и с ясным ослаблением.
 Рис. 87. Скелет Strongylocentrotus purpuratus (1), St. franciscanus (3) и F1 pur. ♀ × ♂ fr. (2). Спереди и сбоку (1a, 2a и За.)
Рис. 87. Скелет Strongylocentrotus purpuratus (1), St. franciscanus (3) и F1 pur. ♀ × ♂ fr. (2). Спереди и сбоку (1a, 2a и За.)
Следовательно общий промежуточный характер скелета этих эмбрионов Довольно легко разлагается в «мозаику» доминантных отдельных признаков обоих родителей, большинство которых здесь оказывается, правда, отцовскими.
Чтобы доказать, что мы имеем здесь дело не с «отцовскими» признаками, а именно с «доминантными», свойственными виду franciscanus, надо было исследовать реципрокных гибридов. Здесь встретилось осложнение, так как эти. гибриды (purpuratus ♂ × ♀ franciscanus) оказались в противоположность реципрокным очень мало жизнеспособными. Дробление их идет со скоростью чистых franciscanus, но со стадии гаструлы начинается отставание (рис. 88). Несмотря на эти затруднения и на гибель большинства личинок purpuratus ♂ × ♀ franciscanus авторы получили все же тысячи таких реципрокных гибридов. Они оказались неотличимыми от реципроков, в частности по булавовидной верхней ветви скелета. Часто встречавшиеся более слабо развитые руки относятся конечно за счет общего замедления развития и болезненности.
 Рис. 88. Плютеусы Strongylocentrotus franciscanus (1 и 2) u F1 fr. ♀ × ♂ pur. (3 и 4) в возрасте 4 дней. (По Лебу, Кинг п Мур.)
Рис. 88. Плютеусы Strongylocentrotus franciscanus (1 и 2) u F1 fr. ♀ × ♂ pur. (3 и 4) в возрасте 4 дней. (По Лебу, Кинг п Мур.)
Таким образом гибриды
f ×
р и
р ×
f оказываются одинаковыми. Они нимало не обнаруживают какого-либо превалирования «отцовских» или «материнских» признаков, но проявляют вполне определенные отдельные доминирующие признаки обоих видов: от purpuratus они получают палкообразную форму ветви
а скелета, от franciscanus — округлую форму тела, быстро растущие руки, сильную сучковатость скелета и большее- развитие его ветвей
с, b, d, f.
Следует отметить в качестве материнского признака окраску плютеусов, но так как- пигмент в плютеусе есть просто пигмент яйца, то это не имеет значения с точки зрения наследственности.
Межродовые гибриды морских ежей. Разобранный только что случай представляет собой сравнительно близкую гибридизацию, так как оба вида являлись представителями одного и того же рода. Полученная здесь картина и ее анализ весьма близки к тому, что мы видели и при скрещивании двух близких видов молюсков в опытах Ланга и в других разобранных выше примерах взрослых гибридов, а именно, что при сравнении гибрида с родительскими формами без особого труда удается установить доминирование ряда отдельных родительских признаков.
Чем систематически дальше будут отстоять друг от друга скрещенные виды, тем большим числом генов они будут различаться, тем труднее при столь же ясно выраженных различиях между их признаками будет подвергать их анализу и выделять из общей комбинации отдельные доминантные признаки. Хорошим примером этого могут служить другие более отдаленные гибриды морских ежей. На рис. 89 представлены результаты такой межродовой гибридизации Echinus × Sphaerechinus. Степень морфологического различия их плютеусов примерно та же, как и в скрещивании St. purpuratus × St. franciscanus, тем не менее выделить отдельные доминантные элементы здесь уже гораздо труднее. Так, можно легко отметить доминантный характер длинных рук Echinus, особенно по длине соответствующих скелетных частей, рецессивный характер сложной сети перекладин скелета туловища Sphaerechinus и доминантный характер более мелких размеров Sphaerechinus, но в большинстве других признаков приходится говорить уже о Промежуточном характере признаков гибрида — форма туловища и направление — рук, строение скелета длинных рук, степени булавовидности скелетных балок, глубины вырезки между руками и т. д.
 Рис. 89. Плютеусы Echinus (3 и За) и Sphaerechinus (2 и 2а) и F, (1 и 1а.)
Рис. 89. Плютеусы Echinus (3 и За) и Sphaerechinus (2 и 2а) и F, (1 и 1а.)
Подводя итоги этим опытам с рыбами и морскими ежами, надо притти к выводу, что закономерности образования фенотипа у этих
ранних эмбрионов те же самые, что и у взрослых гибридов, и что Вместе с тем гибридность фенотипа оформляется уже на самых ранних стадиях эмбрионального развития, как только складываются те признаки, за которыми
технически удается установить наблюдение.
Форма клеток гибридов
Форма клеток гибридов подверглась тщательному изучению Бэнкрофтом (Bancroft, 1912) и Ньюманом (1914) у гибридов Fundulus. У этих рыбок, начиная с первых дней эмбрионального развития, образуются черные и красные пигментные клетки, весьма специфические у каждого вида.
Если обратиться к красным желточным пигментным клеткам (см. цветную таблицу II), то их видовые особенности следующие: у F. diaphanus они красно-коричневого цвета, сильно и глубоко разветвлены (см. цветную таблицу, фиг. d), у F. heteroclitus они ярко-красного цвета, несколько крупнее, но не так сильно разветвлены (фиг. h), и наконец у F. majus они тоже яркокрасные и разветвлены на тонкие немногочисленные ветви (фиг. m).
У гибридов пигментные клетки имеют в большинстве случаев совершенно промежуточный характер. У реципрокных гибридов diaphanus × heteroclitus промежуточный характер наиболее ясно выражен в их желтоватой окраске (фиг. dh и hd). Такая же промежуточная окраска и у клеток гибридов diaphanus × majalis (фиг. dm и md), но форма здесь резко иная, чему гибридов, упомянутых первыми, — участие majalis сообщает гибридным клеткам соответствующий характер ветвления — ветвей гораздо меньше, они длинные, тонкие и заостренные. Наконец у реципрокных гибридов у heteroclitus × majalis (фиг. hm и mh) окраска, как у обоих родительских видов, ярко красная, а форма типично промежуточная.
Интересно отметить, что во всех трех парах между реципроками существуют различия, которые могут быть сформулированы так, что материнский вид оказывает несколько больше влияния, чем отцовский. Так, в парах heteroclitus × majalis и diaphanus × majalis в тех случаях, когда материнским Видом является majalis, заостренность ветвей выражена гораздо отчетливее (фиг. hm и dm). Более спорной является эта разница в паре heteroclitus × diaphanus, где сходство гибридов с родителями можно толковать по различному. Обсуждать это явление будет иметь смысл лишь тогда, когда будет отмечаться пол гибридов и следовательно судьба Х и У-хромосом.
Несколько более своеобразно ведут себя черные пигментные клетки головы. Они также резко различны у трех видов: звездчатые у diaphanus, многоугольные у heteroclitus и радиальные у majalis. Реципрокные скрещивания здесь дают очень различные результаты, изображенные на рис. 90.
В двух скрещиваниях, когда гибриды выходят из яиц majalis, получается полное доминирование
отцовского типа — у гибридов majalis ♀ × ♂ diaphanus, клетки звездчатые, почти как у diaphanus, и во всяком случае нисколько не приближаются к радиальному типу majalis (фиг. dm). Во втором скрещивании majalis ♀ × ♂ heteroclitus клетки гибридов также типичны для heteroclitus, лишь несколько меньше (фиг. mh).
Однако в реципрокном скрещивании diaphanus ♀ × ♂ majalis получаются уже типичные. промежуточные звездчато-радиальные клетки, т. е. majalis начинает действовать, став отцом. Наоборот, в скрещивании heteroclitus ♀ × ♂ majalis снова возникают. Типичные клетки heteroclitus, но к ним добавляется другой тип, «измененный majalis» (Ньюман), который однако лишь с трудом удается распознать как промежуточный heteroclitus-majalis, а вернее толковать их как новообразование (фиг. hm).
Также совершенно различны реципроки diaphanus × heteroclitus. В комбинации diaphanus ♂ × ♀ heteroclitus мы имеем замечательную промежуточную форму — клетки средние между звездчатыми и полигональными, и при этом обнаруживают свойство клеток heteroclitus сливаться друг с другом.
В реципрокном скрещивании heteroclitus ♂ × ♀ diaphanus мы имеем, как и в heteroclitus × majalis два типа клеток — типичные клетки heteroclitus
и наряду с ними довольно типичные звездчатые клетки diaphanus.
 Рис. 90. Черные пигментные клетки Fundulus и их гибридов.
h — heteroclitus, d — diaphanus, m — majalis; у гибридов первая буква указывает отца, вторая — мать, кроме hm u mh, где наоборот. (По Бэнкрофту.)
Рис. 90. Черные пигментные клетки Fundulus и их гибридов.
h — heteroclitus, d — diaphanus, m — majalis; у гибридов первая буква указывает отца, вторая — мать, кроме hm u mh, где наоборот. (По Бэнкрофту.)
Таблица II
 Красные пигментные клетки Fundulus и их гибридов.
h — heteroclitus, d — diaphanus, m — majalis. У гибридов первая буква указывает отца, вторая — мать.
Красные пигментные клетки Fundulus и их гибридов.
h — heteroclitus, d — diaphanus, m — majalis. У гибридов первая буква указывает отца, вторая — мать.
Эти данные действительно замечательны: насколько форма красных клеток была ясно понятна, настолько в судьбе черных клеток мы видим хаос совершенно пока непонятных явлений, не поддающихся никакому обобщению. Мы имеем 1) полное доминирование, 2) промежуточное доминирование признаков и 3) мозаику. Реципрокные скрещивания различаются также совершенно различным образом:
мозаика — промежуточность
доминирование — промежуточность
мозаика — доминирование
Резкое различие реципрокные скрещиваний говорит как бы за роль цитоплазмы яйца. Но тогда следовало бы ожидать матроклинии в типе клеток подобно наблюдающемуся у красных в слабой степени. Ио здесь скорее наоборот — оба скрещивания с majalis дают полное доминирование
отцовского типа тогда, когда гибриды выходят из яиц majalis, а некоторое влияние majalis обнаруживается тогда, когда он фигурирует в качестве отца. В скрещивании diaphanus × heteroclitus ни в одном направлении нет доминирования одного типа над другим, но при dh-направлении мы имеем промежуточный тип (с некоторым преобладанием отца), а в реципрокном — мозаику чистых родительских форм.
Можно думать, что форма клеток принадлежит к числу чрезвычайно глубоких «основных» признаков организма, приводящих к многообразным вторичным признакам. Тем интереснее подчеркнуть, что в данном случае при скрещивании сравнительно близких видов мы встречаемся со столь затруднительным для исследования явлением наследственности гибридов. К сожалению и здесь отсутствуют необходимые данные о поле изученных гибридов, и вопрос явно требует переисследования.
Форма хромосом гибридов
В этой главе также необходимо рассмотреть некоторые вопросы кариологии. Прежде всего следует выяснить, какой вид имеют хромосомы различных видов — если эти хромосомы различны — в гибридной клетке. Здесь возможно несколько априорных предположений. Если вид хромосом, их длина, толщина, перетяжки, место прикрепления нити веретена и пр. являются лишь элементами фенотипа гибрида, то здесь должны действовать те же закономерности, которые нами были выше рассмотрены, т. е. вид хромосом должен стать более или менее промежуточным с наклонностью к атавизму, с отдельными элементами отцовско-материнской мозаики и т. п. Возможно другое ожидание, сделанное в свое время Годлевским (1906), что отцовские хромосомы, попав в плазму, яйца, примут форму хромосом яйца, если считать, что именно плазма определяет форму хромосом. Наконец о ели форма хромосом свойственна самим хромосомам, то можно рассчитывать видеть в клетках гибридов рядом хромосомы обоих родительских видов в их специфической форме.
Одни из первых последовали этот вопрос Годлевский (1906) у ультрагибридов морских ежей с морскими лилиями и Менкгауз (1894) при гибридизации рыб Fundulus × Menidia. Годлевский однако не сумел еще различить хромосомы ежа от хромосомы лилии. Менкгауз в скрещивании Fundulus ♀ × ♂ Menidia наблюдал хромосомы обоих видов, оказавшиеся различными, и видел, как и последовательных делениях эти хромосомы перемешивались, но не утрачивали своего специфического вида.
К таким же выводам и к сожалению почти на том же материале пришла в 1914 г. М. Моррис (Fundulus heteroclitus ♀ × ♂ Ctenolabrus adspersus). Хромосомы этих видов совершенно различны по форме: круглые точкообразные у Ctenolabrus и палочкообразные у. Fundulus. У Fundulus они раза в 3 длиннее, — но несколько тоньше, чем у Ctenolabrus (рис. 91). В клетках гибридов эта разница сохраняется совершенно одинаковой вплоть до той стадии, до которой удалось проследить митозы (шестое деление дробления). К моменту этого деления хромосомы разной формы, располагавшиеся вначале группами, перемешались, но сохранили в точности свою форму.
 Рис. 91. Анафазы первого дробления Fundulus (слепа) и Fundulus ♀ × ♂ Ctenolabrus. (По Моррис.)
Рис. 91. Анафазы первого дробления Fundulus (слепа) и Fundulus ♀ × ♂ Ctenolabrus. (По Моррис.)
Несколько ранее, в 1910 г., Бальцер пересмотрел вопрос и относительно иглокожих, в частности относительно гибридов еж × лилия, изученных Годлевским. Факты, полученные — Бальцером, целиком совпадают с фактами Менкгауза и Моррис. Бальцеру удалось отметить прежде всего общие различия между хромосомами видов. Так, оказалось, что у Arbacia хромосомы короткие, а у Strongylocentrotus более длинные, и лишь немногие меньшие хромосомы Strongylocentrotus не отличимы по длине от больших хромосом Arbacia. В митозах гибридов эта разница сохраняется в полной мере (рис. 92).
 Рис. 92. Хромосомы Strongylocentrotus и Arbacia в митозе гибрида; два среза с одного митоза. (По Бальцеру.)
Рис. 92. Хромосомы Strongylocentrotus и Arbacia в митозе гибрида; два среза с одного митоза. (По Бальцеру.)
Мало того отдельные хромосомы Strongylocentrotus, именно крючкообразные, позволяют без труда узнать себя как в собственной, так и в гибридной пластинке.
 Рис. 93. Хромосомы Strongylocentrotus и Antedon в общем митозе гибрида.
Палочкообразные — ежа, v — образные лилии. (По Бальцеру.)
Рис. 93. Хромосомы Strongylocentrotus и Antedon в общем митозе гибрида.
Палочкообразные — ежа, v — образные лилии. (По Бальцеру.)
Иного характера отличие обнаружилось у хромосом лилии Antedon. Здесь оказались частыми дужкообразные, двуплечие хромосомы, не встречающиеся у ежей или встречающиеся лишь в виде исключения. На рис. 93, представляющем ультрагибридную пластинку еж × лилия, совершенно отчетливо видны и палочкообразные хромосомы Strongylocentrotus и его одна пара крючкообразных (на правом срезе митоза) и наконец несколько двуплечих хромосом лилии. Таким образом хромосомы морской лилии, попадая в столь для них чуждую цитоплазму, как плазма яйца ежа, сохраняют свою форму повидимому неизменной
[39].
В настоящее время таким образом можно считать доказанным, что как бы ни были далеки друг от друга скрещиваемые формы, но если только в гибридах удается рассмотреть митозы, то в них можно найти в неизмененном виде хромосомы родительских форм с их характерным числом, длиной, толщиной и формой. Возможно конечно, что. при тщательном изучении удастся найти некоторые изменения, но они во венком случае будут столь незначительны, что их нужно специально искать, и они нисколько не стушевывают имеющихся между хромосомами различий.
Для гибридизации отсюда возникает ряд очень глубоких вопросов. Прежде всего чрезвычайно важна проблема значения для организма и для его правильной жизнедеятельности того общего «стиля»-хромосом, который так бросается в глаза например при сравнении хромосом Fundulus и Ctenolabrus, Strongylocentrotus и, Arbacia и т. д. Имеет ли какое-либо
приспособительное значение то, что у Arbacia все хромосомы короткие, а у Strongylocentrotus длинные? Стоит ли это в связи с величиной клеток или с силой центросом и крепостью нитей веретена (см. ниже об элиминации хромосом)? Если да, то возможно ли правильное функционирование хромосом в клетках несоответствующей величины, — какой должна быть величина клеток для того, чтобы обеспечить правильное функционирование хромосом, и т. д.? Или может быть (что мало вероятно), эти свойства хромосом большого значения не. имеют и, проектируй и создавая новый организм, можно не заботиться о характере его хромосом, и т. д.? К сожалению в настоящее время мы почти ничего по этому поводу основательного не знаем. На следующих страницах увидим попытку Бальцера непосредственно связать вопрос о длине хромосом с явлением элиминации — попытку чрезвычайно интересную и серьезную.
Митозы гибридов. Элиминация хромосом
Переходя к вопросу о том, как происходят у гибридов митозы, в которых участвуют-хромосомы различного происхождения, цитоплазма одного вида и центросома, принесенная сперматозоидом другого вида, Приходится отметить, что у гибридов митозы могут итти и совершенно правильно и в высшей степени неправильно. При этом важно подчеркнуть, что правильность или неправильность митозов как будто ни в какой обязательной связи с отдаленностью скрещенных форм не стоит. Достаточно-таких примеров: митоз у гибрида двух морских ежей Strongylocentrotus × Sphaerechinus идет совершенно, неправильно, а митоз у ультрагибрида Strongylocentrotus × Antedon (морская. лилия) идет совершенно правильно (Бальцер, 1910). Мало того у реципрокных гибридов Strongylocentrotus × Sphaerechinus митозы идут совершенно различно. Когда отцом является Sphaerechinus, митозы идут неправильно, а когда отцом является Strongylocentrotus — правильно. Во всяком случае необходимо подчеркнуть важнейший факт, что в случае гибридов Strongylocentrotus × Antedon, т. е. морской еж × морская лилия, совершенно правильно могут итти митозы, в которых участвуют хромосомы, принадлежащие животным разных классов, и что неправильные митозы (если говорить пока о митозах соматических) «оставляют не такое частое явление. Нужно впрочем иметь в виду, что вопрос этот изучен у животных еще совершенно не достаточно.
На материале морских ежей, изученном Бальцером (1910), можно хорошо познакомиться с неправильностями гибридных митозов и с проблемой их причин. При опытах гибридизации 4 неаполитанских морских ежей, Echinus, Strongylocentrotus, Sphaerechinus и Arbacia
[40], выяснилось, что неправильные митозы возникают вполне закономерно, именно только тогда, когда отцовским видом является Sphaerechinus (рис. 94 и 47), но зато во всех случаях, т. е. во всех трех скрещиваниях Sph♂ × ♀Str, Sph♂ × ♀Ech и Sph♂ × ♀Arb. Во всех остальных скрещиваниях митозы идут нормально или почти нормально.
 Рис. 94. Элиминации хромосом у Str. ♀ × ♂ Sph. (слева) и Arb.♀ × Sph. ♂ (справа). (По Бальцеру.)
Рис. 94. Элиминации хромосом у Str. ♀ × ♂ Sph. (слева) и Arb.♀ × Sph. ♂ (справа). (По Бальцеру.)
Неправильности митозов выражаются в том, что начиная с метафазы наблюдается отставание некоторых хромосом. В то время как одни хромосомы успели разделиться и разошлись к полюсам, часть хромосом обнаруживает неспособность разделиться и либо они остаются на экваторе, либо, оторвавшись от одной из двух дочерних нитей, отходят с опозданием к одному из полюсов, либо, вовсе оторвавшись от веретена/остаются вне дочерних ядер, сохраняя в некоторых случаях способность к размножению, как было описано в главе VII. Рис. 47 представляет картину таких неправильных митозов. В результате этих неправильностей происходит элиминация хромосом, выпадение их из дальнейшего участия в жизни ядра, выпадение в цитоплазму с последующим разрушением.
Бальцер с доступной тщательностью изучил вопрос о том, 1) сколько хромосом подвергается элиминации и 2) какие именно хромосомы элиминируются.
Гаплоидные числа хромосом ежей следующие:
Strongylocentrotus purpuratus — 1.8
Echinus micrntuberrulatus — 18 или 9 (разные расы)
Sphaerechinus granularis — 20
Arbacia pustulosa — 20
У гибридов Sphaerechinus ♂ × ♀ Strongylocentrotus при оплодотворении число хромосом должно быть 18 + 20 = 38. Однако уже после первого деления их остается гораздо меньше (в среднем из 41 подсчета лишь 21,4) и приблизительно такое же число сохраняется и в дальнейшем: после второго деления — в среднем 21,0 и в разных дальнейших стадиях, до бластулы включительно, насчитано в среднем 21,1. Во всех 14 культурах Бальцера картина получалась одинаковой: принимая во внимание трудности подсчетов, можно считать, что у гибридов Spherechinus ♂ × ♀ Strongylocentrotus остается 21–22 хромосомы и соответственно 17 или 16 элиминируется. Таким образом замечательно, что остающееся число на 3–4 хромосомы больше гаплоидного числа Strongylocentrotus.
У гибридов Spherechinus
♂ × ♀ Echinus, где число хромосом должно было быть также 38, их остается после элиминации тоже 21–22. Наконец у гибридов Spherechinus
♂ × ♀ Arbacia должно было бы быть 20 + 20 = 40 хромосом, а после элиминации остается 22 хромосомы и элиминируется 18.
Какие же хромосомы элиминируются? Так как не все хромосомы могут быть узнаны точно под микроскопом, то этот вопрос может быть решен косвенно, несколькими путями: во-первых, можно проследить судьбу тех хромосом, которые могут быть всегда узнаны (например четыре самые длинные хромосомы Sphaerechinus); во-вторых, можно исследовать вариацию длины хромосом чистых видов и гибридов. Наконец можно сделать некоторые заключения по наблюдениям над многополюсными митозами (тетрасферами). Бальцер показал с высокой убедительностью, что элиминируются только хромосомы Sphaerechinus.
Таблица на стр. 253 показывает распределение хромосом по длине у чистых видов и гибридов.999 258

Длина хромосом определена в миллиметрах по рисункам при увеличении в 2270 раз.
Сравнивая распределение хромосом после элиминации у Sph.
♂ × ♀ Str. с распределениями хромосом у родителей, видно, что 1) самые длинные хромосомы Sphaerechinus несомненно элиминировались и что распределение хромосом гибрида почти точно совпадает с распределением хромосом у Strongylocentrotus, 2) только в классах 3–3,75 и 4–4,75 имеется избыток на 3 (10—7) и 1 (6–5), т. е. на 4 хромосомы. Отсюда можно заключить, что у гибрида элиминировались все отцовские хромосомы кроме 4 из числа наиболее коротких. Это отвечает тому, что плютеусы Spherechinus
♂ × ♀ Strongylocentrotus имеют чисто материнский характер с незначительными порой вариациями в отцовскую сторону (нужно помнить, что неизвестно, как долго сохраняются эти 4 отцовские хромосомы и не элиминируются ли они в дальнейшем).
Еще более убедителен гибрид Sphaerechinus
♂ × ♀ Arbacia. Самые длинные его хромосомы не выходят за класс 3–3,75, как и у чистой Arbacia. Следовательно здесь с уверенностью можно говорить, что 13 более крупных хромосом подверглись элиминации и очевидно у гибрида остались лишь 2 самые короткие хромосомы Sphaerechinus.
Причины элиминации сейчас конечно еще совершенно неясны. Здесь возможны различные гипотезы. Исходя из факта, что элиминация в данной форме происходит только в том случае, когда хромосомы Sphaerechinus попадают в яйца других видов, ее можно сопоставить с более медленным темпом, свойственным дроблению Sphaerechinus. Так как элиминация начинается с того, что хромосомы Sphaerechinus отстают в процессе расщепления от хромосом другого вида, то можно истолковать явления так, что хромосомы каждого вида имеют свой собственный темп деления, отвечающий темпу дробления. «Медленные» хромосомы, попадая в компанию с «быстрыми» в
«быстрое» же яйцо, естественно отстают, тогда как при реципрокном скрещивании отставания уже не будет, а будет, наоборот, некоторое забегание вперед, которое может не оказаться фатальным.
Такое объяснение встречает однако ряд трудностей. Так, темпы дробления Arbacia и Sphaerechinus повидимому совпадают. Также странно, что часть хромосом оказывается в состоянии перестроиться на новый тип дробления (ср. также благополучные скрещивания трех видов рода Fundulus, различающихся темпами дробления).
Также маловероятно, чтобы здесь какую-либо роль могли играть свойства центросом, например недостаток энергии для растягивания крупных хромосом и т. п. С другой стороны, Бальцер обращает однако особое внимание на свойства самих хромосом и в частности на их величину как на единственный пока признак, доступный более точному изучению. Анализируя только что приведенную таблицу, мы действительно видим, что как в случае гибрида Sphaerechinus ♂ × ♀ Strongylocentrotus, так и Sphaerechinus ♂ × ♀ Arbacia в элиминации хромосом обнаруживается очень простая закономерность — элиминируются все хромосомы, превышающие длину хромосом материнского вида. Для эмбриона, развивающегося из яйца Strongylocentrotus, хромосомы свыше 6,75 — предел длины хромосом самого Strongylocentrotus — оказываются уже чересчур длинными и не могут удержаться в митозах. Для яиц Arbacia, предел длины хромосом которого лежит уже при 3,75, «невыносимыми» оказываются уже хромосомы в 4. С другой стороны, для Strongylocentrotus оказываются «терпимыми» короткие хромосомы морской лилии Antedon несмотря на их чрезвычайную чужеродность. Таким образом можно предполагать существование некоторого нормального отношения, в котором должны находиться величина хромосом и окружающая их плазма.
Эти наблюдения для гибридагогии представляют конечно исключительный интерес, так как они, даже и не давая пока полного понимания явления, во всяком случае открывают возможность построения ряда рабочих гипотез, особенно учитывая возможности изменения длины хромосом рентгеном через транслокации. То, что величина хромосом оказывается действительно важным фактором, определяющим возможность сосуществования в общем правильном митозе хромосом различного происхождения — весьма правдоподобно. В этом отношении особенно поучителен конечно гибрид Sphaerechinus
♂ × ♀ Arbacia, элиминирующий все хромосомы Sphaerechinus кроме 2 самых мелких, соответствующих по размерам хромосомам Arbacia.
Возможно ли сохранение в гибриде отцовских хромосом в неактивном состоянии? Этот вопрос с точки зрения хромосомной теории наследственности парадоксален, но мы считаем возможным отвести его обсуждению место ввиду наличия некоторых недостаточно изученных моментов в кариологии гибридов.
Выше излагался процесс элиминации отцовских хромосом при гетерогенном оплодотворении, как то было показано Купельвизером, Бальцером и другими, что сопровождалось возникновением в результате гибридизации чисто матроклинного потомства.
В некоторых случаях однако (Годлевский, 1906, Моррис, 1914) подобная элиминация не только не была доказана, но, наоборот, собранные наблюдения говорили в пользу ее отсутствия, причем личинки оказывались попрежнему чисто матроклинными.
Годлевский (1909) наблюдал в скрещивании морской еж × морская лилия (см. выше, стр. 116) хромосомы, да и то не точно, лишь в первом дроблений. Однако при исследовании личинок уже на стадии гаструлы, протекавшей совершенно точно, как у ежей, он обнаружил величину ядер не только не вдвое меньшую, как должно было бы быть у гаплоидов, но как бы промежуточную между величиной диплоида-ежа и диплоида-лилии, имеющую ядра значительно более крупные.
Рис. 95 показывает эти размеры: рис.
А показывает величину ядер гаструл у чистых Echinus, рис.
С — величину ядер у Antedon, а рис.
В — величину ядер гибрида, имеющего впрочем чисто матроклинный фенотип. Подобная же разница обнаружена и у плютеусов — «гибриды» имеют несколько более крупные, но более малочисленные ядра, чем у чистых Echinus. Годлевский на основании этого заключил, что хромосомы Antedon принимают участие в образовании ядер эмбрионов, хотя тем не менее у гибридов этой генерации признаков Antedon не обнаруживается.
 Рис. 95. Величина ядер в гаструлах у Echinus (А) Antedon (С) и F1 (В).
Рис. 95. Величина ядер в гаструлах у Echinus (А) Antedon (С) и F1 (В).
Данные Годлевского в полной мере подтвердил Бальцер (1910). Им были оплодотворены спермой той же лилии Antedon яйца ежей Strongylocentrotus и Echinus (18-хромосомные) и прослежены митозы. Хромосомы Antedon (11 или вернее 12) — по крайней мере некоторые — оказались характерной двуплечей формы, не свойственной ежам (см. выше рис. 92), и на митозах ультрагибридов можно было совершенно уверенно наблюдать рядом хромосомы обоих типов. Никаких следов элиминации хромосом, комков хроматина в плазме и прочих неправильностей замечено не было, и на всем протяжении дробления число хромосом оставалось постоянным — 29–30.
Развитие гибридов по сравнению с контролем было сильно задержано.
На стадий бластулы наблюдалась остановка развития. Нередко гаструл не образовывалось вовсе, а бластоцели наполнялись комками клеток. Остановка могла продолжаться дня 2. Полученные плютеусы все имели чисто материнский тип. Величина их ядер оказалась неотличимой от диплоидных ядер чистых Echinus и Strongylocentrotus, Поэтому Бальцер присоединился к мнению Годлевского, что элиминации хромосом здесь не происходит и что в этих гибридных плютеусах хромосомы Antedon пребывают в совершенно латентном, инертном состоянии. Нужно все же иметь в виду, что хромосом типа Antedon в гаструлах никто констатировать не пытался.
Совершенно аналогичный случай, упоминавшийся выше, мы встречаем у рыб, по данным Моррис (1911). При скрещивании рыб Fundulus и Ctenolabrus она проследила хромосомы, обоих видов в ряде делений. Они резко различны по форме — палочкообразные у Fundulus и округленные у Ctenolabrus. Эту свою форму они точно сохраняют во всех последовательных делениях, доказывая, что хромосомы обоих видов (родов) одинаково размножаются и с совпадающими темпами распределяются по бластомерам.
Сама М. Моррис проследила развитие гибридов лишь до 12—24-часовой стадии, после, которой они отмирали, как впрочем вскоре (на 24-часовой стадии), отмирали и эмбрионы, чистых видов. Таким образом установить, по какому типу развивался эмбрион, Моррис не могла. Но Лебу (1912) с лучшей аквариумной техникой удалось воспитывать этих гибридов
[41] до месяца и более, так что они развивали сердце, кровеносные сосуды, глаза и пр., хотя и не вылуплялись никогда. Эти гибриды по Лебу были вполне матроклинными, так что Леб считал, что в данном случае имеет место лишь возбуждение яиц к развитию чужой спермой в качестве особого случая искусственного партеногенеза. Некоторые уклонения от нормы, получившиеся среди них, не носили ни в каком смысле гибридного характера и легко могли быть вызваны и у чистых Fundulus легкой прибавкой NaCN.
Если увязать исследования Моррис и Леба в одну цепь, то окажется, что 1) сперматозоид Ctenolabrus действительно оплодотворил яйцо Fundulus, 2) хромосомы Ctenolabrus прослежены ясно во всех делениях вплоть до той стадии, до которой исследование было проведено, и 3) гибриды оказываются матроклинными, откуда вытекает вывод, что хромосомы нормально размножаются, но на организм никакого действия не оказывают, становясь «инертными»
[42].
Однако такого вывода пока делать конечно нельзя, т. е. нельзя без дальнейшего считать условия опытов Моррис и Леба одинаковыми. Развитие гибридов у Моррис, начиная с 12 часов, обнаруживало характерные аномалии в виде образования гигантских клеток и гибрид погибал (по погибал и контроль!). У Леба же возможно действительно был партеногенез, почему личинки и жили благополучно до месяца. Тем не менее исследовать этот вопрос более тщательно представляло бы конечно огромный интерес для всей теории действия генов вообще и в гибридах в частности.
Второе поколение гибридов и дальнейшие поколения
Если по первому поколению гибридов животных мы уже сейчас. имеем обширный и часто чрезвычайно ценный фактический материал, то по дальнейшим поколениям — в противоположность ботаникам — зоологам почти нечего сказать. Причины этому многообразны, начиная с общих социально-экономических и кончая чисто биологическими. Факт бесплодия огромного большинства гибридов, о которых шла речь выше, стоит здесь на первом месте. Нежизнеспособность всех ультрагибридов, с которыми было проделано так много зоологических работ, мешает и здесь дальнейшей работе. Наконец и технические моменты, как неумение паше выращивать иглокожих дальше стадии плютеусов и значительные трудности выращивания до размножения многих рыб, малая плодовитость крупных животных и дороговизна работы с ними, большая редкость многих естественных гибридов, попадавших в руки зоологов уже в мертвом состоянии, — все эти причины приводили к тому, что до настоящего, времени мы имеем лишь единичные работы зоологов, простиравшиеся на второе или обратное, а тем более дальнейшие поколения гибридов. Наконец и из числа этих работ большинство не имеет большого научного значения либо по ничтожному количеству полученных особей либо по недостаточно точному и поверхностному анализу полученного материала.
До настоящего времени мы имеем сведения лишь о дальнейших скрещиваниях зубров, бизонов, туров, гаялов и гауров (Кюн в Галле, Филипченко и Иванов, Фортунатов), об обратном скрещивании яка × зебу × зебу (Завадовский) и некоторые другие данные об яках, разных видах крыс (Гагедорн), географических расах хомячка (Семнер), некоторые данные по собако-волкам, овцам и верблюдам (последние 4 случая являются скорее обычными расовыми скрещиваниями), работы Бонхота по скрещиванию кряквы с шилохвостями и некоторыми другими утками, данные о гибридах с канарейками (Промптов, 1928; Dunker, 1927) о гибридах Gallus bankiva × G. varius, разбросанные и часто малоценные данные о фазанах и др.
Ряд интересных работ касается скрещивания рыб Xiphophorus с Platypaecilus и несколько работ по бабочкам (Гарриса и др. — о Bistonidae, Штандфусса, Гольдшмидта — о сатурниях, Федерлея и Битинского-Зальц — о бражниках и др.), и работы по, скрещиваниям рас Drosophila pseudoobscura, которые могут быть также отнесены к числу гибридологических работ в нашем смысле.
Этот материал к сожалению по своей скудости почти ничего нового не может прибавить к тем идеям, которые разработаны в этой области ботаническими исследованиями, и для ряда таких идей наш материал не может служить даже хорошими иллюстрациями. Так, наиболее важный для всей гибридагогии пункт
о многообразии второго поколения межвидовых гибридов имеет скромный отзвук в нашем материале. Гагедорны (Hagedoorn A. L. а. А. С., 1922) изучили скрещивание крыс. Скрещенные виды или точнее разновидности Mus rattus из Америки, Европы, Сев. Африки, Явы и Суматры — очень близки друг к другу и различаются малосущественными признаками. Однако во втором поколении этих гибридов наступало чрезвычайно интересное и часто неожиданное расщепление. Как характерно для межвидовых скрещиваний например в роде Antirrhinum (Баур, Лотси), здесь F
2 на базе взрыва комбинаторики возникают закономерно такие формы, которые никак нельзя было предсказать или даже предположить. Как известно например в скрещиваниях Antirrhinum majus × mo lie появились формы, столь резко по устройству цветов отличающиеся от обоих родителей, что они оказываются более сходными с цветами растений уже иного рода Rhinanthus. В меньшей степени наблюдали также «взрыв» формообразования и появление непредсказуемых форм и Гагендорны; так, у них появились желтые крысы, белохвостые, пегие, шоколадные, танцующие.
Этот опыт среди зоологических принадлежит к числу крупнейших по количеству выращенных особей F
2, что и дало возможность проявиться здесь этим типичным особенностям F
2 межвидовых скрещиваний. В сходном по масштабу опыте Бонхота (Bonhote, 1902, 1905, 1907, 1909, 1915) по скрещиванию утки-кряквы с шилохвостью (и пестроносой уткой A. poecilorhynchus) благодаря наличию резких различий между скрещенными видами можно проследить также чрезвычайно важное явление комбинирования признаков разных видов. Так, изображенный на цветной таблице III гибрид кряквы и шилохвости (с примесью третьего вида) обладает признаками кряквы (зеленая голова, тогда как у шилохвости она коричневая), в соединении с острым и длинным хвостом, несущим ясные элементы хвоста шилохвости. Весьма разнообразные комбинации видовых различий наблюдались в скрещиваниях золотистого фазана о алмазным.
Таблица III
 Кряква (1), шилохвость (2) тройной гибрид (3). Комбинация видовых признаков.
Кряква (1), шилохвость (2) тройной гибрид (3). Комбинация видовых признаков.
Подобное же комбинирование видовых различий с резко выраженным расщеплением мы видим в обратном скрещивании F
1 зебу × як с зебу, изученном Завадовским. Здесь идет расщепление по черной и серой окраске, наличию и отсутствию горба, по узким и широким ноздрям, большей или меньшей оброслости и некоторым другим признакам, так что отдельные особи (братья и сестры) этого обратного скрещивания оказываются резко различными друг от друга. Несмотря на очень малое число особей для некоторых признаков можно уловить более или менее типичные менделевские расщепления типа 1:1.
 Рис. 96. Pulvillus меж коготков лапки у Deilephila galii.
Рис. 96. Pulvillus меж коготков лапки у Deilephila galii.
Правда, для настоящих межвидовых гибридов не менее характерно большее или меньшее
искажение правильных менделевских расщеплений, связанное с нарушением правильного гаметогенеза у гибридов и возникновением нежизнеспособных комбинаций. Однако для уловления таких искажений конечно необходим значительно больший материал, где могли бы проявляться законы больших чисел, лежащие в основе менделевских пропорций.
Такого сорта искажение мы можем видеть в скрещиваниях рыб пецилий и меченосцев, правда, преимущественно в связи с полом, о чем мы упоминали раньше» Так, здесь возникают в неправильных отношениях карликовые самцы, хотя гаметогенез здесь идет повидимому правильно. Неправильные диплоидные гаметы возникали у гибридов Pygaera, Biston и других бабочек, но с точки зрения судьбы менделевских картин эти скрещивания не были достаточно изучены.
Семнер (Sumner, 1915–1932), производя скрещивание географических рас американских хомячков Peromiscus, подверг тщательному изучению наследование различий между этими расами. Эти субгибридные скрещивания лишены некоторых характерных черт видовых скрещиваний — так, в них не возникало никаких нарушений плодовитости, неправильностей гаметогенеза, искажения пропорций полов и пр. Но, с другой стороны, эти скрещивания в высшей степени характерны для многих межвидовых в том отношении, что они имеют дело с типичными межвидовыми различиями, обусловленными в самых мелочах действием многих «малых генов», накопленных естественным отбором в процессе приспособительной эволюции. Семнер имел дело с географическими расами, приспособленными главным образом по окраске и размерам к различным экологическим условиям — к жизни на белом песке морского побережья, к жизни в условиях каменистой пустыни, в лесу и т. д.
Начиная свои многочисленные исследования, Семнер стоял на ламаркистской точке зрения, рассматривая приспособление как результат прямого действия внешних факторов — влажности, света и прочего на генотип, и считал, что наследование этих различий совершенно не подчиняется менделевским закономерностям. Однако в результате своих экспериментов он решительно изменил свои взгляды, и в последней итоговой работе (1932), излагая их, показал, что, наоборот, и для приспособительных различий географических рас законы менделевской наследственности имеют полную силу приложения. Еще несколько примеров покажут нам все характерные явления наследования межвидовых различий при нормальном гаметогенезе.
Битинским-Зальц и Гюнтером (1930) было изучено наследование видового различия в строении pulvillus в скрещивании бражников Deilephila galii и D. euphorbiae. У D. galii между коготками лапок находится особое образование в виде кожистой овальной подушечки, окруженной у основания крепким хитином. Поверхность подушечки покрыта мелкими киниками (рис. 96,
d). Этот признак имеет серьезное значение в систематике группы Deilephila. У euphorbiae подушечка почти не развита. Гибриды имеют промежуточное развитие этого органа, а в обратных скрещиваниях F
1 × galii и F
1 × euphorbiae возникает сильная изменчивость (рис. 97). Некоторые типы подушечки в обратных скрещиваниях, особенно F
1 × euphorbiae, носят очень своеобразный характер и могут уже считаться новообразованиями, «не предсказуемыми фенотипами», так как предсказать их появление на основании сравнительного изучения родительских видов совершенно невозможно.
 Рис. 97. Наследование pulvillus в F1 и в Fb Deilephila galii × D. euphorbiae.
a — galii, b — euphorbiae, с — F1, d — F1 × gаlii, e — F1 × euphor. (по Битинскому-Зальц и Гюнтер).
Рис. 97. Наследование pulvillus в F1 и в Fb Deilephila galii × D. euphorbiae.
a — galii, b — euphorbiae, с — F1, d — F1 × gаlii, e — F1 × euphor. (по Битинскому-Зальц и Гюнтер).
Таблица А (стр. 264) представляет собой попытку тех же авторов проследить наследование различий скрещенных видов бражников по окраске. Бабочки разбиты на классы, где класс I соответствует-чистым формам galii и классы VIII и IX — чистым euphorbiae. Классы II–VII отвечают различным переходным формам между чистыми видами. Если их взять в такой сложной оценке, то по ряду деталей окраски каждый класс не представляет вполне одинаковых форм, но объединяет особей, обладающих примерно одинаковой степенью отличия от чистых видов. Из таблицы видно, что по степени изменчивости, выражаемой сигмой, F
1 не отличается от чистых видов, но в F
b, возникает значительное многообразие. Можно ожидать, что если бы было поставлено скрещивание F
1 × F
1 и получено F
2, то изменчивость была бы еще более значительной и может быть вышла бы даже за пределы чистых видов. Очень сильную изменчивость по окраске гусеницы и длине рога наблюдал Мори (Могу, 1901) у гусениц природных гибридов Deilephila [(euphorbiae ♂ × ♀ vespertilio) x ♀ vespertilio].
Таблица А. Наследование окраски в скрещивании Deilephila euphorbiae × D. galii

К сожалению F
2 не было получено и в опыте Федерлея (Federley, 1927), откуда мы заимствуем данные о наследовании веса куколки (табл. В, стр. 264) в скрещивании бражников Deilephila (Chaerocampa) elpenor и D. (Metopsilus) porcellus. 05 изменчивости F
1 здесь трудно судить ввиду незначительности материала. Но зато в F
b мы видим огромное повышение изменчивости (σ = ±6,57 для самок) по сравнению с изменчивостью чистых видов (σ = ±0,80 и 2,34), причем в этом обратном скрещивании F
1 × elpenor возникают куколки, вес которых даже значительно превышает вес чистых elpenor. Федерлей (1929) отмечает в частности крупные размеры возникающих здесь сублетальных куколок.
Таблица Б. Наследование веса куколки в скрещивании Deilephila (Chaerocampa) elpenor × D. (Metopsilus) porcellus

Этим кратким изложением нам приходится исчерпать все существенное в области дальнейших поколений гибридов животных. Мы видим здесь сложнейшие, трудно поддающиеся обычному генетическому анализу расщепления с участием множества малых генов, скрывающие подстилающие их картины менделевских отношений. Тем не менее интересные (правда, пока лишь по своей иллюстративности) примеры возникновения в F
2 новых комбинаций видовых признаков заставляют нас желать как можно более быстрого накопления серьезного материала по работе с дальнейшими поколениями гибридов, ибо именно здесь мы стоим у начала грандиозной и увлекательной творческой работы по созданию новых форм животных для обогащения нашей природы и хозяйства.
Глава X. Темпы и направление гибридизационной работы
В настоящей заключительной главе мы должны осветить некоторые общие вопросы, связанные с работой по созиданию новых форм животных. К таким вопросам относится прежде всего вопрос
о темпах созидания новых форм, который в свою очередь связан 1) с вопросом о сокращении сроков смены поколений, 2) с вопросом об умении правильно предугадывать и рассчитывать результаты различных скрещиваний и 3) с вопросом о проектировании, с вопросом об умении четко наметить поставленную задачу и не итти в работе вслепую, стихийно. Эти вопросы имеют вообще говоря более широкое значение для всякой селекционной работы. Тем не менее, как было уже освещено в I главе, для гибридизации они приобретают специфический характер. К сожалению однако нужно констатировать, что эта глава должна явиться в значительной степени делом будущего, т. к. в настоящее время мы не располагаем вовсе необходимым для нее специальным конкретным материалом и вынуждены ограничиться лишь кратким очерком на базе косвенных материалов.
Ускорение смены гибридных поколений
Ускорение смены гибридных поколений представляет для нас вопрос исключительной важности, гораздо более острый, чем для обычной селекции. Эго связано, во-первых, с тем, что при созидании новых форм животных методами гибридизации совершенно неизбежны большие «отходы». Если например поставить себе задачу создать на базе гибридизации зубра с яком и туром новую форму домашних животных крупного размера с сильной оброслостью, ценной кожей и мясом, то совершенно ясно, что эта цель будет достигнута не скоро. Ясно, что интересные комбинации начнут появляться лишь начиная со второго-третьего поколения, что в них будут еще наряду с ценными сторонами иметься и отрицательные, устранение которых потребует опять не менее 2–3 поколений и т. д. И при этом придется подчеркнуть, что до тех пор пока необходимая комбинация не будет достигнута, все предыдущие поколения могут вовсе не представлять никакой народнохозяйственной пользы, быть «нерентабельными». Эта особенность резко отличает гибридизационную работу от обычной селекционной, которая в каждый данный момент работает с полезными, рентабельными животными. Поэтому как бы медленно селекционная работа ни шла, она вообще говоря в каждый данный момент выгодна. Гибридизационная работа, наоборот, становится большей частью выгодной только тогда, когда она достигает намеченной цели, когда она удается.
Отсюда ясна важность проблемы об ускорении работы гибридизатора вообще и в первую очередь о возможном ускорении смены поколений в частности-.
Это ускорение может быть достигнуто за счет ускорения эмбрионального и постэмбрионального развития вплоть до наступления половой зрелости. Для разных животных эти условия различны. У бабочек, у которых иногда имеются полугодичные диапаузы в развитии яиц или одно-двугодичные диапаузы в кукольной стадии, главнейшее ускорение может быть достигнуто за счет сокращения эмбриональных стадий и ничего нельзя достигнуть за счет стадии imago. У млекопитающих, у рыб, наоборот, на эмбриональном развитии можно выиграть пустяки и главнейшее внимание должно быть направлено на постэмбриональное развитие.
Большие перспективы в этом направлении безусловно открываются эндокринологией. Уже клинические наблюдения над человеком знакомят нас со случаями исключительно раннего созревания как мальчиков, так и девочек, достигающих половой зрелости к 4–8 годам, т. е. в 2–3 раза быстрее нормы при некоторых нарушениях гормональной системы, вызванных заболеваниями гонад. Такое же ускорение созревания в последнее время достигалось инъекцией пролана, гравидана. Скадовскому в 1932 г. удалось получить в аквариумах ранней весной икру у окуней, вообще не размножающихся в аквариумах, и получить мальков. Вызывание течки у неполовозрелых мышей в течение трех дней является сейчас общеизвестным лабораторным приемом. Пути, которыми здесь приходится итти, будут вероятно очень сходны с теми, которые мы обсуждали выше в главе, посвященной проблеме борьбы с бесплодием гибридов. Поэтому здесь мы можем быть краткими.
Наиболее радикальными являются перспективы трансплантации молодых гонад во взрослый организм. Такой опыт был проделан покойным И. Г. Коганом, и насколько нам известно, нигде не был опубликован. Он брал семенники у только что вылупившихся цыплят и трансплантировал их в гребень взрослому петуху. Чрезвычайно быстро — насколько мы помним — в 2–3 педели в этих семенниках уже можно было видеть готовые сперматозоиды. Если усовершенствовать эту операцию так, чтобы сперматозоиды могли быть использованы для искусственного осеменения, то поколение кур могло бы сократиться через мужскую линию до 35–40 дней и в течение сезона можно было бы сменить 2–3–4 поколения, что имело бы конечно огромное значение. Еще больший эфект получился бы от такого раннего получения спермы быка, где срок смены поколения через мужскую линию сократился бы почти лишь до длительности беременности.
Правда, в разработке этого метода встретятся препятствия в виде неприживления трансплантата. Как показали опыты Л. Леба, трансплантация у млекопитающих легко удается в пределах имбридированной линии, а гетеротрансплантации по богатому опыту сильно затруднены, У гибридов и в их расщеплениях мы вероятно будем встречаться с условиями, близкими именно к гетеротрансплантациям, и придется разрабатывать более сложную методику. Для нас ясно однако, что, мобилизуя все учение о трансплантациях и эксплантациях, мы должны сосредоточить внимание на разработке этой методики, так как, разрешая радикально проблему ускорения гибридизационной работы, мы сделаем ее и гораздо более эфективной и даже гораздо более" интересной, сократив в ней утомительные моменты длительного пассивного ожидания достижения гибридами-самцами половой зрелости. От самого гибридизатора будет зависеть уже придать его работе тот или иной темп.
Проблема предсказания
Об этой проблеме мы уже немного говорили в первой главе. Сейчас мы можем вернуться к ней с некоторым новым материалом. Для успешности, быстроты и экономности работы нам. конечно чрезвычайно важно научиться и иметь возможность предсказывать или хотя бы достаточно точно предугадывать возможные результаты своих скрещиваний. Для этого возможны различные отправные точки. В предыдущей главе мы обсуждали явление атавизации гибридов, дающее в наши руки одну из таких отправных точек. Включая в свой проект соединение признаков разных видов, мы можем изучить эволюционный характер этих признаков — являются ли они специфическими узковидовыми или более широкими. В первом случае мы будем иметь
больше шансов, что избранные нами для соединения признаки окажутся рецессивными и в первом поколении исчезнут, а во втором будут извлекаться в лучшем случае лишь у 25 %, а при полигенности и еще реже — в 6 %, в 1,5% и т. д. с коефициентом прогрессии в
1/
4 (
1/
4,
1/
16,
1/
64 и т. д.) или 100%:4
n, где n — число генов. Хотя, как мы видели, основанные на подобных соображениях расчеты и не всегда будут оправдываться, тем не менее во многих случаях они будут верны.
Значительно более возможностей для предсказания дает экстраполяция со скрещивания
А ×
В на другие скрещивания
А ×
С, А × D… Так, скрещивание петуха с цесаркой показало почти полное исчезновение гребня у F
1. Отсюда мы, экстраполируя, можем уже с большей уверенностью предполагать, что и в других скрещиваниях петуха — с павлинами, фазанами и пр. — гребень должен также исчезнуть. В частности когда мы ожидали появления первых гибридов курицы с павлином, то настаивали на том, что- только тех птенцов, которые окажутся без гребней, можно будет признать за гибридов, а цыплята с гребнем будут указывать на их негибридное происхождение, что и оправдалось.
Несравненно важнее однако проблема предсказаний явлений в дальнейших поколениях. Они возможны только на базе знания частной генетики скрещиваемых видов. Тем самым только в процессе самих скрещиваний создаются условия, позволяющие рационализировать дальнейшую работу. Поэтому
гибридизация должна на каждом шагу сопровождаться самым напряженным генетическим анализом, захватывающим гибридов возможно более всестороннее. Даже на сравнительно небольшом числе животных, как например в случае скрещивания зебу с яком в Московском зоопарке, можно хотя бы предварительно сделать ряд заключений о генах, принимающих участие в расщеплении. И чем всестороннее эти немногие особи будут изучены, тем о большем количестве генов можно будет составить предварительное представление, которое можно будет положить в основу расчетов дальнейшей работы с данным гибридный материалом.
Эту мысль мы подчеркиваем здесь для того, чтобы отметить совершенно неудовлетворительное изучение, которому подвергаются обычно гибриды. Как правило от них сохраняются только музейные чучела и все богатство анатомических, гистологических, физиологических, рефлексологических и прочих признаков оказывается утерянным. Несмотря на тысячи — и многие тысячи — гибридов, прошедших через руки даже зоологов-ученых, мы имеем например совершенно единичные описания скелетов, да и то лишь черепов; совершенно не имеем описания мягких органов — сердца, легких, желез и пр. А между тем даже десяток особей второго поколения, изученных подобным образом во всех отношениях, мог бы существенным образом пополнить наши представления о генных ресурсах и дать возможность предсказывать, что можно извлечь в дальнейшем из такого скрещивания.
Предсказание по данным изменчивости. В качестве примера хода подобного предсказания рассмотрим следующий. Пусть мы скрещиваем два вида
А и
В в нас интересует рост. Для обоих видов нами изучены их средняя высота и вариация.
М
А σ
А
М
В σ
В
Получив F
1 и не очень большое F
2, мы видим, что и то и другое имеет более или менее промежуточные между
А и
В размеры, и нас начинает интересовать вопрос, каковы дальнейшие перспективы этого скрещивания? Может ли оно нам дать что-либо более интересное, чем то, что мы уже имели у чистых видов
А и
В и их гибрида F
1?
Для ответа мы должны изучить величину. сигмы F
2. Эта величина будет иметь для нас очень большое значение, так как она дает вполне обоснованный ответ на поставленный вопрос. Мы знаем, что величины М и σ описывают характер изменчивости обычно с удовлетворительной полнотой. Можно утверждать, что при достаточном объеме нашего опыта мы получим в F
2 в качестве самых крупных особей
[43] имеющих величину
M
F2 + 3,
5σ
F2
Так как величины M
F2 и σ
F2 нами определены по уже приученному материалу F
2, то мы можем ответить и на вопрос, чего мы можем ожидать от нашего скрещивания. В частности если М
A>МB то необходимо сравнить две величины, определив разность
(M
F2 + 3,
5σ
F2) — (M
A + 3,
5σ
A).
Если эта разность положительна, то положителен и наш опыт-(в данном отношении), так как крайние плюс-варианты из F
2 будут крупнее крайних плюс-вариантов вида
А. Мы получаем даже еще более конкретный ответ: 1) насколько мы выигрываем в этом скрещивании и 2) какое количество особей в F
2 необходимо получить, чтобы добиться заданного превышения.
Конечно подобные расчеты нельзя применять механически… Необходимо более вдумчиво рассмотреть характер распределения вариаций в F
2, эксцессивность или многовершинность его, асимметрию и пр., чтобы иметь возможность оценить приложимость кривой Гаусса и основанных на ней приводимых расчетов. Но в эти детали, не носящие уже специфического характера, мы здесь можем не входить.
Подобным же образом могут быть оценены и шансы достижения комбинации двух (и более) признаков, — Здесь уже необходимо изучение корреляции рассматриваемых признаков α и β и построение для них корреляционного поля, на которое одновременно может быть нанесено и поля родителей. По подобному чертежу можно сделать выводы и о том, какая может быть получена наиболее желательная комбинация двух данных признаков и какое количество особей нужно получить для реализации этой комбинации.
При всяких подобных вычислениях нужно всегда иметь в виду учение о статистических ошибках, определяющих точность выводов, и в данном случае уверенность делаемых предсказаний. В частности вычисляемые для второго поколения величины M
F2 и σ
F2 будут том точнее, чем на большем материале они будут вычислены. Соответственно и предсказания будут точнее, и их точность по общему закону будет пропорциональна корню квадратному из числа особей. Увеличивая число особей в четыре раза, мы повышаем точность вдвое (вдвое уменьшаем величину ошибки). Однако только по этим данным и могут быть достаточно правильно оценены перспективы данного скрещивания. Так, известно по опытам биологов, что при скрещивании двух видов в F
2 идет столь сложное-расщепление, что даже при довольно значительных количествах-особей F
2 не удается извлечь формы, даже повторяющие свойства родителей, а не только выходящие за их пределы. Тем не менее, вычисляя сигму, можно будет с уверенностью предсказать, что выход за пределы родительских форм все же может быть достигнут, хотя бы при помощи дальнейших скрещиваний.
Во всяком случае важно иметь в виду следующее: как бы ни был обширен эксперимент, в F
2 межвидового скрещивания мы получаем лишь
ничтожную долю генотипов, возможных к по-лучению в этом скрещивании. Поэтому-то перед нами и стоит ответственная задача по. этой
ничтожной доле суметь оценить дальнейшие перспективы данного скрещивания. Нужно иметь в виду, что уже при различии между видами в 10 генах в F
2 должно возникать 1024 комбинации генов и 59049 различных генотипов (2
10 и З
10)! Между тем при скрещивании таких сравнительно близких видов растений, как Antirrhinum majus и A. molle, Баур считает, что в F
2 происходит расщепление примерно по 100 генам, т. е. одно число возможных комбинаций значительно превышает величину с 30 нулями (1 000 000 000 000 000 000 000 000 000 000).
Между тем и эта величина очень мала по сравнению с фактической. Если например у коровы 30 пар хромосом, то различие
в 100 генах достигается уже тогда, если в каждой хромосоме коровы всего лишь 3 гена будут отличаться от генов например бизона. На самом деле вероятно каждая хромосома коровы от каждой хромосомы бизона отличается гораздо большим числом генов.
Но даже если число 2
100 уменьшить в миллиарды раз, то и тогда будет ясно, что даже при тысячах особей F
2 это будет лишь
ничтожной долей тех бесчисленных комбинаций, которые могут возникнуть. А поэтому умение оценить по этим намекам действительно возможные перспективы скрещивания чрезвычайно важно. Здесь одинаково важно 1) во-время определить, что в данном скрещивании нет интересных перспектив и что следовательно с ним не стоит дальше работать, не стоит затрачивать на него силы и средства и 2) оценить, что можно ожидать от скрещивания даже тогда, когда первые сотни особой F
2 еще не дают чего-либо интересного.
Проблема проектирования
Проектирование новых форм животных, как мы кратко изложили в. первой главе, должно будет явиться той руководящей теорией, которая станет стержнем гибридагогии. Правда, мыслима гибридизация по «принципу» — скрещивай все и вся, авось что-нибудь ценное и получится. Но это конечно не принцип социалистического, планового хозяйства и науки. Мы должны суметь поднять теоретический уровень работы настолько, чтобы быть в состоянии четко и конкретно ставить себе задачи и прямо и с максимальной производительностью труда и экономией сил и средств эти задачи разрешать. Для этого мы и должны научиться проектировать новые формы живых существ, как бы трудны эти работы ни были и какого бы всеобъемлющего синтеза биологических, а частично и других знаний они ни потребовали.
В проектировании новых форм животных можно различить три ступени:
1) народнохозяйственный эскиз-заказ;
2) морфо-физиологический проект фенотипа;
3) генетический рабочий план построения генотипа.
Народнохозяйственный эскиз-заказ должен определить основной смысл и направление работы по созиданию новой формы животных и дать основные показатели, которым должна удовлетворить проектируемая новая форма для того, чтобы она смогла занять свое место в природе или в хозяйстве человека.
Созидание новой формы животных может быть заказано для различных целей — для разведения в домашних условиях. для пищевых или сырьевых целей, для тяговой силы, для декоративных целей и пр. Можно дать задание создания новых форм рыб для новых огромных искусственных водоемов, возникающих в связи с постройкой гидроэлектростанций, и пр. Новые формы могут быть заказаны для заселения экологически-специфических районов дальнего севера, песчаных пустынь, горных районов, для вытеснения из фауны нежелательных и вредных элементов ее, для использования остающихся неиспользованными тех или иных естественных ресурсов и т. д. Несмотря на то, что до сего времени подобные мысли еще очень редко возникали в уме хозяйствующего человека — теперь, когда природа наконец получила своего истинного единственно достойного хозяина — победоносный пролетариат и колхозное крестьянство, — проблема обогащения фауны и коренной реконструкции животных форм несомненно встанет во весь рост.
Сформулировать поставленную задачу — для каких целей нужно создать животное, для какого района и климата, для каких основных условий питания, какие необходимы темпы роста и размножения, форма и объем продукции и оплата корма — такие и подобные вопросы должны быть продуманы и проработаны для того, чтобы мы могли иметь действительно серьезно обоснованное задание, на выполнение которого можно и должно затрачивать силы. Эта работа конечно не может вестись абстрактно — подумал да и придумал заказ: «хорошо бы вот создать таких-то животных», но в основном должны исходить из
критики существующих уже видов животных, оценки их места и роли в сельском хозяйстве и в природе, на суше и в воде и оценки их недостатков с той или иной точки зрения. Исходя из такой критики, сравнивая друг с другом различные виды, начиная с ближайших друг к другу и постепенно расширяя сравнение, можно будет и разработать те эскизы-заказы, которые должны будут сосредоточить на себе дальнейшее внимание гибридагогии.
Морфофизиологический проект фенотипа является в проблеме проектировании нового животного наиболее трудной ступенью, так как он требует высшего синтеза всех анатомических, физиологических и экологических знаний. Здесь должны быть проработаны основные физиологические показатели — особенности обмена веществ, необходимые. для обеспечения заданных темпов роста, размножения и продукции. Должны быть продуманы необходимые особенности устройства различных органов, начиная с сердца, кровеносной и дыхательной системы, диаметры сосудов, гистологическое строение, особенности эндокринной системы и пр., и в результате решен вопрос о том, может ли быть реализован проект в качестве жизнеспособного животного и каковы должны быть отличия запроектированной новой формы от существующих, ныне живущих.
Для полного обеспечения подобной работы необходимо ближайшее использование и мощное дальнейшее развитие
сравнительной физиологии и сравнительной анатомии, развитых под углом зрения этих новых заданий. В настоящее время яти науки — еще мало развитая сравнительная физиология и односторонне развитая сравнительная анатомия — являются почти исключительно «спутницами» эволюционной теории. Сейчас, когда на база селекции и гибридизации эволюционная теория резко активизирована, привлечена не только к объяснению мира, но и к переделыванию его, должны совершить такой же поворот и ее «спутницы», должны мобилизоваться для того, чтобы помочь строить или по крайней мере проектировать новые формы животных.
Нельзя сказать, чтобы выдвигаемые здесь вопросы были совершенно новы. Наоборот, уже в современной зоотехнии имеется например детально разрабатываемое так называемое
учение об экстерьере, а в последнее время начало разрабатываться — хотя еще и очень слабо —
учение об интерьере. Правда, капиталистическая идеология наложила на учение об экстерьере свою тяжелую печать и в нем наряду с серьезными научными элементами имеется еще достаточное количество хлама, предрассудков, связанных либо с невежеством либо с частнохозяйственными торговыми отношениями. Кроме того здесь имеется еще очень слабая связь с физиологией, так что учение об экстерьере носит преимущественно морфологический характер несмотря на то, что все тезисы этого учения ссылаются именно на связь той или иной особенности экстерьера с особенностями физиологической продуктивности — связь, большей частью мало или даже нисколько не доказанную, вроде знаменитых требований «живых глаз» от молочной коровы, «мужественной головы» от быка и пр.
Другим недостатком современного учения об экстерьере является ограничение ого кругозора только основными видами одомашненных животных. Уже оценка например зубра или яка столь решительно выходит за рамки понятий этого учения, что они вряд ли могут попасть в число «нормальных» форм, а но крайних уродов.
Поэтому только решительный синтез 1) зоотехнического учения об экстерьере и интерьере, 2) антрополого-медицинского учения о конституциях, 3) сравнительной физиологии, 4) сравнительной анатомии, 5) экологии, 6) генетики, при распространении учения и на дикие виды — сможет дать тот необходимый комплекс знаний, который обеспечит научную критику ныне живущих животных и разработку теории проектирования новых животных и возможность самого проектирования.
В самом проектировании конечно различные главы будут представлять очень различные степени трудности. Вероятно наиболее легко будет проектировать
общие пропорции тела, размеры конечностей и их строение, то, что более разработано и в учении об экстерьере зоотехниками. Роль этих элементов может быть легче понята и экологически. Интересны например наблюдения над значением длинных ног курдючных овец в экологической обстановке Средней Азии, где овечьи стада должны совершать большие переходы. По наблюдениям, сообщаемым Кашкаровым, курдючные овцы проходят расстояния в три раза быстрее, чем то могут сделать линкольны, и при переходе через горные речки делают в три раза более длинные прыжки.
Гиппологи дают еще более серьезную оценку роли длины отдельных костей ног — системы рычагов, работающих во время движения лошади. Распространение подобного изучения и на других животных, например антилоп, оленей и пр., а вместе с тем одновременно и на строение и гистологические и физиологические особенности мускулатуры позволило бы построить общую теорию проектирования конечностей.
В настоящее время имеется уже довольно интересный материал, относящийся к
сердцу, на рассмотрении которого можно составить себе общую картину того, какими должны быть те пути, на которых может быть разработана теория проектирования животных. Остановимся в частности на вопросе
о размере сердца, необходимом для организма. Этот размер принято выражать promille, т. е. в числе граммов веса сердца, приходящихся на 1 кг веса тела;
У различных животных этот параметр колеблется весьма сильно, от 0,15‰ у некоторых рыб до 17‰ и выше у некоторых птиц. Если следовательно мы должны запроектировать вес сердца для нашего проектируемого животного, мы должны наметить какую-то цифру в этих пределах. Чем же руководиться в уточнении проектируемого параметра? На этот, вопрос мы можем извлечь ответ из сравнительного изучения ныне живущих форм.
Работа сердца тесно связана с выработкой Энергии. Количество энергии, которую должен выработать организм животного, зависит от структуры расходного баланса ее. Здесь главными статьями будут: 1) поддержание температуры тела, 2) — выработка энергии для работы мускулатуры: а) в покоящемся состоянии, б) для движения, 3) для передвижения крови по кровеносной системе и пр.
Наибольшее значение (у теплокровных) имеет расход энергии на поддержание температуры тела, что доказывается резкой разницей веса сердца у холоднокровных и теплокровных. У рыб вес сердца не поднимается выше 2–2,5‰, у теплокровных это является уже нижним пределом. Так как температура тела — раз она должна поддерживаться на уровне более высоком, чем температура окружающей среды, — зависит а) от выработки тепла и б) от отдачи тепла, то факторы, влияющие на отдачу тепла, тем самым должны определять и выработку тепла. В отдаче тепла прежде всего играет роль отношение поверхности тела к массе его. Как известно, отношение поверхности к массе обратно пропорционально линейным размерам. Следовательно при проектировании величины сердца надо задавать его относительную величину в некоторой обратной пропорциональности с размерами. Функцию эту, следуя Гессе, можно приближенно определить эмпирически, пользуясь данными такого рода:
1. Изменением (падением) удельного веса сердца с возрастом:
Кролик новорожденный — 5,85‰
Кролик 2-недельный — 3,91‰
Кролик 4-недельный — 3,77‰
Кролик взрослый — 2,74‰
Цыпленок новорожд. — 9‰
Корица-молодка — 6,7‰
Курица взрослая — 6,3‰
2. Сравнением (падением с увеличением размеров) сходных видов различных размеров, например:
Домовый сирин весом 170 г — 8,25‰
Неясыть весом 441 г — 5,07‰
Филин весом 1 875 г — 4,70‰
Можно установить далее, что значительную роль играет степень подвижности вида, т. е. расход энергии по статье «Движение». Степень влияния этого фактора можно видеть из таких сопоставлений:
Рыбы
1. Угревые морские рыбы, поджидающие добычу, закопавшись в песок, Ophisurus, Sphagebranchus — 0,15-0,28‰
2. Придонные рыбы Uranoscopus, Trachinus — 0,52-0,62‰
3. Свободно плавающие, нехищные рыбы — 0,60-0,80‰
4. Быстро двигающиеся хищники, макрелевые — 1,60-2,12‰
Так как при подобном сопоставлении картина может осложняться разницей в величине тела, то лучше подобрать формы примерно одного веса.
Вес 20 г
Домовая мышь — 6,85‰
Лесная мышь — 7, 16‰
Нетопырь — 10,00‰
Ласточка деревенская — 14,50‰
Вес 65 г
Крот — 6‰
Удод — 12‰
Дятел большой пестрый — 17,26‰
Конечно к таким сопоставлениям, особенно когда сопоставляются в общем ряду даже птицы и млекопитающие, нужно относиться о-большой продуманностью и располагать материалом не отрывочным, но обширным, систематически обрабатываемым по, строго однообразной методике и статистически правильно. Точно так же и такие например параметры, как «подвижность», «теплоотдача с единицы поверхности» и пр., должны быть определены, не на глаз, а точными методами физиологии.
Например Рубнер тщательно сравнил выработку тепла у собак различного веса и получил такое сопоставление:

Здесь мы видим, что выработка калорий тепла на 1 кг веса оказалась действительно строго пропорциональной поверхности тела, приходящейся на 1 кг веса тела.
При наличии цифр, собранных со всей необходимой точностью, мы получим возможность из сопоставления различных животных друг с другом извлечь очень много данных, необходимых для критики и проектирования.
Если какое-либо животное не будет укладываться в ряд, мы должны будем искать какой-нибудь новый специфический фактор: если равен вес и равна поверхность, то надо будет обратить внимание например на характер волосяного покрова и учесть его роль. Далее могут быть вскрыты уклонения, для объяснения которых придется изучить роль например гистологии сердечной мышцы, частоту пульса, поверхность кровяных шариков, — особенность строения и ветвление артерий и т. д. Шаг за шагом должно быть выяснено, как отражаются на величине сердца различные анатомические, физиологические и экологические факторы.
С другой стороны, такому же анализу должны быть подвергнуты и отдельные элементы среды и той экологии, для которой мы: будем проектировать животное. Если например перед нами ставится задание спроектировать корову для Якутской области, то-мы должны будем представить экологию коровы в условиях Якутской области тоже в определенных параметрах. «Теплоотдача с единицы поверхности» в Якутской области не та, что в Крыму, и ceteris paribus сердце якутской коровы должно быть соответственно больше, чем крымской коровы, и притом на величину, стоящую-в функциональной зависимости от средней температуры Якутии и от других условий, например влажности воздуха, и т. д.
В соответствии с климатом должно стоять и общее развитие поверхности тела — общая «Коренастость» или «стройность», развитие ушной раковины и т. п. Прилагаемые схематические рисунки африканской антилопы Lithocranium и тибетской Pantholops (рис. 98) и контуры голов северного песца, лисицы и африканского фенека (рис. 99) иллюстрируют эту закономерность. Вместе с другими подобными же эта закономерность должна быть использована при разработке теории проектирования животных.
 Рис. 98. Антилопы Pantholops (слева) и Lithocranium (справа) (по Гессе).
Рис. 98. Антилопы Pantholops (слева) и Lithocranium (справа) (по Гессе).
 Рис. 99. Головы песца (а), лисицы (b) и фенека (с) (по Гессе).
Рис. 99. Головы песца (а), лисицы (b) и фенека (с) (по Гессе).
Наиболее трудной, но наиболее важной частью проекта фенотипа будет конечно его физиологическая часть, так как в настоящее время наши сведения даже по домашним животным еще очень скудны, а по диким животным отсутствуют почти нацело, да и методы изучения здесь конечно гораздо более сложны. Но и здесь начинать следует с собирания для каждого гиркона прежде всего количественных показателей всевозможных физиологических процессов — параметров скорости роста, частоты пульса, дыхания, органов выделения жидкостей, скорости кровообращения, данных газообмена, скоростей пищеварительных процессов, составных частей крови и т. д. Изучив их на большом числе видов гиркона ж сопоставив с различными анатомическими параметрами и экологическими параметрами вида, мы будем иметь те данные, на основе которых можно будет запроектировать
и каркас физиологических параметров, согласовывая его элементы так, чтобы иметь жизнеспособную схему.
Нужно иметь в виду, что то, что мы называем проектом нового животного, конечно невозможно будет особенно в ближайшее время составить заранее в законченном виде. Созидание новых животных методами гибридизации будет всегда длительным, многолетним процессом. И в самом ходе этой многолетней работы, к тому же не имеющей конца, будет достаточно времени для внесении различных конкретизаций и дополнений, так как даже в процессе построения какого-нибудь нового завода — задачи, гораздо более посильной современной технике и науке, — и то всегда вносятся поправки, вовсе не снимающие однако важности составления исходного проекта при начале работы. Таким образом вокруг каждой работы по созиданию нового вида животных-должна будет развернуться специальная научно-исследовательская работа, в процессе которой и будут разрешаться многочисленные вопросы, только самый небольшой перечень которых мы здесь дали.
Генетический рабочий план осуществления проекта и в то же время генетический проект должны будут указать, какие гены или группы генов должны быть заимствованы из генотипов таких-то именно видов и рассчитать вес необходимые для этого скрещивания, их последовательность, масштабы и т. д. В тех случаях, когда выбор видов, возможных для введения и скрещивание, мал, будет прост и план работы, но при богатстве гиркона видами составление такого плана явится чрезвычайно ответственным делом; так как от его качества будет в высокой степени зависеть тот срок, в течение которого сможет быть разрешена поставленная задача. Какие-либо общие указания однако здесь вряд ли можно дать, по. крайней мере в настоящее время, при отсутствии развернутых гибридизационных работ, а следовательно и опыта, и при полном нашем незнании генетики диких видов.
ПАВЛИН, ЦЕСАРКА И ИХ ГИБРИД.
Список литературы
Глава I
Серебровский А. С. — 1933 — Гибридизация как наука, Труды Инст. с.-х. гибридизации и акклимат. животных, т. I, стр. 20–36.
Глава II
Ackermann A. — 1898 — Tierbastarde. Zusammenstellung der bisherigen Beobachtungen über Bastardierung im Tierreiche nebst Litteraturnachwcisen. I. Wirbellosc, XLII, S. 103–121, 1896/97, II. Wirbeltiere, XLIII, S. 1 — 79, 1897/98. Abhandlungcn und Bericlitc des Vereincs für Naturkunde in Kassel.
Antonius O. — 1922 — Grundzüge einer Stammesgeschichtc der Haustiere — Jena, Fischer.
Baur E. — 1932 — Arlumgrenzung und Artbildung in der Gattung Anthirrinum, Zschr. ind. Abst. Ver., Bd. LXIII, S. 256–302.
Bonchote I. L. — 1902–1909 — On Hybrid Ducks, Proc. Zool. Soc., p. 318, 1902; p. 147, 1905, p. 598, 1909.
Bonchote I. L. — 1907 — On Hybrid Ducks, Proc. IV Int. Ornitli. Congr. p. 235, 1905.
Boyd M. — 1914 — Crossing Bison and Cattle, J. of Hered., v. 5.
Cronau C. — 1902 — Jagdfasan.
Dubois G. — 1925 — Affinités et origines du furet domestique. — Feuille des Natural., Paris, 46.
Fischer F. — 1880 — Deutsche ldw. Presse, VII, № 40, Berlin.
Fitzinger — 1859/60 — Über die Rassen des zahmen Schafs, Sb. mat. nat. kl. k. Ak. Wise., Wien, Bd. 38–41.
Ghigi A. — 1922 — L’hybridisme dans des génèses des races domestiques d’oiseaux, Genetica, Bd. IV.
Hagen H. — 1858 — Bastardzeugung bei Insecten, Stettiner Ent. Zschr., XIX, 41, 231, 407.
Hilzheimer M. — 1908 — Beitrag zur Kenntnis der nordafrikanischen Schakale, Zoologica, Bd. 20,
Hilzheimer M. — 1912/13 — Üeberblick über die Geschichte der Haustier-forschung, Zool. Annal., Bd. V.
Hilzheimer M. — 1926 — Natürliche Rassengeschichte der Haussäugetiere, Berlin, De-Gruyter.
Hopkinson E. — 1926 — Records of Birds breed in captivity, London, Witherby.
House H. — 1842 — Notice on the occurence of Hybrid Individuals occuring in the genus Smerinthus, Trans. Ent. Soc., III, 1841–1843 (ib. in Stainton Weekly Intelligencer, 77, 1858).
Keller C. — 1903 — Zur Abstammungsgeschichte unserer Hunderassen, Vierteljalirsschr. Naturf. Ges., Zürich, Bd. 48.
Klatt B. — 1927 — Entstehung der Haustiere, Handb. der Vererbungswiss., Bd. Ill, Lief. 2 (III. K.).
Kysela E. — 1909 — Über die Hybriden und ihre Zucht, Polyxena № 3, 4. 5, 6, 7, 8, 9, 10; 1910. № 2.
Lönnberg E. — 1919 — Remarks on some South American Canidae, Arch. Zool., Bd. 12.
Letsy I. P. — 1922 — Die Aufarbeitung der Kuhn’schen Kreuzungsmaterials im Institut für Tierzucht der Universität Hale, Gcnetica, 4.
Nabours R. K. — 1913 — Possibilities for a new breed of cattle for the south, Amer. Breed. Mag., 4.
Radde G. — 1884 — Ornis caucasica, Kassel.
Rusconi Mauro — 1840 — Über künstliche Befruchtung von Fischen und über einige neue Versuche in betreff. künstlicher Befruchtung an Fröschen. Müllers Arch, für Anat. und Physiol.
Schäme R. — 1911 — Die statischen Variationen des Hundeschädels, Zschr. f. Tiermed., Bd. 15.
Schäme R. — 1922 — Die Grundformen des Haushundschadcls, Jahrb. f. Jagdkunde, Bd. VI..
Schlagintwoit R. — 1857 — Bericht über einige Thiergattungen Tübet's, Das Ausland, Oct., S. 948.
Schufeldt R. W — 1890 — Note on the trunck sceleton of a Hybrid Grouse The Auk., July, v. X, p. 281–285.
Sewerzow N. — 1883 — Ein Bastard von Anas crecca mit A. boschas, Bull. d. soc. Nat. de Moscou, S. 1.
Sokolowsky A. — 1909 — Löwen-Tiger-Bastarde. Natur. und Haus, XVII, 305.
Standfuss M. — 1896 — Handbuch der paläarktischen Gross-Schmetterlinge für Forschen und Sammeln (русский, перевод: Штандфусс, Жизнь бабочек, изд. «Просвещения»).
Studer Th. — 1903 — Über den deutschen Schäferhund und einige kynologische Fragen., Mitt, naturf. Ges., Bern.
Westwood J. O. — 1842 — Description of a Hybrid Smerinthus with remarks on Hybridism in general, Trans. Ent. Soc., Ill, 195.
Wohltmann F. — 1911 — Das Landwirtschaftliche Institut der Universität zu Halle a. S., seine Entwickelung und Neugestaltung, Kühn’s Arch., Bd. I, erstes Halbband.
Грим О. — 1881 — Заметка о скрещивании рыб, Тр. СПБ, Общ. Ест., т. ХП.
Колодько М. Н. и
Фортунатов Б. К. (ред.) — 1928 — Степной заповедник Чапли-Аскания Нова, Гиз.
Постановления Коллегии НКЗ СССР о гибридизации, Бюллетень Всес. научно-исслед. инст. животноводства, № 3–4, стр. 87, 1931.
Серебровский А. С. — 1929 — Опыт качественной характеристики процесса органической эволюции, Естествозн. и маркс., и 2.
Филипченко Ю. А. — 1916 — О черепах некоторых видовых гибридов между дикими и домашними формами, Арх. вет. наук.
Якимов В. Л. — 1932 — Иммунен ли зебу, Пробл. животнов., № 11–12.
Глава III
BerIioz — 1927 — Bull. Soc. Zool. Françe, 52, p. 396.
Boulineau P. — 1932 — Un croisemerit antilope-vache, La Nature 15, XII, № 2895.
Brehm A. — 1831 — Handb. Vögel Deutschlands, S. 504.
Fritsch — 1887 — Mt. orn. Ver., Wien, S. 127.
Germar E. F. — 1821 — Begattung verschiedenartigen Insekten untereinander, Mag. der Entomol., Bd. IV, 404 — 409.
Gistl — 1827 — Isis, p. 625.
Gloger C. W. L. — 1853 — Die Neigung der Enten-Arten zur Vermischung dureh Begattung miteinander, J. Ornith., 409-418.
Gloger C. W. L. — 1854. — Das Verbastardieren der Waldhühner (der Gattung Tetrao Lin), J. Ornith. 129–136.
Hagedoorn H. L. — 1901 — On the purely motherly character of the hybrids produced from the eggs of Strongylocentrotus, Arch. Entw. Mech., Bd. 27.
(
Hagenbeck und van Aakeen) — 1897 — Заметка в Vom Fels zum Meer, Jahrs. XVI, H. 26, Stuttgart.
Hertwig G. — 1918 — Kreuzungsversuche am Amphibien I. Wahre und falsche Bastarden., A. micr. Anat., 91., Abt. II, 203.
Heriwig G. — 1913 — Partenogenese bei Wirbcltieren hervorgerufen durch artfremden radiumbestrahlten Samen, A., micr. Anat., 87.
Hertwig P. — 1916 — Durch Radiumbestrahlung verursachte Entwicklung von halbkernigen Triton und Froschenembryonen, A. micr. Anat., 87.
Loeb J. — 1908 — Über die Natur der Bastardlarve zwischen dem Echinodermenei (Strongylocentrotus lividus) und Molluskcnsamen (Chlorostoma funebrale), Arch. f. Entwmech., 26.
Lönnberg, Einar — 1905 — On hybrid hares between Lepus timidus and Lepus europaeus Pall. from southern Sweden, Proc. Zool. Soc., London, 278.
Lönnberg E. — 1929 — A Hybrid between Grey Seal Holichoerus gryphus, Nils and Baltic Ringer Seal, Phoca hispida annellata, Nils — Arkiv för Zoologi, 21, 1–8.
Mann C. — 1867 — (Bastardgemsen), Zoolog. Gart., VIII, 275,
Megušar F. — 1907 — Versuchsresultate über die Bastardierung zwischen Hydrophilus piceus L. unt H. atterrimus. Eschtz, Zbl. f. Physiolog, XXI; Verh. Morph. phys. Gesell., Wien, II, Juni.
Meise W. — 1928 — Die Verbreitungder Aaskrähe (Formenkreis Corvus corone L.), J. Ornith., LXXVI, S. 1-203.
Mory E. — 1901 — Über einige neue schweizer Bastarde der Sphingiden Gen. Deilephila etc., Mitteil. Schweiz, ent. Ges. Bd. X, S. 333–360 u.Anmerkungen.
Mory E. — 1901 — Revision der bis jetzt bekannten von hybr. epilobii B. abgeleiteten Bastarde, Mitteil. Schweiz, ent. Ges., Bd. X.
Natliusius H. v. — 1876 — Über die sogenannten Leporiden, Berlin, Wiegandt und Parey.
NegeIein v. — 1853 — (без заглавия), Naumannia, Arch. f. Orn., 223–225.
Neumoegen B. — 1894 — Notes in a remarcable "interfaunal» Hybrid of Smerinthus, Entom. News, Philadelphia, v. 326.
Ogilvie-Grant — 1893 — Catalogue of the Game Birds in the collection of the British Museum, Cat. of Birds, XXII.
Pflüger — 1882 — Die Bastardzeugung bei den Batrachiern, Pfl. Arch., 29.
Poll H. — 1908 — Mischlingsstudien. Ill, System und Kreuzung, № 6, 127–139.
Poll H. — 1910 — Über Vogelmischling, Ber. über den V. Internal. Orn. Congress, Berlin, 399–468.
Przibram H. — 1907 — Paarungsversuche an Gottesanbeterinncn, Verh. Morph. phys. Ges. Phys. Zbl., 21.
Przibram H. — 1910 — Experimental Zoologie, Leipz. und Wien, 3 T.
Rothschild W. u.
Hartert E. — 1903 — Notes on Papuan Birds, Novit. Zool., 65-116.
Rothschild W. — 1930 — Notes on the preceding article of Dr. Stresemann, Nov. Zool., XXXVI, 16–17.
Rothshild Lord. — 1931 — Notes on Paradisaeidae with a List of the Species, subspecies and Hybrids exhibited at the seventh Int. Orn. Congress, Proc. VII Int. Orn. Congr., 285–294.
Sclater P. L. and
Thomas O. — 1894–1900 — The book of antelops, London, Portir.
Sharpe R. B. — 1899–1909 — A Hand List of the Genera and Species of Birds, v. I–V, London.
Stegmann B. — 1928 — Zur Svstematik der Rotschwänze. J. Ornith. LXXVI, H. 3., 496–502.
Stresemann E. — 1931 — Welche Paradiesvogelarten der Literatur sind hybriden Ursprungs, Proc. VII Int. Orn. Congress, Amsterdam, 284 (автореферат).
Suchetet A. — 1890 — Les oiseaux hybrids rencontrés á l’état sauvage.
Sushkin P. P. — 1929 — On two groups of hybrids in Shrikes and Thrusher. Verh. VI. Intern. Orn. Congress, 382–386.
Villiers de — 1832 — Note sur un accouplement de deux Zygènes d’espfèces differentes, Ann. Soc. Ent. Françe, I, 421.
Walterstorff W. — 1903 — Zur Frage der Bastardnatur des Triton. Blasii de l’lsle, Zool. Anz., XXVI, 692.
Walterstorff W. — 1903 — Über Triton Blasii de l’lsle und den experi-mentellen Nachweis seiner Bastardnatur., Zool. Jahrbuch., Abt. f. Syst., XIX, 647–661.
Walterstorff W. — 1904 — Triton blasii de l’lsle ein Kreuzungsproduct zwischen Triton marmoratus und T. cristatus, Zool. Anz., XXVIII, 82.
Берг Л. С. — 1912–1914 — Рыбы. Фауна России, т. III, вып. 1 и 2.
Богданов А. П. — 1883 — Медицинская зоология, I, стр. 664–687.
Завадовский М. М. — 1931 — Бесплодие самцов, плодовитость самок и материалы по генетике гибридов зебу-як, Тр. по динам. разв., VI, 221–240.
Корнилов И. П. — 1856 — Олени Восточной Сибири, Вест. естеств. наук.
Мензбир М. А. — 1904–1909 — Птицы.
Огнев С. И. — 1931 — Звери Восточной Европы и Северной Азии, т. II, Госизд.
Глава IV
Beebe Will. — 1918–1922 — A monograph of the pheasants, v. I–IV, London, Witherby.
Bemmelen J. F. — 1909 — Über den Unterschied zwischen Hasen und Kaninchenschadeln, Leiden, S. 134.
Friedel E. — 1880 — Die lebenden Wassertiere auf den Intern. Fischerei ausst. zu Berlin im Jahre 1880, Zool. Gart., 333.
Friedenthal H. — 1911 — Tierhaaratlas 989 Abb., Fischer, Jena,
Fiirbringer M. — 1888 — Untersuchungen zur Morphologie und Systematik d. Vögel, Amsterdam.
Ghigi A. — 1915 — Ricerche sull’incrocimento del Gallus sonnerati con polli domestici, Mem. R. Accad. Science, Bologna, Ser. VII, t. 3.
Girtanner — 1880 — Die Pflege der Gemse in der Gefangenschaft, Zool. Garten, XXI, 44.
Glock — 1914 — Rassenverwandschaft und Eiweissdifferenzierung, Biol. Zbl Bd. 34.
Griffini A. — 1913 — Alc. consid. sulle Zebre e sui loro ibridi, Rivista mensila di Sc. Nat., «Natura», v. IV.
Knauthe K. — 1896 — Zool. Gart., № 7, 220.
Kraus K. und
Przibram H. — 1907 — Zur biologischen Differenzierung von Bastardenmittels der Präzipitinreaktion., Verh. Morph. Physiol. Ges., Phys. ZbI.
Leonhardt E. — 1905 — Schleierschwanz Barbus conchonius und einiges über Bfifruchtungsvorgänge — Natur und Haus, XIII.
Lyon M. W. — 1904 — Classification of the Hares and their Allies, Smithson. Miscell. Collect. XLV, Pub, № 1445, v. I, p. 3 and 4, p. 321, 447, Pl. LXXIV–C, t. fig. 44–45.
Nort de — 1769 — Observations sur une mule qui mit bas un petit muleton bien conformé à Saint-Domingue, Mem. Acad. Sc. Paris, A. Hist, 47.
Oppel Albuand. — 1896–1914 — Lehrbuch der vergleichenden microsko-pischen Anatomie der Wirbeltiere, Fischer, Jena.
Regan C. Tate — 1913 — A revision of the Cyprinodont fishes of the subfamiles Poeciliinae, Proc. Zool. Soc., pp. 977 — 1018,
Rhoads Samuel N. — 1896 — A contribution to the Zoology of Tenessee, Proc. Acad. Nat. Sc., Philadelphia, 175–205.
Rhoads Sam. N. — 1896 — Synopsis of the polar hares of North America, Proc. Ac. Nat. Sc., Philadelphia, p. 359–376, Pl. VI, VII, VIII, IX, X.
Sommerfeld K. — 1927 — Das Madurarind, eine Kombinationszuchtung zweier Rinderarten, des Balirindes und des Vorderindischen Zebus, Z. Tierzucht, 8.
Standfuss M. — 1896 — Handbuch der paläarktischen Grossschmetterlinge für Forscher und Sammler, Jena, Fischer.
Watson J. u.
Wailly A. — 1893 — On the hybrid silk-moths hybridised and bred in North America, Entomologist, XXVI, 173.
Weber M. — 1928 — Die Säugetiere, II. Aufl. G. Fischer, Jena.
Алфераки, Гуси России.
Берг Л. С. — 1923 — Рыбы пресных под России, 2-е изд., Госиздат.
Скориков А. С. — 1929 — Новое основание для ревизии рода Apis L., Изв. по прикл. энтомол., т. IV, стр. 249–270.
Тихомиров А. — 1914 — Основы практического шелководства, Москва.
Филипченко Ю. А. — 1928 — Частная генетика, ч. II, Животные.
Шарп (Кузцецов) — 1902–1910 — Насекомые. Библ. естеств. Брокгауз-Ефрон.
Глава V
Anderson, Doroty — 1927. — Am. J. of. Physiol.
Castle W. E. and
Phillips I. C. — 1911 — On germinal transplantation. in Vertebrates, Carn. Inst. Publ., 144.
Domm L. V. — 1930 — Artificial insemination with motile sperm from ovariotomised fowl., Proc. Soc. Exp. Biol. Med., 28(3) 316–318 (Biol. Abst., 1932, II, 3889),
Doubleday H. — 1842. — Entomologist, Abs., p. 357.
Heinroth O. — 1906 — Beobachtungen an Entenmischlingen, Sitzber. d. Ges. nat. Freunde, № 1, S. 3.
Heinroth O. — 1910 — Beobachtungen bei einem Einburgerungsversuch mit der Brautente (Lampronessa sponsa), J. Ornith, LVIII, 101–156, Taf. 1–4.
Heinroth O. und
M. — 1928 — Die Vögel Mitteleuropas, I — III.
Heyer Emil — 1898/99 — Hybridisation zwischen Bombyciden, Soc. Ent., XIII, 137–138.
Hockery — 1870 — Über die Bastardierung der Vögel, J. für Ornith., Bd. 18, 3. Folge, Bd. I, S. 152.
Küpfer — 1920–1923 — Beiträge zur Morphologie der weiblichen Geschlechsorgane bei den Säugetieren. Vierteljahrschr. naturf. Ges., Ziirich, Bd. 65 u. 68.
MuzaIewsky B. M. und
KozIow D. N. — 1933 — Unsere Erfolge in der künstlichen Begattung der Bienenköniginnen, Arch. Bienenkunde, H. 4.
Nicolas — 1774 — Indicàtion sur la maniOre d’élever les papillons, les précautions qu’il faut prendre á Regard des Chrysalides et sur la methode de se procurer des Métis en ce genre — Roziers. J. de Physique, t. IV, p. 482.
Owen — 1861 — Essays and observations by I. Hunter.
Poll H. — 1909 — Mischlinge von Triton cristatus Laur und Triton vulgaris L., Biol. Zbl., XXIX, 30.
Spallanzani — 1768 — Invito a intraprendere sperienze ondre averre muleti nel popolo degl’insetti per tentar di scogliere il gran problema della genera-zione.
Standfuss M. — 1884 — Lepidopterologisches. Zwei neue Hybridcn, Stet-tiner Ent. Zeitung, XLV, 195.
Sturtevant A. H. — 1929 — The Genetics of Drosophila simulans. Publ. Carn. Inst.
Yamane I. and
Egashira T. — 1925 — Über Kreuzungsversuche zwischen Kaninchen (Lepus cuniculus) und Schneehasen (Lepus timidus ainu) durch natürliche Paarung und künstliche Befruchtung, Zschr. Tierzucht.
Yochem D. — 1929 — Spermatozoon life in the female reproductive tract of the Guinea piffand Rat., Biol. Bull., 56.
Wolterstorff W. — 1909 — Über Polls Bastarde zwischen Triton cristatus Laur und T. vulgaris., Zool. Anz., XXXIII, 850.
Бесхлебнов А. и
Лазарев Г. — 1934 — Минеральные разбавители в искусственном осеменении, Пр. жив., № 5, 128–129.
Дарвин Ч., Происхождение человека, ч. 2, гл. XVIII, сноска 33.
Кардымович М., Милованов В. — 1933 — Искусственное осеменение кролика, Пробл. животнов., № 1.
Кузнецова Н., Милованов В., Нечаев В., Нейман О., Скаткин П. — 1932. — Искусственное осеменение рогатого скота, Сельхозгиз.
Кузьмина М. Д. — 1933 — Искусственное осеменение уток, Тр. инст. с. х. гибрид. и аикл. жив. т. I, стр. 197–205.
Милованов В. К. — 1933 — Итоги трехлетней работы по разбавителям для спермы с.-х. животных, Пробл. животнов., № 4.
Милованов В. и Селиванова С. — 1932 — Разбавители для спермы с.-х. животных, Пробл. животнов., № 2.
Милованов В., Хабибулин X. — 1933 — Анабиоз сперматозоидов и его использование в соц. животноводстве, Пробл. животнов., № 5.
Морган Л. — 1899 — Привычка и инстинкт, русск. изд. Павленкова.
Музалевский Е. М. и
Козлов Д. Н. — 1933 — Искусственное осеменение пчелиных маток, Пробл. животнов., № 6, стр. 92–97.
Никитина М. В. — 1932 — Проблема искусственного осеменения кур, Пробл. животнов., № 9-10, стр. 97-100.
Осипов М. Ф. — 1933 — Техника иск. осемен. при межвидовой гибридизации, Тр. Инст. с. х. гибридиз. и аккл. жив., т. I, 188–196.
Румянцев Б. Ф. — 1934 — Видовая гибридизация как метод создании новых пород дом. животных, Пр. жив. № 5, 60–64.
Серебровский А. С. и
Соколовская И. И. — 1934 — Электроэякуляцня у птиц, Пр. жив., № 5, 57–59.
Тиняков Г. Г. — 1933 — Гибрид от скрещивания курицы с павлином, Пробл. животнов, № 1, стр. 82–86.
Тиняков Г. Г. — 1933 — Гибрид. от скрещивания павлина с курицей, Тр. Инст. гибрид., т. I, стр. 85–98.
Тиняков Г. Г. — 1933 — Новый метод получения спермы у куриных, Пробл. животнов., № 6, 90–92.
Фортунатов Б. К. — 1928 — Зоопарк. Сборн. «Степной заповедник Аскания-Нова», стр. 50-108.
Щорохова А. А. — 1933 — Спермии человека и т. д. Журн. УзНКЗдрава, № 11–12, стр. 53–82, Ташкент.
Штандфусс М. — 1901 — Жизнь бабочек, под. ред. Шевырева, изд. «Просвещение».
Глава VI
Agassiz — 1874 — Note sur la fertilisation artificielle de deux espêces d’étoiles de mer. Arch. Zool. exp., III Notes et revues, p. XLVI.
Baltzer T. — 1910 — Über die Beziehung zwischen dem Chromatin und der Entwicklung und Vererbungserscheinungen bei Echinodermenbastarden, Arch, f. Zellforsch., 5.
Born G. — 1883 — Beiträge zur Bastardierung zwischen den einheimischen Anureenarten, Pfl. Arch., 32.
Boveri Th. — 1888 — Über partielle Befruchtung, Sb. Ges. Morph., München, 4.,
Boveri Th. — 1899 — Ein geschlechtlich erzeugtes Organismus ohne mutter diche Eigenschaften, Ber. d. Ges. f. Morph, u. Phys., München.
Boveri Th. — 1895 — Über die Befruchtungs und Entwickelungsfahigkeit "kemloser Seeigeleier und über die Moglichkeit ihrer Bastardierung, A. Entw. mech., 2.
Boveri Th. — 1900 — Über den Einfluss der Samenzelle auf die Larvenchara-ktere der Echiniden, Arch. Entw. mech., 16.
Boveri Th. — 1904 — Noch ein Wort über die Seeigelbastarde, Arch. Entw. mech, 17.
Brachet A. — 1913 — Recherches sur le determinisme héréditaire de l’oeuf des mammiffêres. Developpement in vitro de jeunes vésicules blastqdermiques de lapin, Arch, de Biol., XXVIII.
Driesch H. — 1903 — Über Seeigelbastarde, Arch. Entw. mech., 16.
Driesch H. — 1892 — Über rein mutterliche Charaktere an Bastardlarven von Echinodermen, Arch. Entw. mech., 7.
Godlevsky E. — 1905 — Die Hybridisation der Echiniden und Crinoideen-faitiilien, Bull. Acad, de Sc., Cracovie.
Godlevsky E. — 1906 — Untersuchungen über die Bastardierung der Echiniden und Crinoidenfamilie, Arch. Entw. mech., 20.
Godlevsky E. — 1909 — Das Vererbungsproblem im Lichte der Entwicklungsmechanik betrachtet, Roux’s Vortr., Leipzig.
Godlevsky E. jun. — 1912 — Studien über Entwicklungserregung. Arch. Entw. mech., 33.
Goldschmidt R. — 1915 — Some experiments on spermatogenesis in vitro, Proc. Nat. Ac. Sc., p. 220.
Goldschmidt R. — 1916 — Notiz über einige bemerkenswerte Erscheinungen in Gewebekulturen von Insekten, Biol. Zbl., XXXVI, 160. "
Goldschmidt R. — 1917 — Versuche zur Spermatogenese in vitro, Arch. Zellforsch., XIV.
Gruzdeff W.. S. — 1896 — Versuche über die künstliche Befruchtung von Kanincheneiern, Arch. f. Anat. u. Phys.,-S. 269–304.
Herbst C. — 1905 — Vererbungstudien, I–III, Arch. Entw. mech., 21.,
Herbst G. — 1906 — Vererbungstudien, IV, Arch. Entw. mech., 22.
Hertwig O. u.
R. — 1887 — Über den Befruchtungs- und Teilungsforgang des tierischen Eies, Jena.
Kupelwieser H. — 1906 — Versuche über Entwickelungserregung und Membranbildung bei Seeigeleiern durch Molluskensperma, Biol. Zbl., S. 744–798.
Kupelwieser H. — 1912 — Weitere Untersuchungen über Entwiklungserregungen durch stammfremde Spermien, Arch. Zellforsch., 8.:
Levis M. R. and
Robertson R. — 1916 — The Mitochondria and other Structures observed by the Tissue Culture Methode in the Male Germ. Cells of Chorthippus curtipennis Scudd., Biol. Bull., XXX.
Loeb J. — 1903 — Über die Befruchtung von Seeigeleiern durch Seesternsamen, Pfl. Arch.
Loeb J. — 1903 — On a method by which the Eggs of a Sea-Urchin can be fertilised with the Sperm of a Starfisch., Univ. of Calif. Public. Physiol., I. 1.
Loeb J. — Weitere Versuche über die heterogene Hybridisation bei Echinodermen, Pfl. Arch., Bd. 104.
Morgan T. H. — 1895 — Studies of the «partial» larves of Sphaerechinus, Arch. Entw. mech., 2.
Morgan T. H. — 1895 — The fertilisation of non-nucleated fragments of Echinodfirmeggs, Arch. Entw. mech., 2.
Pfeffer — 1881–1885 — Untersuchungen aus dem botanischen Institut zu Tübingen, I, 363.
Pincus G. — 1931 — Observations on the living eggs of the Rabbits, Proc. Roy. Soc., Ser. B., v. 197, N. 749, 132–168.
Seeliger O. — 1894 — Gibt es geschlechtlich erzeugte Organismus ohne mutterliche Eigenschaften? Arch. Entw. mech., 1.
Seeliger — 1895 — Bemerkungen über Bastardlarven der Seeigel., Arch. Entw. mech., 3.
Teichmann E. — 1903 — Über Furchung befruchtcter Seeigeleier ohne Beteiligung der Spermakerncn, Jen. Ztschr. d. Naturwiss., 37.
Tennent D. H. — 1909 — Dep. of Marine Biology of the Carn. Instit. Annual Rep. of Director.
Vernon H. M. — 1909 — Cross-fertilisation among Echinoides. Arch. Entw. mech., 9.
Vernon H. M. — 1900 — The relation between the Hybrid or parent forms of Echinoderm larvae, Phil. Tr. R. S., v. 13, p. 190.
Wolff E. K. und
Zondek B. — 1925 — Die Kultur menschlichen Ovarial und Amniongewebe, Virchow’s Arch., Bd. CCLIV, 1.
Вермель E. M. — 1923 — Культивирование вне организма ткани семенника петуха. Тр. Инст. с.-х. гибридизации и акклим. животных, т. I, стр. 155–167.
Красовская О. В. и
Диомидова Н. А. — 1934 — Оплодотворение яиц кролика in vitro, Биолог. журн., т. III, стр. 10–19.
Леб Ж. — 1926 — Организм как целое, Совр. пробл. естествозн., кн. 33, Гиз.
Лилли Ф. Р. — 1925 — Проблемы оплодотворения, Совр. пробл. естествозн., кн. 29, Гиз.
Глава VII
Baltzer F. — 1930 — Über die Entwicklung des Tritonmerogons, Rev. Suisse. Zool., 37.
Bytinski-Salz — 1933 — Lepidopterenhybriden II, Arch. Entw. mech., 129.
Clement — 1923 — Essai d’hybridisation entre une chienne et un renard, C. R. Soc. Biol., v, p: 1300.
Gordon, Myron — 1931 — Hereditary base of melanosis in hybrid fishes, Amer. J. Cancer, v. XV.
Guyer M. F. — 1909 — On the sex of hybrid birds, Biol. Bull., v, 16, 4, 193–198.
Hadorn E. — 1932 — Über Organentwicklung und histologische Differenzieruug in transplantierten merogonischen Bastardgeweben, Arch. Entw. mech., 125, 495–565.
Haldane J. B. S. — 1922 — Sex ratio and unisexual sterility in hybrid animals, J. Genetics, XII.
Hanssler G. — 1928 — Über Melanombildungen bei Bastarden von Mannchen var. pulchra, Bl. f. Aquar.
Hanssler G. — 1928 — Über Melanombildungen bei Bastarden von Xiphophorus helleri und Platypoecilus maculatus var. rubra, Klin. Wschr., 7, 1561–1562.
Kosswig, C. — 1928 — Über Krcuzungen zwischen den Teleostieren Xiphophorus helleri und Platypoecilus maculatus. II. Ztschr. ind. Abst. Vererb, Bd. 47.
Kosswig C. — 1929 — Über die veränderte Wirkung von Farbgonen des Platypoecilus in der Gattungskreuzung mit Xiphophorus. Ztschr. ind. Abst. Vererb., Bd. 50, 63.
Kosswig C. — 1929 — Melanotische Geschwulstbildungen bei Fischbastarden, Verh. Dcutsch. Zool. Ges., 90–98.
Kosswig C. — 1929 — Zur Frage der Geschwulstbildung bei Gattungsbastarden der Zahncarpen-Xiphophorus und Platypoecilus, Ztschr. ind. Abst. Vererb, Bd. 52, S. 114–120.
Reed H. D. and
Gordon M. — 1931 — The morphology of melanotic overgrowths in Hybrids of Mexican Killifisches, Amer. J. Cancer., v. XV.
Schultz W. — 1912/13. — Bastardierung und Transplantation I, II, III, Arch. Ent. mech., Bd. 35, S. 484, 1912; Bd. 36, S. 353; Bd. 37, S. 265, 1913.
Schultz W. — 1917 — Gleichlauf von Verpflanzung und Kreuzung Froschlar-ven, Arch. Ent. mech., Bd. 43, S. 363.
Schultz W. — 1913 — Vorschläge zur Studien der somatischen Vererbung der Bastardfruchtbarkeit u. s. w., Arch. Ent. mech., Bd. 37, S. 285.
Standfuss M. — 1930 — Successful rearing of Amorphc populi ♂ and Smerintus ocellata ♀, Ent. Rec., 14, 191.
Федерлей Л. — 1930 — Сублетальные комбинации хромосом, Труды съезда генетиков и т. д., т. II, стр. 509–582.
Глава VIII
Dobzhansky Th. — 1933 — On the sterility of the interracial hybrids in Drosophila pseudoobscura, Proc. Nat. Sci Sc.
Dobzhansky Th. and
Boche R. D. — 1933 — Intcrsterile race of Drosophila pseudoobscura, Biol. Zbl., Bd. 53.
Federley H. — 1913 — Das Verhalten der Chromosomenarten bei der Spermatogenese der Schmetterlinge Pygaera anachoreta, curtula and pigra sowie einiger ihrer Bastarden. Ein Beitrag zur Frage der konstanten intermediaren Artbastarde und der Spermatogenese der Lepidopteren (Taf. 1–4), Z. ind. Abst. Vererb., 1 — 110.
Federley H. — 1916 — Chromosomenstudien an Mischlingen III. Die Spermatogenese des Bastards Choerocampa porcellus×elpenor, Öfversigt af Finska Vetenskaps-Societetens Förhandl., 58 A, № 12, p. 17.
Fraser A. C. and
Gordon, Myron — 1928 — Identity of genes and sex factors in intergeneric killifisch hybrids, Anat. Rec., 41, 198.
Fuller Edw. — 1836 — Notice of the rearing of two hybrids from a Barndoor Hen having a cross from a Pheasant and a Pheasant cock (Tree quarter bred pheasants), Proc. Zool. Soc., IV, 84–85.
Greenwood — A. W. — 1925 — Gonad grafts in embryonic chics and their relation to sexual differentiation, Brit. J. Exp. Biol., 2.
Guyer M. — 1930 — Spermatogenesis of normal and hybrid pigeons, A dissertation, Chicago.
Guyer M. — 1912 — Modifications in the testes of hybrids from the guinea and the common fowl, J. of Morphol., v. 23, p. 45–65.
Iwanoff E. — 1905 — Untersuchungen über die Ursachen der Unfruchtbarkeit von Zebroiden, Biol. Zbl.. Bd. 25, S. 789–804.
Müller R. — 1908 — Das Problem der sekundaren Geschlechtsmerkmale und die Tierzucht, Stuttgart, Ferd. Ekk., 89, S.
Meisenheimer J. — 1924 — Die Vererbung von Art und Geschlechtsmerkma-len bei Biston-Artkreuzungen, Zool. Jahrb., Abt. f. allg. Zool. und Phys. d. Tiere, Bd. 41, S. 51–90.
Pariser K. — 1927 — Zschr. f. Zellf. u. Micr. Anat., 5.
Riddle O. — 1916 — Sex-control and known correlation in pigeons, Am. Nat., 50; 385–410:
Riddle O., Smith G. C.. a.
Benedict F. G. — 1932 — The basal metabolism of the mourning dove and some of its hybrids, J. Amer. Physiol., 101.
Poll H. — 1906 — Mischlingstudien I. Der Geschlechtsapparat der Mischlinge von Cairina ♂ moschata und Anas boschas var. dom ♀, Sitzb. Ges. nat. Freunde, № 1, 4–7.
Poll H. — 1908 — Demonstration auf der 22 Versammlung der Anatomischen Gesellschaft zu Berlin, Erg. Heft zum Anat. Anz., Bd. 32, S. 304.
Poll H. — 1910 — Mischlingsstudien IV. Keimzellbildung bei Mischlingen. Verh. der Anat. Gesellsch. 2 Int. Kongr. in Briissel., Ergänzungsheft zum Anat. Anz., Bd. 37, S. 32–57.
Poll H. — 1911 — Misehlingsstudien. V. Vorsamenbildung bei Mischlingen, Arch. micr. Anat., Bd. 77, Abt. 2, S. 210–239.
Poll H. — 1911 — Misehlingsstudien. VI. Eierstock und Ei bei fruchtbaren und unfruchtbaren Mischlingen, Arch. micr. Anat., Bd. 78, Abt. 2. S. 63 — 127.
Poll H. und
Tietensee W. — 1907 — Misehlingsstudien. II. Die Histologie der Keimdrüsen bei Mischlingen, Ztschr: Ges. nat. Freunde, № 6, 157–166.
Smith, Geoffrey — 1912 — Studies in the experimental analysis of sex. Part 9. On spermatog. and the formation, of giant spermatozoa in hybrid pigeons, Quart. J. Micr. Sci., v. 58, part I, 159–170.
Schultz J. and
Dobzhansky Th. — 1933 — Triploid Hybrids between Drosophila melanogaster and Drosophila simulans, J. exp. Zool., v. 65.
Thomas R. H. and
Huxley J. S. — 1927 — Sex-Ratio in Pheasant, species-crosses, J. Genet., XVIII, p. 233–246.
Turner C. L. — 1929 — The gonads of some Platypoecilus — Xyphophorus hybrids, Anat. Rec., 44, 243.
Willier B. H. — 1927 — The specifity of sex, of organization and of differentiation of embryonic chics gonads as shown by grafting experiments, J. Exp. Zool., 46.
Witschi E. — 1927 — Sex reversal in parabiotic twins of the Rana silvatica, Biol. Bull., 52.
Witschi E. — 1927 — Testes grafting in tadpoles of Rana temporaria and its-bearing on the hormon theory of sex determination, J. Exp. Zool., 47.
Whitman Ch. O. — 1919 — Posthumous Works, v. II, Carnegy Inst.
Керкис, Ю. Я. — 1935 — Гибридизация Dr. melanogaster и Dr. simulans в связи с вопросом о причинах стерильности и т. д. — Докл. Ак. Наук СССР № 1.
Шапиро Н. И. — 1934 — Межвидовая гибридизация внутри рода Drosophila — Зоол. журнал, XII. 153-175
Глава IX
Adlersparre A. — 1918 — Poephila gouldiae E Poephila mirabilis En. korsing mellan australiska väfvarfaglar, Fauna och Flora.
Biddle R. L. — 1932 — The bristles of Hybrids between Drosophila melanogaster and Drosophila simulans, Genetics, 17, p. 153–174.
Bonchote J. L. — 1915 — Vigour and Heredity, London, West Newnan & Co.
Denso P. — 1908 — Die Ersclieinnng der Antizipation in der ontogenet. Entwicklung hybrider Schmetterlingsraupen., Zschr. wissensch. Insektenbiol., IV, S. 123–125, 201–208.
Dunker H. — 1931 — Erblichkeitsverhaltnisse bei Vögeln, Proc. VII. Int. Orn. Congress, Amsterdam, 1930.
Elsasser Th. — 1925 — Die Struktur schellender Federn, J. ornith. 73, 337–389.
Federley H. — 1915 — Finska Veteriscaps. Soc. Forhandlinger, 57.
Federley H. — 1922 — Über einen Fall von Criss-Cross-Vererbung bei einer Artkreuzung, Hereditas, Bd. 3, 125–146.
Federley H. — 1923 — Bilden Chromosomenkonjugation Mendelspaltung und Fertilitat bei Speziesbastarden einen Dreibund? Hereditas, 4, 161–170.
Federley H. — 1927 — Ist die Chromosomenkonjugation eine conditio sinequa non für die Mendelspaltung, Hereditas, 9, 391–404.
Fischel A. — 1906 — Über Bastardierungsversuche bei Echinodermen, Arch. Entwick. mech., 22.
Foot and
Strobell — 1914 — The Chromosomes of Euschistus variolarius E. servus and the Hybrids of the F
2 anf Generationen, Arch. Zellforsch., 12, S. 485.
Clerschler M. W. — 1914 — Über alternative Vererbung bei Kreuzung von Cyprinodontiden-Gattungen — Ztschr. und. Abst. Vererb. Bd. XII, s. 73.
Ghigi A. — 1900 — Di nu ibrido fra Numida e pavone. Monit. Zool. ital. XII Anno suppl. Rend della la Assembl. ord e del Conv. dell’ Univ. Zool. ital., Bologna, 16–17.
Ghigi A. — 1907 — Contribute alio studio dell’ibridismo negli ucelli, Atti della-R. Acc. dei Lincei, Anno CCIV ser V Renadic., v. XVI, Fasc. 9, 791–800.
Ghigi A. — 1908 — Silla dissociazione dei caratteri specified negli ibridi comp-lessi di alcuni ucelli. Atti della R. Ace. dei Lincei Anno CCIV, ser. V. Rendic, v. XVII, 452–461.
Guyer M. — 1909 — Atavism in guinea-chiken hybrids, J. exp. Zool., v. 7, 723–745.
Guyer M. — 1909 — La livrée du plumage chez les hybrids de pintade et de poule. Bull, du Mus. d’hist. natur., № 1, p. 3–5.
Hagedoorn A. L. a.
A. C. — 1922 — Species cross in Rats, Z. ind. Abst. Vererb., XXIX.
Hertwig G. u.
P. — 1914 — Kreuzungsversuche an Knochenfischen A. Micr. Anat., B. 84, S. 49.
Kammerer P. — 1907 — Bastardierung von Flussbarch (Perea fluviatilis L.) und Kaulbarch (Acerina cernua L.), Arch. Entw. mech., XXIII.
Kammerer P. — 1907 — Demonstration neuer Tierbastarde. Verh. Morph.. Phys. Gesellsch., Wien, Zbl. f. Physiol., XXI.
Kirk P. — 1890 — Hybrids between Smerinthus ocellatus a. S. populi, Ent. Records, I, 95, 202 (The Enthomologists Records and Journal of Variation, London, E. Knight).
Kleiner E. — 1913 — Untersuchungen am Gcnitalapparat von Helix nemoralis, hortensis und einer weiteren Reihe von lang gezüchteter Bastarde der beiden Arten, Taf. 7 — 10, Ztschr. ind. Abst. Vererb., IX, 216–262.
Lang A. — 1908 — Über die Bastarde von Helix hortensis Muller und Helix nemoralis L. (Mit Beitragen von Bosshard, Ilesse, und Kleiner). Univ. Jena, Fischer, 114 S., 4 Taf.
Loeb, King and
Moore — 1910 — Über Dominanzerscheinungen bei den Hybrideen Pluteen und Sceigeln. Arch. Entw. mech., 29.
Moenkhaus W. J. — 1904 — The chromatin in the developement of hybrids. Amer. J. Anat., v. 3.
Moenkhaus W. J. — 1904 — The developement of Hybrids between Fundulus heteroclitus and Menidia notata etc., Amer. J. Anatomy, v. 3.
Moenkhaus M. J. — 1906 — Fundulus hybrids Rep. Stat. Exp. Evolut., 103.
Morris M. — 1914 — Behaviour of chromatin in Hybrids, J. exp. Zool., 16, 501–511.
Morton S. G. — 1847 — Description of two living hybrid Fowls between Gallus. and Numida, Ann. mag. nat. hist., XIX, 210.
Nabours R. K., Larson J. and
Hartwig — 1933 — Inheritance of Color patterns in the Grouse Locust Acrydium arenosum, Genetics, 13, p. 159.
Naumann J. F. — 1822 — Naturgeschichte der Vögel Deutschlands, 11, 63.
Newmann H. H. — 1908 — The process of heredity as exhibitet by the develop. of Fundulus hybrids, J. exp. Zool., V, 503; — 562.
Newmann H. H. — 1910 — Further studies as the process of heredity in Fundulus hybrids, J. exp. Zool., VIII, 143–163.
Newmann H. H. — 1916 — J.. of exp. Zool., v. 18.
Noack Th. — 1908 — Über den Schädel eines Bastards von Tiger ♀ und Love ♂ Zool. Anz., XXXVIII, 677, 9 Fig.
Phillips Y. C. — 1913 — Reciprocal Crosses Reeves’s Phaesant and the Common Ring-Neck Pheasant producing unlike Hybrids. Amer. Nat., XVII, p. 701–704.
Pierce F. N. — 1906 — Notes on Hybrid Notodonta ziczac × dromedarius, Enthomologist, XXXIX, 122–124.
Poll H. — 1912 — Misehlingsstudien, VII. Mischlinge von Phasianus und Gallus. Sitzber. Kgl. Preuss. Acad. Wissensch., 38, S. 864–883.
Robertson W. R. B. — 1931 — Chromosome studies. III. Inequalities and deficicnces in homologous chromosomes. Their bearing upon synapsis and the loss of unit characters, J. Morph., 26, 109–141.
Rotschild W. — 1911 — On recently described Paradiseidae with notes on some other new species, Ibis, 350–367.
Serebrovsky A. S. — 1929 — Observation on interspecific Hybrids of the fowl, J. Genet., v. XXI.
Standfuss M. — 1898 — Experimented Zool. Studien mit Lepidopteren. Neue Denkschriften der allgem. Schweiz. Gesellschaft f. d. ges. Nat. Wiss.
Sumner F. B. — 1932 — Genetic, distributional and evolutionary studies of the subspecies of the deer mice (Peromyscus), Bibliogr. Genetica, IX.
Steinbruck H. — 1902 — Über die Bastardbildung bei Strongylocentrotus lividus und Sphaerachinus granularis, Arch, Entw. mech., 14.
Tennent D. H. — 1910 — The dominance of maternal or of paternal characters in Echinoderm. Hybrids, Arch. Entw. mech., 29.
Zawadowsky M. M. — 1931 — Zebu-Yak Hybrids, J. Heredity, XXII, 297–313, 17 Fig.
Гиц С. — 1933 — Гибриды Между яками и швпцами, Пробл. жив. № 5, 59–60.
Власов П., Гершензон С. и
Поляков А. — 1932 — Гибридизация между яком и крупным рогатым скотом, Пробл. животнов., № 1, стр. 48–57.
Промитов А. Н. — 1928 — Гибридизация вьюрков, Журн. эксп. биол. Сер. А, т. IV, стр. 30–64.
Соколовская И. И. — 1934 — Опыты по межвидовой гибридизации (в печати).
Глава X
Adametz L. — 1934 — über die Dicke der Muskelfasem als angebliches Mass der Konstitution, Zschr. f. Tierzucht. biol., Bd. 1.
Donaldson — 1909 — On the relation of the Body Length to the Body Weight of the Brain and the Spinal cord in Albino Rat (Mus. Norvegicus var. albus), J. comp. Neurol. Physiol., XIX, 155–167.
Hesse R. — 1921 — Das Herzgcwicht der Wirbeltiere, Zool. Jahrb., Abt. allg. Zool. u. Phys., XXXVIII, 243.
Hesse R. — 1924 — Tiergcographie auf ökologischer Grundlage, Jena, Fischer. —
Klatt B. — 1913 — Über den Einfluss der Gesamtgrösse des Schädelbild., Arch. Ent. mech., Bd. 36.
Parrot C. — 1893 — Über die Grossenverhältnisse des Herzens der Vögel, Zool. Jahrb. VII, Abt. Syst., 496.
Przibram H. — 1922 — Form und Formel im Tierreiche. Beiträge zu einer quantitativen Biologie. I–XX, Leipzig und Wien. F. Deuticke. —
Riboisière J. de la — 1910 — Recherches organometriques en fonction du regime alimentaire sur les oiseaux. Collection de morphologie dynamique, Paris, Herman et fils.
Tieman Otto. — 1919 — Verglcichende Untersuchungen an Haus und Wildenteneiren., Zool. Jahrb., Abt. Zool. u. Phys., XXXVI, 621.
Welker H. u.
Brand A. — 1903 — Gewichtswerte der Körperorgane bei dem Menschen und den Tieren, Arch. Antrop., XXVIIl.
Гессе Р. — 1913 — Тело животного как самостоятельный организм, изд. Девриена.
Примечания
1
Известный уже грекам (υβρισ) и римлянам (hybrida, ibrida, hybris, ibris).
(обратно)
2
от αγωγη — ведение, руководство.
(обратно)
3
У дрозофилы например наследственная изменчивость очень сильна, а на всем огромном протяжении ареала этого вида нет никаких ясных географических форм (Серебровский, 1929, стр. 70).
(обратно)
4
Предположение об участий третьего вида, зондской бородавчатой свиньи. (Sus verrucosus), не подтверждается.
(обратно)
5
Североамериканский луговой волк или койот.
(обратно)
6
Прекрасный анализ такого распадения вида на местные популяции дал недавно Баур применительно к Antirrhinum — испанскому львиному зеву (Baur, 1932).
(обратно)
7
Наследственный характер «дикости» муфлонов- и «смирности» домашних овец можно видеть например на гибридах муфлон × овца в Аскании-Нова.
(обратно)
8
Насколько эти выводы Баура могут быть перенесены на другие группы видов, в настоящее время сказать еще совершенно невозможно за отсутствием необходимых исследований.
(обратно)
9
Возможно, что основанием этой легенды, как и в случае с бесхвостыми кошками, служит какое-либо наследственное уродство, распространенное среди местного скота, вроде знаменитых телят-бульдогов, появление которых на одной опытной станции ставилось в связь если не с гибридизацией, то с психическим воз действием собаки директора станции на тельных коров. Повидимому с летальным геном у коз связаны сообщения о гибридах коз с серной. Недавно нами получено сообщение о появившихся будто бы в 1925 г. в Западной Сибири гибридов свиньи с кобелем, одетых собачьей шерстью, три из которых даже «лаяли». Возможно, что и здесь дело идет о каком-либо уродстве.
(обратно)
10
В 1930 г. в одном заграничном птицеводном журнале была помещена в серьезном тоне написанная статья с прекрасными фото гибридов утки с курицей, совмещавших типичные утиные голову и ноги с прекрасным петушиным хвостом. Некоторые работники в области птицепромышленности приняли эту статью всерьез, не обратив внимания на то, что она была помещена и первоапрельском номере. Получая иностранные журналы с значительным запозданием, советские граждане не раз становились жертвами подобной мистификации.
(обратно)
11
С другой, стороны, и некоторые новые редкие виды признавались первоначально за гибридов. Таков пример дальневосточной Pseudotadorna (ср.
Бутурлин, Зоол. журн., XI, стр. 196).
(обратно)
12
В Британском музее имеется экземпляр, определенный как продукт обратного скрещивания глухаря с межняком. См. Cat. of Birds, XXII, p. 65, 1899 и Fritsch, p. 127, 1887.
(обратно)
13
Подобное сообщение недавно поступило из области Кеми. Надо думать, что и здесь речь идет о какой-либо наследственной аномалии.
(обратно)
14
Матронлинные «псевдогибриды» получались Пржибрамом (Przibram, 1907) при гибридизации богомолов благодаря партеногенезу, а Лангом у молюсков— благодаря самооплодотворению. Подобные источники ошибок при гибридизации полезно иметь в виду.
(обратно)
15
Здесь важно отметить, что с точки зрения гибридизации обычность или редкость вида не имеет никакого значения кроме технического.
(обратно)
16
Интересно например намеченное Осиповым (1933) различное отношение — спермы тура, зебу и бантенга к разбавителям при искусственном осеменении.
(обратно)
17
Приводя этот пример, мы не входим в критический разбор использованного материала, различного у различный авторов.
(обратно)
18
Об этом прекрасно плавающем зайце см. например Roads, pp. 181–182. 1896.
(обратно)
19
Упомянутая выше легенда о гибридах кролика с кошкой возникла на той почве, что в Москве нередки так называемые куцые кошки, имеющие удлиненные задние, ноги и передвигающиеся прыжками несколько наподобие кроликов. Это уродство вызвано мутацией, наследующейся по законам Менделя, хотя и с не всегда хорошим проявлением, и к гибридизации вообще, а тем более к гибридизации с кроликом никакого отношения не имеет, как не имеют отношения к гибридизации с индюками куры-голошейки, о чем тоже существовало мнение среди птицеводов.
(обратно)
20
Наоборот, в 1899 г. Шарп возвел ряд различий в еще более высокое таксономическое значение, дав такую систему куриных:
Отряд Galliformes. Подотряд 1. Megapodii. Сем. 1. Megapodiidae. Подотряд 2. Crares. Сем. 1. Cracidae. Подотряд 3. Phasiani. Сем. 1. Tetraonidae. Сем. 2. Phasianidae. Сем. 3. Numididae. Сем. 4. Meleagridae. Сем. 5. Odontophoridae.
(обратно)
21
По Лейнису в сем. Cyprinidae около 110 родов и 800 видов, а по Иордану — более 175 родов и почти 2 000 видов.
(обратно)
22
Объем сем. Cyprinidae разными авторами принимается различно. Так, в известной русской сводке Никольского («Гады и рыбы») в сем. Cyprinidae включены род Catostomus, выделяемый теперь в особое семейство, и гольцы, вьюны и шиповки, выделенные также в особое сем. Cobitidae.
(обратно)
23
Лаборатория искусственного осеменения Всесоюзного института животноводства.
(обратно)
24
Нам пришлось наблюдать поведение индюка, увидевшего свое отражение в зеркале. Он вступил с своим изображенном в длительную драку.
(обратно)
25
Селезни домашних уток например начинают ухаживать за самками огорей (Casarca casarca), но, услыхав их своеобразный голос, шарахаются в сторону в состоянии резко выраженного испуга.
(обратно)
26
Нахшпиль на первый взгляд как будто не имеет уже биологического смысла, так как осеменение произошло. Однако он систематически и характерно встречается в специфической форме у многих видов. Возможно поэтому, что биологический смысл нахшпиля достаточно серьезен, но пока не раскрыт. Быть может реакции самки на него должна способствовать продвижению спермы? Но такая реакция, если она есть, могла бы установиться и в качестве ответа на саму садку, топтание, контакт клоак и т. д.
(обратно)
27
Нормальный процесс сваривания у L. sponsa несколько иной, чем у кряквы. Самка ложится вытянувшись на воду и производит впечатление павшей — совершенно неподвижно она остается в атом положения многие минуты. В это время самец проделывает возле нее характерные приветственные «попивании», касается кончиком клюва воды к быстро поднимает голову снова. Обычно это приготовление продолжается очень долго, он отплывает и вновь приплывает к самке, плывет за ней в описанном положении. Наконец происходит спаривание, и момент «повисания» самка обычно издает тихий «Locton», и самец своеобразно спрыгивает с ее спины. Характерного для кряквы «нахшпиля» он
не производит, по отплывает с поднятой головой несколько в сторону (Heinroth, 1910, 128).
(обратно)
28
Весной 1933 г. в Московском зоопарке белощекие казарки загнездились, избрав для этого каменную горку, где на клочке сена самка отложила 4 яйца, одев их пышным слоем пуха. По словам П. А. Мантейфеля пытались гнездиться даже 3 пары, но их прогоняли другие обитатели, и только одной паре, в которой оказался достаточно боевым самец (он сидит недалеко от гнезда на той же каменной горке), удалось отстоять избранное место от других гусей. Этот случай не позволяет сказать, что обычное бесплодие этих казарок является лишь экологическим, так как каменная горка имелась и раньше, но тем не менее недостаток подходящих мест для гнезда и конкуренция с другой птицей привели и в этом году всех казарок кроме одной к бесплодию несмотря очевидно на наличие физиологических предпосылок.
(обратно)
29
Теория разбавления впрочем довольно сложна. У видов, у которых естественная сперма сильно разбавляется секретом добавочных желез (лошадь, хряк), разбавление искусственное способно улучшить сперму. У птиц, наоборот, лучше работать с неразбавленной спермой при помощи микропипеток. Бернштейн настаивает на возможности пользоваться для разбавления одним NaCl.
(обратно)
30
Леб. Организм как целое, стр. 62.
(обратно)
31
Леб. Организм как целое, стр. 59.
(обратно)
32
Впрочем и Леб и Купельвизер отмечают, что у довольно значительного числа самок опыт не удается вовсе — очевидно в связи с какими-то индивидуальными свойствами их яйцеклеток.
(обратно)
33
Этот исключительный случай к сожалению обычно фигурирует в сводках (например
Филипченко, Экспериментальная зоология, рис. 56) при описании опытов Купельвизера.
(обратно)
34
Морские лилий являются классом подтипа Pelmatozoa, морские звезды и морские ежи — классами подтипа Eleutherozoa, Оба подтипа иглокожих существуют раздельно уже в кембрии. Звезды также существуют уже в кембрии; так что не только расхождение лилий от звезд и ежей, но даже расхождение стволов звезд и ежей должно было произойти еще до кембрия.
(обратно)
35
Конечно недоразвитие гамет возможно и при экологическом и физиологическом бесплодии.
(обратно)
36
В Японии имеется много различных по силе рас, и в частности самые сильные. Здесь мы имеем в виду «сильные» японские расы.
(обратно)
37
Термин «стохастические процессы» мы предлагаем взамен данного нами ранее (1927, 1930) термина «автоматические», мало удачного но своей двусмысленности.
(обратно)
38
Мы полагаем, что развиваемая Фишером теория эволюции доминантности не настолько доказана, чтобы помешать говорить о доминантных мутациях вообще, особенно. если мы включим сюда и полудоминантные неоморфы типа Bar (полудоминантная мутации у дрозофилы, вызывающая сужение глаза).
(обратно)
39
Следует впрочем отметить, что а отличие от остальных скрещиваний: в случае Antedon и Ctenolabrus их хромосомы в чистом виде изучены не были.
(обратно)
40
Echinus — сем. Echinidae; Strongylocentrotus и Sphaerechinus — сем. Echinometridae; Arbacia — сем. Arbacidae, более отдаленное.
(обратно)
41
А также Fundulus × Menidia и Fundulus × Stenotomns.
(обратно)
42
Инертные — хромосомы, вернее большие части их, установлены точно у Drosophila в Х-хромосомах.
(обратно)
43
Можно принять предел изменчивости равным ±4σ или, наоборот, ±Зσ в зависимости от характера материала.
(обратно)
Оглавление
Предисловие
Глава I. Гибридизация как наука
Гибридизация и гибриды
Проблема овладения гибридизацией. Краткий очерк
Глава II. Роль гибридизации животных в прошлом и ее перспективы
Гибридизация в истории домашних животных
Гибридизация в капиталистический период
Научная гибридизация
Гибридизация в СССР
Гибридизация и освоение мировых зоологических ресурсов
Применение метода гибридизации
Гибридизация и доместикация
Гибридизация и адаптация (акклиматизация)
Гибридизация при воздействии на взаимоотношения животных
Гибридизация для селекции и реконструкции домашних животных
Приложение к главе II
Глава III. Проблема пределов гибридизации
Естественная гибридизация
Препятствия к гибридизации
Возможности преодоления препятствии к гибридизации
Таксономические пределы гибридизации
Глава IV. Мировые ресурсы гибридизации
Ресурсы видов
Понятие о мировых ресурсах и их классификация
Гиркон и его состав
Проблема «формы» гиркона
Ресурсы признаков
Феноресурсы
Геноресурсы
Краткий обзор главнейших гирконов
Лошади
Коровы
Овцы и козы
Марала
Северного оленя
Кролика
Курицы
Фазана
Карпа и карася
Шелкопряда
Пчелы
Глава V. Проблема гетерогенного осеменения
Получение спермы
Хирургический метод
Провоцирование эякуляции
Получение спермы нормальным половым актом
Рефлексология садки
Собирание спермы
Введение спермы в самку
Разбавление спермы
Хранение спермы
Проблема достижения сперматозоидом яйца в половых путях самок чужого вида и помощь ему в этом
Проблема своевременности гибридного осеменения
Глава VI. Проблема гетерогенного оплодотворения
Гетерогенное оплодотворение
Процесс оплодотворения и его элементы
Усиление активности сперматозоида
Привлечение сперматозоида в яйцу
Внедрение сперматозоида в яйцо
Слияние гетерогенных ядер при оплодотворении
Ультрагибридизация
Гетерогенное оплодотворение теплокровных in vitro
Глава VII. Проблема нежизнеспособности гибридов
Нежизнеспособность гибридов
Как связана жизнеспособность гибридов со степенью родства скрещиваемых видов?
Классификация явлений нежизнеспособности
Полиспермия как причина гибели гибридных зигот
Патология гибридов морских ежей
Расстройства клеточного деления в результате гибридизации
Патология гибрида как целого
Патология гибридной беременности
Патологии вылупления
Экологическая нежизнеспособность гибридов
Генетический анализ жизнеспособности гибридов
Роль цитоплазмы в определении жизнеспособности гибридов
Жизнеспособность гибридов дальнейших поколений
Проблема преодоления нежизнеспособности гибридов
Глава VIII. Проблема размножения гибридов
Значение проблемы
Вариация степени плодовитости гибридов первого поколения
Что такое бесплодие гибридов
Классификация гибридов по гаметогенезу. Работы Поля
Нарушение плодовитости у разных полов
Нарушение плодовитости в реципрокных скрещиваниях
Бесплодие в F2 и дальнейших поколения гибридов
Цитогенетика бесплодия гибридов
Причины бесплодия гибридов
Сила полоопроделяющих генетических факторов
Пропорция полов у гибридов птиц
Противоречие полового отбора
Проблема преодоления бесплодии гибридов
Глава IX. Явления наследственности при гибридизации
Особенности первого поколения гибридов
Закономерности образования признаков гибридов
Атавизация
Новитация
Гетерозис
Развитие признаков гибридов
Начало влияния сперматозоида
Наследование эмбриональных признаков у гибридов
Форма клеток гибридов
Форма хромосом гибридов
Митозы гибридов. Элиминация хромосом
Второе поколение гибридов и дальнейшие поколения
Глава X. Темпы и направление гибридизационной работы
Ускорение смены гибридных поколений
Проблема предсказания
Проблема проектирования
Список литературы
*** Примечания ***