Сергей Викторович Мейен
Следы трав индейских

МОСКВА «МЫСЛЬ» 1981
РЕДАКЦИЯ ГЕОГРАФИЧЕСКОЙ ЛИТЕРАТУРЫ
Художник С. В. Юкин
© Издательство «Мысль». 1981
ПРЕДИСЛОВИЕ
«…Удивления достойны морские черепокожие, к переселению и переведенству неудобные гадины, кои находят окаменелые на сухом пути в горах лежащие к северу, где соседственные моря их не производят, но родят и показывают воды, лежащие под жарким поясом в знатном количестве. Еще чуднее, что в холодных климатах показываются в каменных горах следы трав Индейских, с явственными начертаниями, уверяющими о подлинности их породы. Сии наблюдения двояко изъясняют испытатели натуры. Иные полагают бывшие главные земного шара превращения, коими великие оного части перенесены с места на место чрезвычайным насильством внутреннего подземного действия. Другие приписывают нечувствительному наклонению всего земного глобуса, который во многие веки переменяет расстояние еклиптики от полюса».
Это слова М. В. Ломоносова из § 163 его книги «О слоях земных» — замечательного произведения, написанного в 1757–1759 гг., но опубликованного в 1763 г. в дополнении к «Первым основаниям металлургии». С тех пор прошло более двух веков, и уже, казалось, перестали быть «удивления достойны» и морские черепокожие (моллюски) в горах, и «следы трав индейских» в холодной Арктике. Все это вошло в учебники, не раз описывалось популяризаторами. Каждый студент — биолог, геолог или географ — объяснит, не задумавшись, в чем здесь дело, расскажет в импровизированной популярной лекции, как поднимаются и опускаются континенты, приходят и уходят моря, воздвигаются горы и скользят по поверхности земной полюса. Все это уже не достойно его удивления. И зря. Мы привыкаем к картинам прошлого, восстановленным поколениями ученых, и часто забываем удивиться тому, что прошлое это действительно было, что действительно на месте городов были моря, что сушу заселяли удивительные живые существа, что — и это главное — история Земли была куда сложнее, чем мы привыкли считать. По-видимому, такова особенность человеческой психики, что он легко привыкает к необычному. Еще сто лет назад было дикостью представить себе полет не в космические дали, а всего лишь без помощи воздушного шара или машущих крыльев. Сейчас мы не поднимаем глаз на пролетающий самолет. Так же привыкаем мы к тому, что в нашей крови множество микроскопических красных телец и что землетрясения — это подвижки в глубоких земных недрах. Привыкли мы и к чудесам палеонтологии. Только школьники и далекие от естествознания люди нет-нет да и порадуют своим волнением от находки в овраге доисторических раковин или костей. В общем-то прошлым Земли и жизнью на ней люди интересуются, но степень загадочности этого прошлого как-то не осознают. Приходится упрекнуть за эту потерю удивления и популяризаторов науки, и авторов учебников. Слишком уж там все гладко, как будто все написанное незыблемо, прочно, а оставшееся непознанным — второстепенные детали. Будто в летописи, доставшейся нам от природы, мы прочли самое главное и теперь лишь подбираем отдельные недостающие листки.
Речь не идет обязательно о недоверии к нынешнему прочтению геологической летописи. Пусть все написанное в книгах — истинная правда. Но даже если это и так, то это лишь небольшой кусочек всей правды. А вообще-то ошибок в литературе немало. Недаром сказанное в одной книге противоречит сказанному в другой. Об этих расхождениях ниже пойдет речь, но не для того, чтобы потешаться над научными баталиями. Хочется, чтобы читатель почувствовал суть и состояние проблем, проникся духом непредвзятого исследования. Может быть, читатель поймет, как мало мы в сущности знаем об отдаленнейшем прошлом Земли, как увлекательно раскрывать все новые страницы и как много открытий нам еще предстоит сделать.
Как знать, может быть, найдутся у этой книги и юные читатели, которые загорятся желанием ступить на палеоботаническую стезю. Чтобы не приманивать их понапрасну, в книге специально оттенена рутинная и не слишком разнообразная сторона работы палеоботаника. И пусть потом не попрекают меня те, кто будет изнемогать от бесконечного сидения за микроскопом и подсчета многих тысяч пылинок и спор, скользящих в поле зрения, от необходимости доставать и читать статьи на всевозможных языках. Разочарование от палеоботанической деятельности, сначала казавшейся столь привлекательной, — обычное дело.
Эта книга не учебное пособие. Как ни стараться быть точным в изложении материала, каких-то упрощений невозможно избежать. Опущено многое из того, что накопила палеоботаника по истории растительного покрова. Упрощена и терминология. В книге оставлены лишь те термины, без которых невозможно вести рассказ. Это в особенности названия геологических периодов, эпох и веков (рис. 1), а также названия (особенно латинские) растений и животных. Самые важные термины поясняются, смысл других, надеюсь, будет ясен из контекста. Оставшиеся непонятными слова читатель может просто пропустить. На общем понимании текста это едва ли скажется.
 Рис. 1. Геохронологическая шкала эр (групп) и периодов (систем); линиями показано время существования основных групп высших растений
Рис. 1. Геохронологическая шкала эр (групп) и периодов (систем); линиями показано время существования основных групп высших растений
И последняя, может быть, самая важная оговорка. Чуть ли не самое интересное в науке — не полученные результаты, а сам процесс исследования. Чтобы рассказать о нем, те, кто не занимается историей науки специально, вынуждены обращаться к собственным исследованиям. И тут возникает проблема. Любое исследование начинается не с пустого места. Всегда приходится сталкиваться с работами предшественников, и далеко не всегда выводы предшественников подтверждаются. Стоит ли вспоминать чужие заблуждения, называть ошибавшихся по именам?
Думается, что стоит. Иначе получится искаженная и безликая картина исследования. Только важно помнить, что вскрывшиеся непроизвольные просчеты не компрометируют настоящего ученого. Более того, порой они сильнее стимулируют дальнейшие исследования, чем правильные выводы. В конце концов самую грубую ошибку делает тот, кто уверен в собственной непогрешимости.
Глава I
ЗАВОЕВАНИЕ СУШИ

В истории нашей Земли было немало вех, значение которых для правильного прочтения всей истории ясно каждому исследователю и которые тем не менее так и остаются загадкой. Мы убеждены, что когда-то не было самой Земли, а после того, как она образовалась, на ней появилась жизнь. Не всегда населяли Землю окружающие нас животные и растения. Совсем недавно в масштабе геологического времени появился сам человек. Эти и другие важнейшие события — от возникновения Земли до происхождения человека — источники неисчислимых гипотез. Время от времени то одна, то другая завоевывает чуть ли не всеобщее признание, но потом тускнеет под напором новых идей. Калейдоскоп гипотез рождает скепсис. Порой начинает казаться бессмысленной игра фантазии вокруг событий, не оставивших следов в геологической летописи. Исследователи обращаются к другим проблемам, и тут вдруг обнаруживаются новые факты, заставляющие вернуться к оставленной проблеме и попытаться решить ее иначе.
Именно так получилось с завоеванием суши растениями. От того времени, как зародилась на Земле жизнь, и до тех геологических эпох, когда на суше появились первые леса, прошли миллиарды лет. Растительность вполне современного облика покрывала континенты всю последнюю эру — кайнозойскую. В мезозое и в двух последних периодах палеозойской эры — пермском и каменноугольном — суша тоже была покрыта густой растительностью. Известны многочисленные остатки наземных растений и в предшествовавшем девонском периоде. А дальше? Что было на суше в силуре, ордовике, кембрии и в еще более древние — докембрийские — времена? Этого никто не знает.
Вольно или невольно мы навязываем далекому прошлому черты мира, окружающего нас сейчас. Ставшее привычным кажется непреходящим. Как ребенку трудно представить, что когда-то не было мамы и папы, дома, в котором он живет, так и исследователи с трудом приходят к мысли, что привычные черты нынешнего облика планеты — нечто появившееся не так уж давно. Особенно трудно свыкнуться с мыслью, что когда-то облик планеты был настолько иным, что взгляни мы на нее в иллюминатор самолета, то и не узнали бы Земли, а решили бы, что под нами неведомое небесное тело. С точки зрения геолога, привыкшего к срокам в сотни миллионов и даже миллиарды лет, Земля была именно такой, чужой и непривычной, не так уж давно — в середине палеозойской эры.
Надо честно признаться, что мы так и не знаем, как выглядела бы Земля с птичьего полета в начале девонского периода — 400 млн. лет назад. Мы, конечно, не увидели бы лесов. Пейзаж скорее напомнил бы нам полупустыни в дни, когда снег уже сошел, а зелень еще не появилась. Может быть, только по берегам рек и морей вилась тонкая ниточка зелени. Так считают многие палеоботаники. Не раз высказывалось мнение, что различные низшие растения заселяли сушу и в додевонские времена. Но никаких их остатков до сих пор не находили. Гипотезы о заселении суши растениями могут опираться лишь на косвенные свидетельства.
Почему-то заселение суши растениями далеко не всеми исследователями признается проблемой. Некоторым кажется, что здесь не над чем особенно ломать голову. Разве не очевидно, что растения вышли когда-то из моря на сушу? Сначала они заселили мелководье, двинулись вдоль рек в глубь континентов, растеклись по берегам озер, а потом шаг за шагом, все дальше и дальше от воды. Надо лишь разыскать этих пионеров в древнейших континентальных толщах и прочитать по ископаемым остаткам, когда и как все было. Но и без этих находок дело казалось в целом ясным. В начале 40-х годов один крупный ученый без колебаний писал: «…первый расцвет жизни на суше был приурочен к приморским местностям, особенно к берегам лагунных озер и болот, лежащих между песчаными береговыми валами и подгорной террасой. Здесь впервые выработался тип растения, нижней своей частью находящегося в воде, а верхушкой — в воздушной среде под прямыми лучами солнца. Споры, развившиеся в мелкой нагретой воде лагун, при понижении уровня воды вследствие испарения образовывали заростки в виде плоских зеленых лопастных пластинок. Отдельные участки их, выдвигаясь из воды и испаряя воду, вызывали возникновение токов воды, возмещавших испаряемую, и этим способствовали росту своих тканей. Образовывалось подобие первичного стебля».
В таком же стиле, без тени сомнения описывались и дальнейшие события. В литературе тех и последующих лет можно найти немало подобных рассказов. Что это — считанная с документов история или правдоподобный домысел? Сейчас это выглядит почти как домысел, но в начале 40-х годов дело представлялось иначе. Тогда еще свежи были в памяти открытия конца 10-х — начала 20-х годов, когда англичане Р. Кидстон и В. Лэнг описали из девона Шотландии прекрасно сохранившиеся остатки простейших с виду высших растений. Особое впечатление произвели ринии — голые безлистные прутья с мешочками-спорангиями, содержавшими споры, на концах. Лучшего предка для всех высших растений нельзя было и желать. Присовокупляя другие известные к тому времени девонские растения, ученые стали конструировать генеалогические деревья, домысливая образование листьев, корней, стволов, разнообразных органов размножения. Некоторые из предложенных эволюционных схем были затем подтверждены новыми находками девонских растений.
Естественно, что взгляды исследователей обратились к более древним, додевонским отложениям — силурийским, ордовикским и кембрийским. Остатки, внешне похожие на высших наземных растений, в них попадались давно. Это были какие-то подобия стеблей, обрывки тканей, сходных с древесиной, микроскопические оболочки, похожие на споры высших растений. Особенно широко обсуждались открытия в кембрии Сибири остатков, названных Aldanophyton (фиг. I), и якобы спор высших растений в кембрийских и даже докембрийских отложениях Прибалтики. О находке спор сообщалось в печати как о важном открытии, Об этом писали в газетах и научно-популярных журналах.
В 50-х годах сложились две точки зрения на проблему заселенности суши растениями в додевонские времена. Одни исследователи, условно назовем их «энтузиастами», говорили, что суша была заселена высшими растениями даже в докембрийские времена. Более осторожные из них соглашались, что все додевонские документы, если брать их порознь, недостоверны. Но в сумме, считали они, «улик» вполне достаточно. Другие исследователи, «скептики», соглашались учитывать только неоспоримые документы.
К 60-м годам сообщений о додевонских находках накопилось столько, что настало время как следует в них разобраться. Пошли на строгий экзамен микроскопические оболочки, описанные как споры. В 1965 г. в «Палеонтологическом журнале» вышла статья Н. А. Волковой. Она тщательно проанализировала все сообщения о докембрийских, кембрийских и ордовикских спорах. Выяснились любопытные закономерности. Прежде всего оказалось, что наиболее важные находки иллюстрированы не фотографиями, а схематическими рисунками. Чтобы отличить споры высших растений от оболочек микроскопических водорослей (внешне эти два типа остатков могут быть похожими), очень важно найти трехлучевую фигуру, которая образуется у спор еще тогда, когда они сидят в спорангиях (фиг. II). Материнские клетки спор делятся, и из каждой клетки формируются четыре споры, расположенные крест-накрест. Такая группа из четырех спор называется тетрадой. Трехлучевая фигура (она называется еще трехлучевой щелью или тетрадным рубцом) обращена внутрь тетрады. Через трехлучевую щель происходит прорастание спор, когда они, высеявшись из зрелых спорангиев, попадают на подходящую почву. О присутствии трехлучевой щели у древнейших спор и писали «энтузиасты». Однако ни в одном случае это наблюдение не было подтверждено фотографиями. Трехлучевые щели изображались только на рисунках. В палеонтологии часто бывает так, что некоторые детали ископаемых организмов хорошо видны наметанному глазу, но подтвердить наблюдение фотографией не удается. Например, швы на ископаемых черепах иногда маскируются рельефом кости. Поэтому на фотографиях получается бугристая поверхность, и проследить на этом фоне тонкие линии швов невозможно. В таких случаях вместо фотографии документом служит рисунок. Но при трехлучевой щели спор дело обстоит иначе. Здесь всегда есть возможность сделать вполне убедительные фотографии. Тем не менее их не было.
Н. А. Волкова обнаружила и другую закономерность. В работах 40—50-х годов ссылки на находки древнейших спор звучали уверенно и количество спор в препаратах указывалось большим. В начале 60-х годов те же исследователи, описывая тот же материал, что и раньше, уже писали, что количество спор с трехлучевой щелью очень небольшое. Все чаще слышались оговорки, что, дескать, это споры мхов и других сравнительно низко организованных высших растений. Энтузиазм начал давать трещину. К концу 60-х годов «энтузиастов» почти не стало слышно.
Не надо воспринимать эту историю как некое умышленное введение в заблуждение одними учеными других, а эти другие, дескать, разоблачили очковтирателей. Дело было отнюдь не так. Заблуждение было искренним, и не честь мундира спасали те, кто продолжал настаивать на присутствии спор в древнейших, даже докембрийских, толщах. Что же касается злополучной трехлучевой щели, то за нее принимали случайные складки на микроскопических оболочках водорослей. При жизни эти оболочки были шаровидными. Попав в породу, они сплющивались, и возникала сложная, беспорядочная система складок. Найти в переплетении складок трехлучевую фигуру не сложнее, чем увидеть неведомого зверя в прихотливо изогнутом лесном корне или голову старика в силуэте одного из Красноярских столбов.
Не забыли скептики и о крупных растительных остатках из додевонских отложений. С большинством остатков разделались быстро. На них не обнаружилось никаких деталей, подтверждавших, что перед нами действительно наземные растения. Труднее пришлось с сибирским алданофитом. В 1971 г. в Новосибирске состоялась Международная палинологическая конференция, на которую съехались авторитетные и весьма скептически настроенные палеоботаники. Лучшая коллекция алданофитов хранится сейчас в Томском университете у проф. А. Р. Ананьева. Он привез несколько образцов в Новосибирск, где состоялся своеобразный консилиум. Отношение к образцам было непредвзятое, но придирчивое. Смотрели на образцы в лупу и под микроскопом, смачивали ксилолом, чтобы ярче проявились детали. Заключение было единодушным: алданофит — неизвестно, что такое. Внешне он действительно похож на некоторые палеозойские плауновидные, но никаких деталей, подтверждающих общее впечатление, не видно. Главное, что нет никаких, даже самых слабых, намеков на проводящую систему. Это может быть и водоросль, и какое-то неведомое животное.
Обсуждая вопрос о природе алданофита, участники консилиума помнили и об условиях его захоронения. Это карбонатные морские отложения, образовавшиеся в открытом море. Как могли попасть так далеко от берега остатки наземных растений и не разумно ли ждать больше их остатков в отложениях, откладывавшихся ближе к берегу? Направление, в котором проходила береговая линия этого моря, примерно известно. Но находок алданофитов больше нигде не было. В общем разошлись участники консилиума, пожимая плечами и отказываясь высказываться об алданофите хоть сколько-нибудь определенно.
 Рис. 2. Уровень появления в геохронологической шкале основных структур высших растений: 1 — трахеиды; 2 — устьица; 3 — дихотомически ветвящиеся оси; 4 — побеги с главной осью; 5 — выросты (эмергенцы) на оси, не снабженные проводящим пучком; 6 — то же с проводящим пучком; 7 — вильчатые листоподобные органы; 8 — уплощенные листья; 9 — звездчатый в сечении проводящий цилиндр в оси; 10 — округлые спорангии на концах осей; 11 — эллиптические спорангии; 12 — собрания спорангиев; 13 — спорангии, раскрывающиеся продольной щелью; 14 — спорангий в пазухе листа; 15 — двурядное расположение спорангиев с придатками между ними; 16 — споры с трехлучевой щелью и без орнаментации оболочки; 17–23 — споры с различной орнаментацией и разного размера; во второй колонке слева яруса (века) силура и нижнего (раннего) девона (по В. Г. Чалонеру, с изменениями)
Рис. 2. Уровень появления в геохронологической шкале основных структур высших растений: 1 — трахеиды; 2 — устьица; 3 — дихотомически ветвящиеся оси; 4 — побеги с главной осью; 5 — выросты (эмергенцы) на оси, не снабженные проводящим пучком; 6 — то же с проводящим пучком; 7 — вильчатые листоподобные органы; 8 — уплощенные листья; 9 — звездчатый в сечении проводящий цилиндр в оси; 10 — округлые спорангии на концах осей; 11 — эллиптические спорангии; 12 — собрания спорангиев; 13 — спорангии, раскрывающиеся продольной щелью; 14 — спорангий в пазухе листа; 15 — двурядное расположение спорангиев с придатками между ними; 16 — споры с трехлучевой щелью и без орнаментации оболочки; 17–23 — споры с различной орнаментацией и разного размера; во второй колонке слева яруса (века) силура и нижнего (раннего) девона (по В. Г. Чалонеру, с изменениями)
На XI Международном ботаническом конгрессе в 1969 г. собрался симпозиум по основным событиям в эволюции растений, документированным палеоботаникой. Английский палеоботаник В.-Г. Чалонер собрал все доступные к этому времени материалы о древнейших наземных растениях. Его доклад был затем опубликован. Чалонер рассуждал так: есть три признака, совместное присутствие которых у ископаемого растительного остатка позволяет считать, что перед нами остаток высшего растения, — это древесинные клетки (трахеиды), споры с трехлучевой щелью и устьица на поверхности тела. Устьица можно видеть, сняв с отпечатка покрывающую его пленку (кутикулу)! с оттисками клеток. Кутикула состоит из высокополимерного органического вещества и хорошо сохраняется в ископаемом состоянии. Обычно ее отделяют, растворяя обугленный остаток растения в смесях, не действующих на вещество кутикулы. Чалонер учитывал, что часто мы встречаемся с такими остатками, у которых нельзя изучить все три признака. Поэтому он предложил оценивать время появления высших растений иначе. Он свел в одну таблицу все известные к тому времени находки, позволившие наблюдать хотя бы один из трех признаков (рис. 2). Оказалось, что первые споры с настоящей трехлучевой щелью появляются с самого начала силурийских отложений, а остатки с кутикулой и устьицами или с трахеидами — с самого конца силура. Все сообщения о более древних находках остатков с такими признаками оказались сомнительными. В одних случаях не были надежно датированы породы. В других — остатки оказались вмытыми по трещинам из более молодых отложений. В третьих случаях сообщения не подкреплялись убедительными иллюстрациями.
Скрупулезный анализ, выполненный Чалонером, убедил многих, что о появлении высших растений не то что в докембрии, кембрии и ордовике, но даже в начале силура нельзя говорить хоть сколько-нибудь уверенно. Правда, исследователей смущали находки спор с трехлучевой щелью по всему разрезу силурийских отложений (хотя и не во всех местах, где эти отложения известны). Впервые раннесилурийские споры обнаружил и изобразил на хороших фотографиях У. С. Гофмейстер в 1959 г. Его материал происходил из Ливии. Затем число находок стало увеличиваться, причем исследователи заботились о том, чтобы наблюдения были хорошо документированы (фиг. III). В 1971 г. Д. Грей и А. Буко описали нижнесилурийские споры в виде еще не распавшихся тетрад.
Эти находки спор в раннесилурийских отложениях не были случайностью. Дело в том, что для изучения силурийских и более древних пород стала все шире применяться методика спорово-пыльцевого анализа. Куски породы дробят и растворяют в кислотах, органические остатки просветляют и отделяют специальными жидкостями и рассматривают в микроскоп. При этом находят большое количество одноклеточных водорослей и проблематичных микроскопических остатков. По их составу удается сопоставлять разрезы осадочных толщ. Среди таких остатков и попадаются споры. Одновременно стали встречаться и интересные обрывки, возбудившие дискуссии. Во-первых, это были трубки со спиральными утолщениями. Во-вторых, нашли органические пленки с отпечатавшейся системой многоугольных клеток (рис. 3).
 Рис. 3. Органические пленки с клеточной структурой и трубки со спиральными утолщениями из нижнего силура Вирджинии; длина линейки — 10 микрон (по Х. П. Бэнксу, Л. М. Пратт, Т. Л. Филлипсу и Д. М. Деннисону)
Рис. 3. Органические пленки с клеточной структурой и трубки со спиральными утолщениями из нижнего силура Вирджинии; длина линейки — 10 микрон (по Х. П. Бэнксу, Л. М. Пратт, Т. Л. Филлипсу и Д. М. Деннисону)
Некоторые исследователи стали высказывать мнение, что трубки со спиральными утолщениями не что иное, как трахеиды, а органическая пленка — кутикула высших растений. Один из наиболее авторитетных исследователей древнейших растений, X. П. Бэнкс из Йельского университета, в специальной статье, опубликованной в 1975 г. в международном журнале, призвал коллег к осторожности. Он писал, что спиральные утолщения на трубках иные, чем у трахеид растений. У нижнесилурийских трубок утолщение как бы встроено в стенку, а у растений оно наложено на нее. По мнению Бэнкса, такие трубки вполне могут принадлежать животным. К животным же или к каким-то неизвестным водорослям могут относиться и фрагменты с клеточной структурой. Споры с трехлучевой щелью тоже не обязательно принадлежат высшим растениям. Сходные споры находили у некоторых очень своеобразных ископаемых растений (протосальвиний), относимых сейчас обычно к водорослям.
Статья Бзнкса не осталась без внимания. Два года спустя в другом международном журнале Д. Грей и А. Буко в специальной большой статье возразили Бэнксу. Они писали о том, что ни у одной известной группы животных нет таких трубок, что клетки на кутикулах больше похожи на растительные и что ни у одной настоящей водоросли не бывает спор с трехлучевой щелью и такой оболочкой, которая бы выдерживала обработку спорово-пыльцевыми методами.
Стоит напомнить и еще об одном открытии последних лет. После того как было показано, что палинологи ошибочно дорисовывали трехлучевую щель у микроскопических оболочек раннего палеозоя и докембрия, Н. А. Волкова обнаружила в самых верхах докембрия очень мелкие (диаметром до 10 микрон) оболочки. У них была совершенно ясная трехлучевая щель — ее каждый может видеть на опубликованных фотографиях (фиг. IV). Тем не менее эта находка, видимо, не имеет прямого отношения к происхождению высших растений и заселению ими суши. Дело в том, что в породах, содержащих эти оболочки, в большом количестве встречаются загадочные пока водоросли, изучавшиеся М. Б. Гниловской и отнесенные ею к особой группе вендотенид. Их связь с другими водорослями остается неизвестной, поскольку не вполне понятна их структура и, в частности, органы размножения. Вполне возможно, что обнаруженные Волковой оболочки (споры —?) принадлежат вендотенидам. Для нашей темы в этой находке важно то, что не только у высших растений могут образовываться споры с такой оболочкой, которая могла бы выдерживать химическую обработку, применяемую в палинологии. Впрочем, и здесь рано строить гипотезы. Еще предстоит изучить эти древнейшие остатки с помощью электронной микроскопии.
До окончания начавшейся дискуссии еще далеко, и сейчас еще слишком мало материалов, чтобы отдавать предпочтение какой-либо точке зрения. Нужны более детальные исследования всех этих странных остатков. Может быть, удастся изучить их химический состав или тонкую структуру, различимую лишь в электронный микроскоп.
Однако не только дальнейшее накопление фактических данных и сравнение остатков с другими организмами помогут решению вопросов. Надо поразмыслить над уже собранным материалом и еще раз подумать о выводах из имеющихся наблюдений.
Существует широко распространенный миф, что наука развивается от фактов к обобщениям. Над опровержением этого мифа, бытующего главным образом среди исследователей с эмпирическим складом ума, трудилось несколько поколений философов и методологов науки. Никто не спорит, что иногда случайное наблюдение приводит к крупнейшему научному открытию. Но даже и в этом случае оценить значимость факта можно лишь с помощью теории. Факты питают теорию, теория через обобщения и дедукции ведет к новым осмысленным наблюдениям и экспериментам. Связь фактов и обобщений, индукций и дедукций, теории и наблюдения непрерывна и взаимна.
С этой точки зрения интересно проследить развитие взглядов на заселение суши и разобраться, как влияли на обобщения, с одной стороны, новые находки, а с другой — отказ в доверии прежним наблюдениям. Здесь невозможно изложить в деталях всю историю. Поэтому ограничимся ее главными чертами. Мнение о том, что высшие растения происходят от водорослей, высказывалось еще в прошлом веке. В первой трети нашего века уже детально расписали всю последовательность выхода растений на сушу. Наглядные схемы показывали, как некие водоросли, жившие вблизи берега, сначала высунули на воздух «голову», затем заселили приливно-отливную зону, потом, постепенно превращаясь в высшие растения, целиком вышли на берег. За этим последовало постепенное завоевание суши.
Какие же наблюдения легли в основу этой гипотезы, обычно преподносимой в литературе — научной, учебной, популярной — чуть ли не как хорошо обоснованная историческая реконструкция, для которой нет и не может быть альтернатив? Как ни странно, таких наблюдений не было ни в те времена, когда эта гипотеза выдвигалась, ни потом. Это было и остается чисто умозрительным построением. В основном исследователи опирались на очень общие филогенетические соображения и косвенные аналогии.
После выхода в свет «Происхождения видов» Ч. Дарвина начала развиваться филогенетическая систематика организмов. Систематические группы были отождествлены с ветвями филогенетического древа. Особую роль в развитии этих взглядов сыграл Э. Геккель. В 1866 г. во втором томе своей «Общей морфологии» он нарисовал филогенетическое древо всех организмов. Настал период конструирования филогенетических деревьев, одно время это было повальным увлечением. В ботанике еще в прошлом веке была построена филогенетическая линия, уходящая от покрытосеменных к голосеменным, а от них — к высшим споровым растениям. Дальше эта линия обрывалась. Никаких фактических данных о происхождении высших растений в XIX в. по существу не было.
Как обычно бывает в филогенетике (так же поступают при установлении отцовства), предков подбирали методом исключения. После того как зоологи построили филогенетическую линию, идущую от млекопитающих через рептилий и амфибий к рыбам, возникло представление, будто население суши произошло от обитателей морей. Вполне естественной показалась и мысль, что растения суши тоже взяли начало от водных растений. Вывести высшие наземные растения из высших же, но водных растений было невозможно. Взгляды исследователей обратились к водорослям. Дальше пошли методом исключения. Из современных многоклеточных водорослей только зеленые, красные и бурые могли быть претендентами на предковое положение. Сейчас большинство филогенетиков, интересующихся происхождением высших растений, выводят их из зеленых водорослей, хотя есть сторонники ставить в корни высших растений бурые и красные водоросли.
К сожалению, как в прошлом веке, так и сейчас палеоботаника ничего не может предложить участникам спора. Она свидетельствует только о том, что в кембрии и ордовике уже были разнообразные морские водоросли. Хотя вывести из этих древнейших водорослей высшие растения не легче, чем из современных, этот факт важен. Он позволил заключить, что многоклеточные морские водоросли старше высших растений и потому действительно могли быть их предками. Дальше было достаточно не удержаться от соблазна превратить принципиальную возможность в историческую гипотезу. А чтобы гипотеза превратилась в убеждение, достаточно привыкнуть к ней. И еще надо, чтобы у гипотезы не появилось серьезных противников.
Гипотеза о происхождении высших растений от морских водорослей подкреплялась и косвенной аналогией. По берегам современных морей можно видеть несколько поясов растительности. Некоторые водоросли всегда живут в воде. Другие заселяют приливно-отливную полосу. Третьи довольствуются лишь брызгами прибоя. Им хватает этой воды для жизни и размножения. Немало водорослей живет на суше. Этот ряд вполне можно прочитать как протокол выхода растений из моря на сушу. И даже если не делать этого, вполне можно предположить, что, следуя похожему ряду, растения когда-то выбирались на сушу, попутно становясь настоящими высшими растениями, развивая способность образовывать стебли, проводящую систему, корни, распространять споры по воздуху, обходиться без водной среды при размножении. Короче говоря, история выхода растений на сушу и возникновения высших растений не была реконструирована по палеоботаническим документам, а была гипотетически вычитана из разнообразия форм и условий жизни современных растений.
Сам по себе этот прием неизбежен, когда мы хотим реконструировать прошлое или когда невозможно проследить все стадии процесса, происходящего в наше время. В этих случаях исследователи пользуются методом, который известный французский философ А. Бергсон назвал «кинематографическим». Приняв, что некие объекты развиваются по одному и тому же правилу, мы наблюдаем разные фазы развития на разных объектах, получаем отдельные «кадры» происходящего процесса, а затем мысленно нанизываем эти кадры на воображаемую киноленту. Так мысленно восстанавливается процесс. Именно таким способом Гарвей когда-то пришел к выводу, что не только птицы, но и млекопитающие развиваются из яйца. Эмбриология научилась прослеживать онтогенез одной яйцеклетки не так уж давно, а наиболее фундаментальные открытия были сделаны отцами эмбриологии К.-Э. фон Бэром и К.-Ф. Вольфом с помощью кинематографического метода. Они прослеживали разные стадии развития зародыша, сравнивая разные зародыши.
Так же поступают и геологи. Когда они утверждают, что море размывает берег, окатывает камни и намывает пляжи, то это не значит, что они годами сидели на одном месте и следили, как море, подмыв в каком-то одном месте берег, взялось за рассыпавшиеся камни и одни из них окатало в гальку, а другие истерло в песок. Все происходило иначе. Геологи сравнивали форму камней, разбросанных вдоль берега и в разной степени окатанных, сравнивали прибрежные скалы и рассеянные в воде камни, анализировали минеральный состав песка на пляже и пород на прилежащем берегу. Они видели, как время от времени происходят обвалы берегов, как таскает прибрежная волна песок и камни. Так реконструировались кадры, а из них складывалась «кинолента» процесса, происходящего хотя и на наших глазах, но чрезвычайно медленно.
Примерно так же поступили и при реконструкции заселения суши растениями. Были взяты две «киноленты». Одна изображала происхождение высших растений от морских водорослей и была сначала составлена только из современных растений, а затем современные растения были заменены девонскими формами. Другая «кинолента» была составлена из кадров, изображающих современные растения, живущие вблизи береговой линии водоемов. Совместив эти ленты, мы получим нынешние представления о том, как растения завоевывали континенты. Эта схема стала подтверждаться палеонтологическими документами уже в нашем веке, когда начались детальные исследования девонских растений и когда удалось как следует разобраться в последовательности напластований силурийских и девонских отложений. Если оставить в стороне вызывавшие споры находки ордовикских и более древних растений, а взять только несомненные остатки высших растений девона и верхов силура, то складывается такая картина. В самых верхах силура встречаются очень однообразные остатки простейших высших растений, относимых к роду куксония и внешне сходных с риниями. В низах девона, отвечающих жединскому веку, примерно та же картина. В отложениях следующего, зигенского века мы находим уже множество разнообразных высших растений, принадлежащих нескольким родам (рис. 4). С этого времени высшие растения разбиваются на две группы. Одна из них пойдет к плауновидным, в том числе к древовидным лепидодендронам каменноугольного периода (они сыграли большую роль в накоплении углей Донбасса, западноевропейских и североамериканских угленосных бассейнов). Сейчас потомки этой группы представлены травянистыми плаунами и немногими другими растениями. Вторая группа поведет к папоротниковидным, голосеменным и покрытосеменным.
Детальное изучение самих девонских растений и тех отложений, в которых они погребены, показало, что некоторые из них вели полуводный образ жизни. Нижними частями они сидели в воде и высовывали наружу лишь оси с органами размножения. Казалось бы, все свидетельствовало о том, что в отложениях верхов силура — нижней части девона запечатлен процесс выхода растений на сушу.
Эта вполне правдоподобная картина тем не менее не дает ответа на два вопроса. Во-первых, непонятно, кому принадлежат споры с трехлучевой щелью, встречающиеся в более древних силурийских отложениях. Где те растения, которые продуцировали эти споры? На материале из Прибалтики было показано, что оболочки таких спор закономерно распространены в морских осадках. Силурийская палеогеография Прибалтики реконструирована довольно хорошо. Во всяком случае известно, с какой стороны располагался здесь берег моря. Если наложить на палеогеографическую карту количество трехлучевых спор, встречающихся в пробах, то можно ясно видеть, как обилие спор увеличивается по направлению к берегу. В противоположном направлении в пробах увеличивается количество остатков различных морских водорослей. Д. Грей и А. Буко, собравшие материалы, сделали вывод, что эти споры принадлежат высшим наземным растениям. Они отметили сходство силурийских спор с теми, которые извлечены из спорангиев высших растений девона.
 Рис. 4. Различные девонские проптеридофиты («псилофиты»): вверху слева и в центре — савдония (Sawdonia, по А. Р. Ананьеву и С. А. Степанову); справа вверху и внизу — пертика (Pertica, no Э. Касперу и Г. Н. Эндрьюсу); внизу слева — зостерофилл (Zosterophyllum, по Д. Уолтону)
Рис. 4. Различные девонские проптеридофиты («псилофиты»): вверху слева и в центре — савдония (Sawdonia, по А. Р. Ананьеву и С. А. Степанову); справа вверху и внизу — пертика (Pertica, no Э. Касперу и Г. Н. Эндрьюсу); внизу слева — зостерофилл (Zosterophyllum, по Д. Уолтону)
Получается какая-то несуразица. Споры вроде бы документировали заселение суши уже с начала силура. А остатки стеблей и других частей растений документируют заселение суши в конце силура — начале девона. Длительность силура сейчас принимают примерно в 35 млн. лет. В масштабе всей земной истории, длившейся миллиарды лет, это не так много, но для тех, кто занимается силурийскими и близкими по возрасту отложениями, расхождение почти на целый период обращается в невыносимое противоречие.
Второй вопрос, на который не дает ответа принятая сейчас гипотеза, касается растений верхов силура и девона. После того как были открыты и изучены различные девонские растения, в литературе появились реконструкции девонских ландшафтов. На них изображают берег моря, вдоль которого и чуть поодаль рассажены ринии и их современники. Некоторые из них лишь торчат из воды, а остальные вполне освоились на суше. Пейзаж довольно унылый, ибо население берега низкорослое, листьев еще нет, одни только голые или шиповатые прутья. Вот здесь-то и возникает вопрос: как эти растения после смерти попадали в захоронения? Известно, что растительный остаток может захорониться на века, лишь попав в воду. Иначе при доступе кислорода он быстро истлеет. Обычно палеоботаники имеют дело с листьями, опавшими с деревьев и занесенными в воду, болотными и водными растениями. Если растения живут в стороне от воды, то скорее всего от них сохранятся лишь рассеявшиеся по воздуху споры, пыльца или семена. Обычная трава, растущая по берегам рек и озер, может захорониться на века лишь в исключительных случаях. Поэтому об истории злаков и полыней палеоботаники судят только по остаткам пыльцы.
В девоне настоящих болот еще не было. Поэтому мы не находим в девонских отложениях пластов углей, а те, которые известны, образовались из остатков водорослей (их раньше ошибочно принимали за остатки высших растений). В конце силура — начале девона не было растений с опадающими листьями, так как еще не было самих листьев. Почему же низкорослые девонские растения, изображаемые на реконструкциях по берегам водоемов, попадали в воду и захоронялись? Какие у них были для этого преимущества перед современной травой?
На эти вопросы нет ответа, поскольку исследователи девонских растений не задали себе таких вопросов. Тогда возникает другой вопрос: правильны ли реконструкции девонских ландшафтов, вошедшие в литературу? На каком основании ринии и их современники рассажены по берегам? Может быть, они сидели в воде? И еще такие вопросы: если появление крупных остатков в отложениях верхов силура — низов девона свидетельствует о выходе растений на сушу, то почему мы не видим всех стадий самого процесса преобразования водорослей в высшие растения? Ведь в таком случае все происходило в прибрежных водах, где условия для сохранения геологической летописи наиболее благоприятны. Прибрежно-морские отложения известны в отложениях всех геологических периодов и служат одним из главных источников информации о жизни в геологическом прошлом.
Вспомним теперь упомянутое соображение, что некоторые девонские растения были полуводными. А почему, собственно, некоторые? Почему все те растения, чьи остатки изучают палеоботаники и чьи реконструкции вошли в сводки и учебники, не могли быть водными и полуводными? И почему появление крупных остатков растений в верхах силура — низах девона надо обязательно толковать как следы выхода растений на сушу? Может быть, это следы переселения высших растений в воду. Именно так: не из воды, а в воду.
Достаточно принять эту гипотезу, как многое становится на свои места. Прежде всего находит объяснение факт, что в нижнедевонских отложениях богатство спор гораздо больше, чем можно предполагать по крупным остаткам растений. Это указывает на то, что какие-то растения сидели на берегу и только их споры долетали до мест захоронения. Тогда и силурийские (в том числе и нижнесилурийские) споры можно считать спорами тех растений, которые росли на берегу, так что их стебли не могли добраться до воды и попасть в геологическую летопись. Правда, здесь можно возразить, что, дескать, мы относим время выхода растений из моря на сушу на начало силура и лишь отодвигаем ту же проблему в глубь веков, а не решаем ее. Ведь и в начале силура процесс выхода растений на сушу должен был пройти через прибрежно-морскую полосу, где неплохо ведется геологическая летопись. Записей же об этом процессе в начале силура нет. Стало быть, нет и опровержения гипотезы, что растения действительно вышли из моря и по мере выхода водоросли превращались в высшие растения. Опровергнуть эту гипотезу действительно нет возможности. Но это не достоинство, а дефект любой гипотезы. Неопровергаемость превращает гипотезу в догмат. Эта же неопровергаемость, правда, дает возможность выдвигать иные гипотезы. Скажем, такую: почему бы не предположить, что заселение суши водорослями произошло в досилурийские времена, а преобразование водорослей в высшие растения происходило целиком в наземных условиях? На какой-то стадии этого процесса водоросли обрели способность обтягивать свои споры прочными, химически устойчивыми оболочками. Это событие мы и отмечаем в геологической летописи, когда оболочки спор появляются в силурийских породах. К концу силура уже развились несомненные высшие растения и некоторые из них отправились жить в воду, причем совершенно не обязательно «обратно» в воду.
Ничего экстраординарного в таком процессе нет. Мы знаем, что многие сухопутные организмы со временем заселяли воду. Это ихтиозавры и плезиозавры из рептилий, китообразные и ластоногие из млекопитающих, многие насекомые. Есть водные цветковые растения, хотя их непосредственные предки, судя по всему, не жили в воде. Для природы такие переходы не составляли какой-то исключительной трудности. Условия жизни в воде и на суше различаются очень сильно, но, может быть, не для всех организмов эти различия действительно существенны. Для человека немыслимо поселиться в воде без специальных технических приспособлений. Другие организмы прекрасно чувствуют себя и на земле, и в воде (достаточно вспомнить лягушек). Многим из них важно только не уходить от воды далеко или иметь хотя бы временами, например для размножения, влажную среду вокруг.
Конечно, высказанная здесь гипотеза может вызвать тот же упрек в недоказуемости, а стало быть, и догматичности. Действительно, пока трудно указать пути ее проверки. Все же кое-что можно
предпринять. Например, было бы весьма важно не ограничиваться рассматриванием силурийских спор в обычные световые микроскопы. Сейчас начали изучать их с помощью электронного сканирующего микроскопа, который позволяет рассмотреть мельчайшие детали поверхности спор, но не позволяет заглянуть внутрь. Для этого нужен просвечивающий электронный микроскоп. Очень важно разобраться в тонкой внутренней структуре оболочек этих спор, сравнить ее со структурой спор водорослей и высших растений девона. Палеоботаники, имеющие дело с девонскими растениями, обращали много внимания на их структуру и систематику, но почти не занимались их экологией. Теперь требуется тщательно изучить особенности их захоронения, происхождения и условий образования толщ, содержащих растительные остатки. Детальный анализ палеогеографической приуроченности силурийских спор пока был выполнен только в Прибалтике. Мы не знаем, насколько выдерживаются в других местах установленные в Прибалтике закономерности.
Надо более обстоятельно заняться и теми загадочными остатками, из-за которых возникла дискуссия между Грей и Буко, с одной стороны, и Бэнксом — с другой. Вовсе не обязательно ставить в прямую зависимость эти остатки и находки спор в силурийских отложениях. Вполне возможно, что споры принадлежат наземным и, может быть, даже высшим растениям, обрывки с клетками — каким-то неведомым водорослям, а трубки — неведомым животным. Пока все эти остатки изучаются лишь под микроскопом. Не обращаясь к более совершенным физическим и химическим методам, спорить дальше об их природе бессмысленно.
Наконец, надо внимательнее отнестись к додевонским отложениям, континентальным по происхождению. А. Грей и Буко отметили, что в тех додевонских толщах, которые считаются наземными, нет остатков спор. Вскоре после выхода их статьи американские палеоботаники Л. М. Пратт, Т. Л. Филлипс и Д. М. Деннисон описали все те же трубки, споры и кутикулы из нижнесилурийских отложений штата Вирджиния. По составу, характеру слоистости и условиям залегания эти отложения считаются континентальными. Еще один американский палеоботаник, Д. М. Шопф, присоединившийся к дискуссии, обратил внимание на то, что трубки со спиральными утолщениями обычно попадаются в захоронениях вместе с хитинозоями — проблематическими мелкими животными, обычно встречающимися в морских отложениях.
Словом, пока еще слишком рано уверенно говорить о том, к каким организмам относятся силурийские спороподобные остатки, трубки и куски кутикулы с клетками. Лучше повременить и с уверенными заключениями о том, что же было на суше в додевонские времена.
Впрочем, и в отношении раннего девона надо высказываться осторожно. В самых низах девонского разреза растительных остатков не многим больше, чем в верхах силура. Лишь в отложениях зигенского века (второго от начала девонского периода) во многих местах Земли находят разнообразные и многочисленные остатки растений. Гораздо разнообразнее становятся и оболочки спор. Это увеличение числа и разнообразия растительных остатков указывает не только на успехи в эволюции растений, но и на увеличение площадей, пригодных для устойчивого захоронения растительных остатков. Многие зигенские толщи с растительными остатками заведомо отложились на континентах.
Редкость континентальных отложений в дозигенские времена можно объяснить отсутствием развитого растительного покрова. Без него не было настоящих почв, не было и регуляции стока вод. После каждого дождя вода скатывалась в низины беспорядочными и ничем не сдерживаемыми потоками. Осадки, накапливавшиеся на континентах, непрерывно перерабатывались потоками, пока суша не нивелировалась. Как только возникли растения, способные заселять сушу и собираться в достаточно густые поселения, начался регулируемый сток вод, появились настоящие почвы. Растения могли защищать склоны от непрерывного и быстрого размыва. Речные долины закладывались надолго, началось континентальное осадконакопление с образованием устойчивых, не подвергающихся переработке захоронений растительных остатков. Возможно, что именно это и произошло в начале зигенского века. Раз начавшись, этот процесс мог быстро захватить обширные участки суши.
Можно полагать, что в основном суша была заселена к концу девонского периода, поскольку в начале следующего, каменноугольного периода на Земле образовывались уже практически все основные типы осадков, ныне отлагающиеся на континентах. В самом начале карбона начинается образование угленосных внутриконтинентальных толщ. Само присутствие углей показывает, что на пути стока вод стояли мощные растительные фильтры. Не будь их, не могли бы образовываться угольные пласты, ибо остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, сильно обогащенные растительными остатками, — углистые сланцы или песчаники, а не настоящие угли. С этого времени Земля стала зеленой.
Если спросить далекого от палеонтологии человека, какие главные вопросы должны интересовать, по его мнению, специалиста по мезозойским рептилиям, то в ответ мы, может быть, услышим о вымирании динозавров. Ведь об этом то и дело пишут даже в газетах. Стало быть, специалисту тем более важно понять, почему вымерли динозавры в конце мезозойской эры, произошло ли это от похолодания или от вспышки сверхновой звезды. И тем не менее это не так. Специалисты по динозаврам в общем довольно равнодушны к происходящей дискуссии, в которой участвует кто угодно, но только не они (вернее, участвуют редко и высказываются, как правило, скупо). Все дело в том, что для исследователя, профессионально работающего с конкретным материалом, гипотеза приобретает интерес лишь тогда, когда она открывает путь к новым исследованиям, когда можно ее проверять наблюдением, а не заниматься умозрениями, вперив взор в потолок. По той же причине «снежный человек» больше интересует журналистов, чем антропологов.
Так и проблема заселения суши растениями и происхождения высших растений. Она долго была объектом умозрений, далеких от конкретных палеоботанических исследований. Поэтому палеоботаники проявляли к ней интерес только тогда, когда появлялись новые находки и их можно было изучать в лаборатории. Если надежды не оправдывались, интерес к этой проблеме опять исчезал и существовал лишь в среде людей, довольно далеких от палеоботаники. Пересмотр остатков якобы высших растений из додевонских отложений породил скепсис ко всем таким находкам. Повысились требования к качеству материала, детальности его изучения, точности датировок документов. Тем не менее сейчас начинает открываться новое поле исследований, причем не только для палеоботаников, но и для специалистов по осадочным породам. Палеоботаника должна идти рука об руку с палеоэкологией. На этой стадии особенно важно смотреть на проблему как можно шире, допуская к рассмотрению любые гипотезы, только бы они помогали осмысленному поиску новых фактов, помогали увязывать накопленные разнородные наблюдения. Здесь важно не спешить с выводами, не цепляться за одну гипотезу, а упорно и вдумчиво работать. Именно такой смысл — еще одной рабочей гипотезы — и имеют мысли, высказанные в этой главе о заселении суши растениями.
Глава II
ПЕРВЫЙ РАСКОЛ В РАСТИТЕЛЬНОМ ЦАРСТВЕ

Люди по-разному воспринимают прошлое. Одни следуют Екклесиасту: «Что было, то и будет; и что делалось, то и будет делаться, — и нет ничего нового под солнцем. Бывает нечто, о чем говорят: «смотри, вот это новое»; но это было уже в веках, бывших прежде нас». Однако стремление навязать прошлому черты наших дней часто терпит неудачу. Тогда начинает казаться, что раньше все было не так.
Остатки давно вымерших организмов производили большое впечатление на первых своих исследователей. Странным был облик животных и растений, да и встречались их остатки в необычных местах. Перед учеными постепенно открывался новый, неведомый органический мир. Остатки растений — листья, крупные стволы — попадались в высоких арктических широтах. В зоне вечной мерзлоты встречались кости мамонта, родство которого со слонами было совершенно очевидно еще в XVIII веке. Стало быть, климат на Земле когда-то был иным, более теплым и равномерным, чем сейчас. В прошлом веке было расшифровано ледниковое происхождение валунов, рассеянных на европейских равнинах. Значит, за теплыми временами настала эпоха грандиозного оледенения, остаток которого — нынешние арктические и антарктические льды.
На представления о прошлом Земли сильно влияли и космогонические гипотезы. Долго господствовало убеждение, что Земля, когда-то раскаленная, постепенно остывает. Внутренним теплом Земли были склонны объяснять заселение высоких широт богатой растительностью, от которой остались мощные пласты углей. Нынешняя картина климатических поясов рассматривалась как образование последних геологических эпох. Возникла проблема появления климатической зональности.
Некоторое непостоянство климатов в прошлом не вызывало сомнений у исследователей. Тем не менее было вовсе не обязательно видеть в следах оледенений или, наоборот, остатках южных растений в высоких широтах однонаправленное изменение климата. Гипотезе об остывании Земли был противопоставлен взгляд о постоянных климатических колебаниях. Его разделял знаменитый английский геолог Ч. Лайель.
С тех пор многое изменилось в представлениях об истории нашей планеты. Была выдвинута концепция холодного происхождения Земли и ее последующего разогрева. Стало ясно, что связывать климатические события с внутренним теплом недр рискованно. Тем не менее убеждение, что когда-то на Земле не было климатической зональности, удерживалось в умах многих исследователей. Оно поддерживалось и многочисленными фактами, которые свидетельствовали, что в местах, лежащих сейчас в совершенно различных климатических зонах, встречаются остатки одних и тех же организмов, сходных ассоциаций животных и растений.
Палеоботанические исследования начались в Европе в начале прошлого века и разворачивались довольно быстро. В 1804 г. Эрнст Фридрих фон Шлотгейм, живший в Тюрингии, впервые стал описывать ископаемые растения по образцу современных и присваивать им родовые и видовые латинские названия. В 1820 г. граф Каспар Штернберг, основатель и президент Национального музея в Праге, начал печатать выпуски капитального труда «Опыт геогностически-ботанического изложения флоры древних времен». Последний выпуск вышел в 1838 г., в год смерти Штернберга. В 1828–1837 гг. Адольф Броньяр, профессор ботаники в Париже, опубликовал замечательную сводку — «История ископаемых растений», к которой до сих пор обращаются палеоботаники. Штернберг и Броньяр не только дали систематическое описание всех известных тогда ископаемых растений, но и впервые установили закономерное изменение их ассоциаций по геологическому разрезу. История растительного мира была поделена ими на три этапа, примерно соответствующие палеозойской, мезозойской и кайнозойской эрам.
Естественно, что сначала в распоряжении палеоботаников были европейские коллекции. Из других частей света поступали лишь единичные случайные образцы. Тогда же в Европе складывалось и общее представление о геохронологии. Были выделены осадки разных геологических периодов и самим периодам были даны названия. К началу 1850-х годов завершилось составление геохронологической шкалы, удержавшейся в общих чертах до наших дней. К этой шкале можно было привязать и местонахождения ископаемых растений. В составлении шкалы принимали участие и палеоботаники, поскольку некоторые интервалы шкалы в Европе лучше охарактеризованы ископаемыми растениями, чем остатками животных.
Постепенно палеоботанические исследования распространились и на другие материки. Еще до середины прошлого века стали появляться описания коллекций, собранных в Сибири, Индии, Австралии, Северной Америке. Во второй половине века палеоботанические работы охватили все части света, кроме Антарктиды. Были открыты почти все наиболее известные местонахождения ископаемых растений. Правда, вопрос о возрасте многих местонахождений, расположенных далеко от Европы, отнюдь не всегда решался просто, о чем специально пойдет речь в следующих главах. Тем не менее к концу прошлого века сложилась некая общая картина эволюции растительного покрова Земли от девонского периода до наших дней.
Для наиболее молодых, кайнозойских флор устанавливалась ясная климатическая зональность, хотя и сильно отличающаяся от нынешней. На арктических островах были найдены листья деревьев, растущих сейчас в широколиственных лесах. Мезозойские флоры оказались значительно более однообразными по всей Земле. Одни и те же роды растений находили в Сибири, Англии, Индии. Сложнее было с палеозойскими флорами. Позднепалеозойские флоры Европы и Северной Америки выглядели примерно одинаковыми и в то же время отличались от флор южного полушария. Еще ниже по геологическому разрезу, в отложениях раннекаменноугольной эпохи, снова наступало единство флор разных материков. Едиными были и флоры предшествовавшего девонского периода.
Эти изменения типов флор во времени и пространстве были сопоставлены с климатическими условиями. Соответственно можно было описать изменение климатической зональности. Она была минимальной в девоне и начале карбона, затем усилилась в позднем палеозое, снова ослабла в мезозое, потом опять начала усиливаться, достигнув максимума в новейшие геологические времена.
В общем виде такая последовательность климатических событий признается специалистами и сейчас, однако есть и отличия нынешних взглядов от тех, что сложились в прошлом веке. История оказалась значительно сложнее, и расшифровка даже главных ее черт затянулась.
Начиная с 30-х годов стало разрушаться мнение о единстве мезозойских флор. Различия между юрскими флорами Сибири и Европы были подмечены еще в прошлом веке шведским палеоботаником О. Геером. Исследовав коллекции из Иркутского бассейна, он обратил внимание на большое количество в них листьев гинкговых. Для юрских флор Европы это не характерно. В 30-х годах А. Н. Криштофович, а затем В. Д. Принада и особенно В. А. Вахрамеев показали закономерное изменение комплексов юрских растений на территории Евразии. Выделилась самостоятельная Сибирская палеофлористическая область. Единство юрских флор мира оказалось кажущимся.
К сожалению, по опубликованной статье далеко не всегда можно понять, что навело автора на те или иные мысли, как он приходил к выводам, что послужило толчком к исследованию. Поэтому можно лишь гадать, насколько повлияли на специалистов по палеозойским флорам заключения о том, что мезозойские флоры не были однообразными по всей планете. Но показательно, что, как только укрепились взгляды о климатической зональности в юрском периоде, нашедшей отражение в распределении ископаемых флор, так появились сходные идеи и по отношению к флорам девона и раннего карбона, которые долго считались космополитными.
В 1955 г. в Ленинграде состоялась первая сессия Всесоюзного палеонтологического общества. Одним из докладчиков был ленинградский палеоботаник Г. П. Радченко. Показательно само заглавие его доклада: «К вопросу о первом проявлении ботанико-географической и климатической зональности в Северной Евразии». Чтобы оценить новизну идей Радченко, надо хотя бы кратко рассказать о господствовавших тогда взглядах. Всеми было признано, что в среднем-позднем карбоне и далее, в перми, существовало несколько палеофлористических областей. Из Северной Америки и Гренландии через Европу и Северную Африку в Среднюю Азию и далее на восток, до Китая и Юго-Восточной Азии, протягивалась огромная Еврамерийская область, населенная наиболее теплолюбивой, тропической флорой тех времен. В перми эта область разделилась. Ее восточная часть обособилась в Катазиатскую область тоже с тропическим климатом. К северу от этого тропического пояса, на материке Ангарида, располагалась Ангарская область умеренного климата, примерно соответствовавшая нынешней Северной Азии и временами захватывавшая крайний восток и северо-восток европейской части СССР. На противоположной стороне Земли располагалась Гондванская область. Она занимала материк Гондвана, объединявший Южную Африку, Южную Америку, полуостровную Индию, Австралию и Антарктиду. Гондванская флора тоже была умеренной или даже холодно-умеренной.
Во всех перечисленных местах есть и местонахождения раннекаменноугольных растений, но их состав казался удивительно однообразным. Голландский палеоботаник В. Йонгманс, один из создателей только что обрисованной картины областей позднего палеозоя, проанализировал раннекаменноугольные флоры Европы, Северной и Южной Америки, Австралии, Урала и других мест и пришел к выводу, что для выделения областей этого времени серьезных оснований нет. Хотя известное своеобразие можно отметить для каждого местонахождения, наметить какие-то регионально выдерживающиеся отличия местонахождений невозможно. Эту мысль высказывал еще в 1897 г. французский палеоботаник Р. Зейлер. В статье 1954 г. В. Йонгманс подтвердил ее.
Г. П. Радченко выступил с докладом через год и утверждал нечто совсем иное. Он убеждал слушателей, что отчетливые палеофлористические области можно установить с самого начала карбона. Расхождение во мнениях Йонгманса и Радченко можно понять. Йонгманс почти ничего не знал и не мог знать о раннекаменноугольных растениях Сибири. Эти растения были впервые описаны киевским палеоботаником И. Ф. Шмальгаузеном (отцом известного эволюциониста и. И. Шмальгаузена) еще в 70-х годах прошлого века. Потом отдельные небольшие коллекции описывали М. Д. Залесский, В. А. Хахлов и М. Ф. Нейбург. Однако сохранность остатков была довольно плохой, описания получались схематичными, и к тому же наиболее важным коллекциям были посвящены статьи, по-видимому оставшиеся не известными Йонгмансу. В этих статьях описывались некоторые новые, неизвестные в Европе роды и виды растений, но в целом как будто выявлялось сходство раннекаменноугольных флор Сибири и Европы.
В отличие от Йонгманса Радченко мог опереться не только на опубликованные в литературе списки латинских названий и несовершенные описания, но и на коллекции самих растений. В конце 40-х годов на всей территории нашей страны широко развернулись геологосъемочные работы. Была поставлена задача — получить достаточно детальные геологические карты страны. В палеонтологические лаборатории стали поступать многочисленные коллекции, собранные геологами для определения возраста пород. Такие коллекции поступали и к Радченко. Постепенно выяснялось, что отличия растений нижнего карбона Сибири и смежных районов от европейских не случайны. Радченко понимал, что для палеофлористического районирования недостаточно анализировать списки растений. Флористические области так или иначе отражают климатические условия, и поэтому важен не только списочный состав растений каждой области, но и климатически обусловленные особенности структуры растений. Радченко пришел к выводу: растения Сибири указывают на менее влажный и менее теплый климат, чем в Центральной Европе.
 Рис. 5. Предложенное Г. П. Радченко ботанико-географическое районирование Евразии в ранневизеиском веке; суша показана штриховкой; I — Средиземноморская область, II — Шотландско-Казахстанская область, III — Северо-Евразиатская область
Рис. 5. Предложенное Г. П. Радченко ботанико-географическое районирование Евразии в ранневизеиском веке; суша показана штриховкой; I — Средиземноморская область, II — Шотландско-Казахстанская область, III — Северо-Евразиатская область
Можно было ожидать в раннем карбоне примерно те же области, что и в остальной части палеозоя. Соответственно на территории СССР должны были бы выделяться Еврамерийская и Ангарская области раннего карбона. Однако Радченко пришел к другому выводу. На его карте (рис. 5) флористические границы пошли совсем иначе, чем на картах более поздних геологических эпох. Шпицберген, большая северная часть Гренландии, север европейской части СССР, половина Урала и вся Сибирь отошли к Северо-Евразиатской области. Далее к югу следовал пояс, протянувшийся от Шотландии через Прибалтику и Подмосковный бассейн, Средний и Южный Урал, Прибалхашье на Северо-Восток Китая. Это Шотландско-Казахстанская область. К югу от нее Радченко выделил Средиземноморскую область.
В 1965 г. началась подготовка к изданию целой серии аналогичных атласов для всех периодов. Я не знаю, чем руководствовались организаторы нового издания, но они решили составить новый атлас и для карбона. Предусматривалось включить в каждый выпуск карты биогеографического, в том числе и палеофлористического районирования. Я получил приглашение составить карты районирования флор карбона и принялся за дело. Проще всего было взять карты, уже подготовленные Г. П. Радченко, и представить их редколлегии. Однако ответственный за карбоновую часть атласа В. М. Познер предупредил такую возможность и сказал, что от меня ожидается иная схема районирования. Какие конкретно замечания были к схеме Радченко, я так и не узнал. Было только сказано, что климатические выводы Радченко недостаточно хорошо согласуются с результатами литологических исследований и выводами по другим группам организмов. Тогда же в нашей лаборатории палеофлористики Геологического института Академии наук СССР наметилась подготовка крупной сводки по фитогеографии всей Евразии в палеозое и мезозое. Палеозойская часть этой сводки так или иначе падала на меня. Поэтому в любом случае я должен был сам проработать накопившиеся данные по карбоновым (и пермским) флорам Евразии.
Человеку, далекому от палеоботаники, довольно трудно представить себе объем палеоботанической литературы. Наверное, ему будет странно узнать, что только палеозойским растениям и только Евразии посвящены несколько тысяч статей и десятки толстых монографий. Проработать всю эту литературу досконально просто невозможно. Принявшись за дело и начав с библиографической картотеки, составлявшейся моей наставницей М. Ф. Нейбург (она скончалась за несколько лет до этого), а затем мной, я быстро понял, что только для начала предстоит просмотреть как минимум полторы-две тысячи работ, большей частью на английском, французском и немецком языках. Пришлось еще раз с благодарностью вспомнить М. Ф. Нейбург, которая, когда принимала меня в ученики, поставила условием элементарной самостоятельности чтение литературы на этих языках без словаря. Мне пришлось выполнить это условие. Зато теперь я не испытывал ужаса перед горой литературы на разных языках.
Разумеется, прежде всего я тщательнейшим образом отпрепарировал статьи Радченко и Йонгманса, которые в моих глазах олицетворяли противоположные точки зрения. Я понимал, что со средним и поздним карбоном больших трудностей не будет, ибо здесь разногласия между разными исследователями были небольшими. Главная проблема — ранний карбон. Если прав Йонгманс, я должен признаться редколлегии атласа, что для этого времени никакого районирования дать вообще нельзя. Если же прав Радченко, то придется срисовывать его схему или создавать какую-то иную. Самым трудным в этой работе было найти правдоподобную рабочую гипотезу. Конечно, идеальным способом было бы просмотреть коллекции из разных мест, разобраться в систематике соответствующих растений и только потом приниматься за районирование. Но этот путь, кажущийся единственно разумным, был почти нереален: подавляющее большинство коллекций, послуживших для составления списков растений из того или иного места, недоступно.
Трудности возникают и с систематикой растений. Часто сохранность остатков недостаточно хороша, чтобы наблюдать все необходимые признаки.
Определения родов и видов становятся условными. Поэтому механическое собирание опубликованных списков растений и их формальный анализ не могли привести к успеху. Сами списки не слишком надежны.
Тем не менее ничего другого не оставалось. Я составлял многочисленные списки и складывал их в папки по основным регионам. Довольно долго работа была не очень осмысленной. Несколько обстоятельств изменили положение и привели к появлению желанной рабочей гипотезы. Я прочел статью по стратиграфии нижнего карбона Шпицбергена и в ней нашел упоминание старой статьи немецкого палеоботаника В. Готана, который в начале нынешнего века описал отсюда куски древесины. Самым важным было то, что в шпицбергенской древесине не было годичных колец (палеоботаники называют их «слоями прироста»). Факт очень важный. Обычно во внетропических флорах встречается древесина с ясными слоями прироста. Их нет только у деревьев, растущих в низких широтах, в условиях теплого климата с минимальными сезонными колебаниями. Однако древесина может быть занесена откуда угодно морскими течениями. Нужно было убедиться, что прочие растения Шпицбергена не противоречат заключению по древесине.
Выяснить ситуацию на Шпицбергене было особенно важно, ведь на схемах палеофлористического районирования, составленных для среднего и позднего карбона, Шпицберген попадает в тропическую Еврамерийскую область. На схеме же Радченко для раннего карбона он оказывается далеко за пределами тропиков. Если прав Радченко, надо допустить крупное перемещение границ палеофлористических областей на рубеже раннего и среднего карбона. Если же можно доверять древесине, описанной Готаном, то придется поставить под сомнение схему Радченко. Раз он неправильно интерпретировал флору Шпицбергена, то вполне мог ошибиться и с другими флорами.
 Рис. 6. Относительное количество древесины (зачернена) в маноксилическом (слева) и пикноксилическом стволах
Рис. 6. Относительное количество древесины (зачернена) в маноксилическом (слева) и пикноксилическом стволах
Подмога пришла сразу с трех сторон. Во-первых, вспомнилось, что растения, преобладающие в нижнем карбоне Шпицбергена (их Готан не описывал), принадлежат плауновидным с толстыми стволами. У плауновидных ствол сложен преимущественно корой, а древесины очень немного. Незадолго перед этим я слушал доклад специалиста по ископаемой древесине В. Г. Лепехиной, которая упомянула мнение ее учителя, крупного специалиста по анатомии растений А. А. Яценко-Хмелевского. Он считал, что стволы с таким строением, как у плауновидных (маноксилическим, что значит «малодревесинным»; см. рис. 6), если они многолетние, свидетельствуют о безморозном климате. Это хорошо видно у современных растений, хотя причина такой климатической приуроченности маноксилических стволов остается непонятной. Шпицбергенские плауновидные имели настолько толстые стволы, что считать их однолетними не приходится. Тогда можно сделать вывод, что климат Шпицбергена в раннем карбоне был безморозным. Заключение, сделанное по древесине, подтверждалось.
В статье двух индийских палеоботаников, Д. Бхарадваджа и Б. Венкатачала, изучавших споры и пыльцу из нижнего карбона Шпицбергена, основные роды и многие виды те же, что и описанные ранее из одновозрастных отложений Центральной Европы, Англии, Подмосковного бассейна. Помощь со стороны палинологии (науки, изучающей споры и пыльцу современных и ископаемых растений) была особенно важна, поскольку микроскопические оболочки спор и пыльцы гораздо чаще встречаются в породах, чем крупные остатки растений, и поскольку спорово-пыльцевые комплексы нижнего карбона изучались довольно интенсивно для целей нефтяной и угольной геологии. Обращение к палинологической литературе быстро убедило меня в том, что по спорам и пыльце нельзя относить юг, центр и север европейской части СССР к разным палеофлористическим областям. Вообще по всей Европе состав спор и пыльцы, хотя и обнаруживал некоторую специфику в том или ином районе, в целом был однообразен. Стало быть, и растения, продуцировавшие эти споры и пыльцу, были повсюду примерно одни и те же. Разделение Европы на три области не подтверждалось. Однако по этой причине еще нельзя было присоединяться к мнению Йонгманса. Надо было разобраться с Сибирью, Казахстаном и остальной частью Азии.
Проще всего оказалось с Центральным Казахстаном Еще в 30-х годах при изучении угленосного карбона Карагандинского бассейна были получены его палинологические комплексы По их составу можно не только определять возраст пород но и прослеживать угольные пласты на месторождениях. Палинологи С. Н. Наумова, А. А. Любер и И. Е. Вальц отметили своеобразие спор и пыльцы Карагандинского бассейна по сравнению с европейской частью СССР, но нашли и общие элементы между этими районами. Примерно тот же вывод позже сделали и специалисты по отпечаткам растений, предлагавшие выделять самостоятельную Казахстанскую провинцию. Можно было последовать этой идее.
Оставались Восточный Казахстан и Сибирь. Их флористическое единство сомнений не вызывало, но ясно сформулировать, чем флора этих районов отличалась от европейской, было трудно. Дело упиралось в недостаточно разработанную систематику растений нижнего карбона этих краев. Для одного и того же местонахождения разные палеоботаники давали настолько различные списки, что опираться ни на один из них было нельзя. Но и взяться за полный пересмотр систематики времени не было. Материалы должны были поступить в редколлегию атласа по возможности скорее, поскольку при составлении атласа не было запланировано переизучение первичного палеонтологического материала. Иначе работа затянулась бы на десятилетия.
Надо было найти какие-то такие особенности сибирско-казахстанских растений, которые можно легко проследить и которые бы давали наиболее надежные отличия от растений других районов. Что требовалось для решения проблемы, я понял уже потом, а само решение пришло благодаря счастливому случаю. Примерно за год до того, как я ломал голову над нижнекарбоновыми растениями Сибири, мне принесли образец из нижнего карбона Южного Урала и попросили определить возраст. Это был кусок корненосца плауновидного растения. Такие корненосцы называют «стигмариями». Они встречаются в Европе почти по всему разрезу карбона и заходят в нижнюю пермь. Поэтому мне пришлось разочаровать человека, принесшего образец, и сказать, что возраст можно определить лишь в таких широких пределах. Сам образец был довольно плохим, а растение — довольно обычным для европейской части Союза. Я не стал его регистрировать, но и не выкинул, а бросил в ящик стола до лучших времен вместе с различными инструментами. Там он и валялся, попадаясь каждый раз на глаза, когда я лез за молотком или зубилом.
Выдвинув однажды в очередной раз ящик и снова увидев этот образец, я внезапно сообразил, что это самое обычное для Европы растение отсутствует в наших коллекциях из нижнего карбона Сибири. Оно достаточно характерно, чтобы его не заметить или неправильно определить. Слои со стигмариями хорошо знают все геологи, работающие в Донбассе или в любом ином палеозойском угольном бассейне Европы. Их неизменно отмечают при описании геологического разреза, даже не обращаясь за помощью к палеоботаникам. Махнув рукой на остальную работу, я стал перелистывать списки сибирских растений и описания геологических разрезов, в которых были встречены в Сибири и Восточном Казахстане остатки плауновидных. Указаний на стигмарии нигде не было. Когда я нанес на карту все известные местонахождения раннекарбоновых стигмарий, у меня получился пояс, совпадающий с Еврамерийской областью среднего-позднего карбона, а свободным от точек местонахождений оказалось большое пятно, совпадающее с Ангарской областью (рис. 7).
Говоря о стигмариях, важно помнить, что это не какой-то отдельный вымерший род. Родовое название Stigmaria не применяется к целому реконструированному растению. В палеоботанике вообще редко приходится сталкиваться с описаниями целых растений. Обычно в распоряжении палеоботаника оказываются изолированные листья, семена, стебли, корни и другие части растения, прижизненное сочетание которых неизвестно или гипотетично. Поэтому все эти части описываются под разными родовыми названиями. Так поступают даже в тех случаях, когда обнаруживается прижизненное сочетание частей. Эта осторожность необходима. Ведь всегда может оказаться, что часть данного облика сочеталась у разных растений с разными по облику прочими частями. Именно так происходит со стигмариями. Сходные по облику корненосцы имели плауновидные разных родов — лепидодендроны, сигиллярии и др. Таким образом, присутствие стигмарии указывает на распространение целой группы растений. Но и не это самое важное. Стигмарии особенно характерны для плауновидных, населявших приливно-отливную зону морских побережий. Эта зона сейчас занята в тропических странах мангровыми зарослями (о них еще пойдет речь в этой книге). Отсутствие стигмарий в Сибири навело на мысль, позже подтвердившуюся, что здесь приливно-отливная зона морских побережий не была заселена растительностью, как было в Еврамерийской области.
 Рис. 7. Стигмарии и их аппендиксы (видны в два ряда на левом рисунке) плауновидных карбона; на карте показано распространение стигмарии в визейских отложениях Евразии, они есть в Еврамерийской области (Е) и Казахстанской провинции (K), но отсутствуют в Ангарской области (А)
Рис. 7. Стигмарии и их аппендиксы (видны в два ряда на левом рисунке) плауновидных карбона; на карте показано распространение стигмарии в визейских отложениях Евразии, они есть в Еврамерийской области (Е) и Казахстанской провинции (K), но отсутствуют в Ангарской области (А)
Еще до этого С. Г. Горелова изучила закономерности размещения различных растительных остатков в разных породах нижнего карбона Кузбасса и выяснила, что остатки плауновидных обычно приурочены к более грубозернистым породам — песчаникам, тогда как растения с листвой папоротникового типа более характерны для тонкозернистых пород. Ничего подобного нет в Европе. Здесь тоже есть определенные зависимости состава растений от разностей пород, но они совсем иные.
Сопоставление всех этих и других фактов привело к выводу: как Радченко, так и Йонгманс были и правы, и не правы. Йонгманс был прав в том, что усмотрел существенное единство раннекаменноугольных флор на месте Еврамерийской области среднего и позднего карбона. Но он неправильно заключил, что в раннем карбоне вообще не было флористических областей, контролируемых климатом. Г. П. Радченко справедливо отметил своеобразие сибирской флоры раннего карбона, но ошибочно отождествил с ней флору Шпицбергена. Не подтвердилось его деление Европы на три самостоятельные области. Правда, более поздние исследования раннекаменноугольной флоры Европы, Северной Америки и Северной Африки показали, что и здесь можно выделить более дробные флористические районы, но их разница невелика и к тому же их границы не те, что нарисовал на своей схеме Радченко.
Таким образом, схема палеофлористического районирования, давно составленная для среднего и позднего карбона, оказалась в общих чертах применимой и для раннего карбона. Естественно, что тут же возник вопрос: нельзя ли эту схему протянуть и в предшествующие геологические эпохи, принадлежащие уже девонскому периоду?
Возможность того, что флоры девона не были вполне однообразными по всей Земле, допускалась палеоботаниками еще в 30-х годах. В 60-х годах сначала чешский палеоботаник И. Обргел, а затем Н. М. Петросян, ученица Г. П. Радченко, сделали попытку предложить схему палеофлористического районирования девона. При этом Н. М. Петросян предложила выделять палеофлористические области даже для раннего девона. Но отношение других палеоботаников к этим попыткам до сих пор довольно прохладное. К тому же предложенные схемы не согласуются друг с другом. Возможно, это происходит из-за того, что сами схемы строились путем анализа списочного состава многочисленных частных флор. При этом можно было класть в основу районирования самые разные растения. Не выявив какие-то четкие флористические тенденции, отражающие климатические различия территорий, едва ли можно достичь согласия между схемами районирования. В 1973 г. Диана Эдварде, работающая в Кардиффе, попыталась обобщить данные по девонским флорам всего мира и нанести их на карту. Выводы не были обнадеживающими. Единственное, что ей удалось сделать, — это подметить некоторую специфичность флор южного полушария, но и здесь пока рано говорить о четких палеофлористических областях.
Подводя итог многолетним попыткам найти палеофлористические области в девоне, приходится ограничиться следующими наметками. Для всего девона можно отметить лишь одну достаточно четкую закономерность. Имеется в виду распределение, толстоствольных плауновидных, которые полностью отсутствуют: в девоне Сибири и не характерны для Европы, но зато часто встречаются в Центральном Казахстане, на Шпицбергене и в Северной Америке. Распространение этих растений намечает некий пояс, совпадающий с северной периферией будущей Еврамерийской области карбона. В остальном приходится признаться, что девон во всяком случае не был временем отчетливой географической дифференциации растительного покрова. Этот вывод, кстати, подтверждается и палинологическими данными. Ю. С. Надлеру и Е. В. Чибриковой удалось прямо сопоставить девонские палинологические комплексы Кузбасса и европейской части СССР, т. е. мест, флоры которых в карбоне были совершенно разными. Н. Г. Пашкевич провела такое же сопоставление между Якутией и Тиманом, причем тоже успешное. Значительная общность палинологических комплексов известна между Европой и Австралией. Многие роды, установленные по спорам, имеют практически всесветное распространение. Их находят всюду, где вообще удается получить палинологический комплекс подходящего возраста.
Впрочем, и по отпечаткам растений устанавливается очень широкое распространение некоторых родов и даже видов. Род Archaeopteris известен в Западной Европе, на Земле Элсмира, в штате Нью-Йорк, на Медвежьем острове, на Тимане, в Донбассе, в Минусинском бассейне и в других местах. Такого широкого распространения одного рода в карбоне мы не знаем. Примечательно, что и в морской фауне девона географические области выделяются недостаточно четко. Отмечались провинциальные различия морских фаун раннего и среднего девона, которые сильно сглаживаются в позднем девоне. Как раз к среднему девону относится уже упомянутая закономерность в распределении толстоствольных плауновидных, а широкое распространение рода Arhaeopteris падает на поздний девон. Если это совпадение с палеозоологическими наблюдениями не случайно, то можно ожидать возрастания географических различий между флорами раннего девона. Однако этого не получается. Некоторые раннедевонские роды распространены чуть ли не на всех материках.
С чем связано расхождение палеозоологических и палеоботанических данных в отношении биогеографической картины раннего девона, сказать трудно. Наиболее правдоподобным кажется такое объяснение. Чтобы получилась достаточно отчетливая картина палеофлористического районирования, надо обнаружить зависимость распределения растений от климата. Однако раннедевонские растения, видимо, были преимущественно водными. Кроме того, судя по их величине и анатомическому строению, большая их часть принадлежала к однолетникам. Распространение таких растений — водных или полуводных однолетников — могло в минимальной степени зависеть от климатических условий. Вода спасала их от сухости, жары и холода, а неблагоприятный сезон они могли пережить в виде спор или заростков. Может быть, однолетними были только их воздушные части, а стелющиеся по грунту побеги жили дольше. Для таких растений тем более не было проблемы перенести неблагоприятный сезон, ведь не случайно современный тростник живет и в дельте Ганга, и в устье Северной Двины.
Глава III
«ВЫМЕРШИЙ» КЛИМАТ

Я не знаю, проследил ли кто-нибудь из историков науки, как появились в нашем языке выражения «вымершие животные», «вымершие растения». Мы так свыклись с этими словами, что они кажутся непременной частью нашего языка.
К тому, что кажется непреходящим, эпитет «вымерший» мы, конечно же, не применяем. Мы не говорим «вымерший песчаник» или «вымершая температура». Слово «вымерший» не подходит и к единичным объектам. Про гору, которая когда-то стояла, а затем была размыта, мы скажем «исчезнувшая». «Вымершими» же мы называем классы объектов. Песчаник как класс горных пород не вымирал. А что вымирало, кроме групп животных и организмов? О чем еще можно говорить как о вымершем? Это один из сложнейших вопросов, стоящих перед науками о Земле. Есть некоторые типы пород, образовывавшихся лишь в очень далеком прошлом. Таковы джеспилиты — тонкослоистые железистые кварциты, огромными массами накапливавшиеся в докембрии. Почти «вымерли» доломитовые илы, которые сейчас встречаются лишь в немногих водоемах, а раньше осаждались в обширных морских бассейнах.
Продолжить этот список вымерших или вымирающих классов объектов, если оставить в стороне организмы, нелегко. Думаю, что к составлению такого списка не готов ни один геолог. А выполнить эту работу стоило бы. Пора как следует задуматься над тем, какие классы объектов неживой природы исчезли на Земле безвозвратно или по крайней мере не окружают нас сейчас, но, может быть, восстановятся в будущем. Важное место в таком списке, наверное, займут «вымершие» климаты.
Читая работы о климатах далекого прошлого, обычно встречаешь характеристики, заимствованные из климатической картины современной Земли. Говорится о тропическом, субтропическом, умеренном климате, проводятся сравнения древних и нынешних ландшафтов. В сознание ученых с трудом проникает представление, что в прошлом могли существовать климаты и соответствующие им ландшафты, даже близкого подобия которых нет на Земле сейчас.
И все же мысли о необычных климатических обстановках геологического прошлого возникали. Они пришли в голову и мне, когда я занимался историей флор Евразии в каменноугольном периоде и рисовал карты палеофлористических областей.
Необходимое условие этой работы — установление возрастных соотношений разных флористических комплексов, т. е. решение проблем стратиграфии. Ясно, что если мы хотим составить карту Российского государства в XVIII в., то надо брать и документы этого времени. То, что в причерноморских степях есть древние скифские курганы, важно для понимания русской истории, но ничего не скажет нам о том, где надо провести южную границу России в петровские времена.
Для эпох, когда климатическая зональность была ослаблена и виды животных и растений беспрепятственно расселялись по морям и континентам, проблемы
синхронизации отложений решаются довольно легко. Чем более контрастны климаты Земли, тем ярче региональные особенности фаун и флор, тем труднее сопоставление отложений разных регионов. Именно с этой трудностью и приходится сталкиваться, когда мы сопоставляем флоры разных мест планеты, существовавшие после первого раскола в растительном царстве.
Эта трудность впервые всплыла перед исследователями каменноугольных и пермских толщ Сибири в предреволюционные годы, когда началось систематическое изучение и освоение Кузбасса. Тогда уже было известно, что под угленосной толщей Кузбасса лежат безугольные песчано-глинистые слои с растительными остатками, позже названные острогской свитой. Ниже лежит морская толща, преимущественно сложенная известняками с раннекаменноугольной фауной. О возрасте острогской свиты разгорелись споры. Правда, в ней тоже нашли прослой с морской фауной, но ее возраст палеозоологи определяли по-разному: от раннего карбона до ранней перми включительно — разногласие на целый период, невыносимое для стратиграфов. Палеоботаники были более единодушны и считали острогские растения раннекарбоновыми, но постепенно они приходили к выводу, что в Кузбассе нет европейских родов и видов, а поэтому и сопоставление флор Кузбасса и Европы стало восприниматься с недоверием, причем совершенно справедливым.
В послевоенные годы морская фауна, сходная с острогской, была обнаружена на Таймыре и на северо-востоке СССР, но ее датировка вызвала те же споры. Вопрос о возрасте острогской свиты стал притчей во языцех, и им даже попрекали палеонтологов, которые-де переоценивают роль палеонтологии в стратиграфии. Только в 60-х годах появилась возможность прямо сопоставить некоторые арктические разрезы Азии с европейскими по остаткам аммоноидей (вымерших головоногих моллюсков) и сузить диапазон разногласий. Тогда же всплыли и некоторые палеоботанические данные, на которые раньше не обращали внимания.
В главе II мельком упоминались растения с маноксилическими стволами, которые, если они многолетние, свидетельствуют о безморозном климате. Еще в 30-х годах М. Ф. Нейбург, обобщая данные по позднепалеозойской флоре Кузбасса, выделила в ее истории два главных этапа. На первом господствовали плауновидные растения, а на втором — кордаиты, вымершие голосеменные растения. Плауновидные характерны для нижней части острогской свиты, а выше они довольно быстро исчезают и вскоре начинают появляться листья кордаитов, к которым постепенно переходит главная роль среди растительных остатков, особенно в угленосных отложениях. Такая же закономерность потом обнаружилась и в других частях Ангариды.
У острогских плауновидных (фиг. V) не было таких толстых стволов, как у их европейских родственников (в Европе попадались стволы около метра в диаметре), но все же они были слишком толстыми, чтобы считать их однолетними. Сомнений в том, что эти стволы из Кузбасса, а также из Тунгусского и Минусинского бассейнов, Северо-Восточного Казахстана, Верхоянья, Магаданской области были маноксилическими, не возникало. Поэтому, как и в случае с раннекаменноугольными плауновидными Шпицбергена, фигурировавшими в прошлой главе, можно было считать острогские растения свидетелями безморозного климата. Раннекарбоновая Ангарида, оказывается, не знала морозов, по крайней мере в низинах, заселенных известными нам растениями.
С другой стороны, все данные об условиях формирования вышележащих угленосных отложений Ангариды свидетельствовали о климате скорее всего тепло-умеренном или даже умеренном. Напрашивалась гипотеза, что намеченные Нейбург этапы имеют палеоклиматическую подоплеку. Исчезновение толстоствольных плауновидных на огромной территории Ангариды, занимавшей не менее 10 млн. кв. км, наводило на мысль о быстром и сильном похолодании. Столь большие перемены не могли пройти незаметно и для соседних областей. Поэтому я обратился к европейским материалам, и здесь мое внимание сразу привлекло явление, подмеченное еще в прошлом веке. Англичанин Р. Кидстон назвал его флористическим разрывом, а немецкий палеоботаник В. Готан — флористическим скачком.
В Европе, как и в Кузбассе, в нижней части карбона преобладают морские толщи, перекрываемые угленосными отложениями. Соответственно европейские геологи выделяют в карбоне два отдела — нижний карбон, или динант, и верхний карбон, именуемый силезием. Нижний член силезия называется намюрским ярусом. В нем еще много морских осадков, и облик его переходный между двумя отделами.
Выделение динанта и силезия опиралось на историю европейского осадконакопления. Если бы стратиграфы уделили больше внимания палеоботаническим данным, они провели бы границу между отделами карбона иначе. Именно на это и указал Р. Кидстон. Он писал, что растения в нижней части намюра имеют еще динантский облик и что палеоботаническая граница проходит внутри намюра. Эта граница отмечена довольно серьезным изменением состава растительных остатков. Еще важнее другое: ведь изменяется не только родовой и видовой состав флоры, но и ее общее разнообразие. Поэтому польский палеоботаник 3. Стопа предложил говорить не о «разрыве» и не о «скачке», а об «обеднении» флоры. Например, в намюрских отложениях Бельгии, флора которых изучена особенно хорошо, количество видов сокращается вдвое (почти с двухсот видов примерно до сотни) и восстанавливается лишь в верхней части намюрского яруса.
Кажется логичным предположить, что намюрское «обеднение» флоры Европы — отклик на ангарское похолодание. Тогда открывалась возможность сопоставлять разрезы Сибири и Европы не по спискам видов и родов, а по климатической пере стройке, одной и той же по всей Евразии. Всеми этими соображениями я поделился с палеозоологами, выступил с докладами и статьей. Нельзя было строить иллюзий, что эта идея будет сразу принята, поскольку слишком давно шла дискуссия и трудно ожидать ее немедленного прекращения. И тем не менее отношение к мысли о намюрском похолодании было сочувственным. Не вызвало возражений и сопоставление нижней части острогской свиты, содержащей толстоствольные лепидофиты, с нижней частью намюрского яруса Европы. Тут подоспели некоторые новые данные о морских фаунах Северной Азии. Они согласовывались с палеоботаническим заключением. Дискуссия о возрасте острогской свиты и сопоставляемых с нею отложений быстро и незаметно утихла.
Когда осталась позади стратиграфическая проблема в самой трудной ее части, можно было поразмыслить о том, что же получилось. А получилось нечто весьма своеобразное. Палеонтолог-стратиграф В. Г. Ганелин прислал мне большую коллекцию раннекарбоновых плауновидных, собранных в бассейне реки Омолон. Поступили коллекции примерно того же возраста из Верхоянья и из других мест Восточной Сибири. Еще раньше M. Ф. Нейбург получила прекрасные коллекции плауновидных из острогской свиты Кузбасса. Занявшись вплотную этими коллекциями, я только поражался сходству растительных остатков из всех этих мест.
Чтобы читатель понял мое удивление, стоит напомнить, что коллекции с северо-востока СССР были собраны не так уж далеко от района, где помещался северный полюс того времени. Положение полюса было определено еще до моих исследований по совокупности литологических и фаунистических данных. Полагают, что полюс находился где-то в районе низовьев Лены. Правда, по палеомагнитным данным приходится его загонять дальше на восток, в Тихий океан, но так получается только в том случае, если допустить, что материки всегда сидели на нынешних местах, а диаметр Земли не менялся. Оба этих допущения, как мы еще увидим, по меньшей мере необязательны.
Впрочем, как бы мы ни оценивали положение полюса в раннем карбоне, он все равно попадает в пределы Ангариды или омывавших ее с севера морей. Так или иначе остатки плауновидных острогского и более древнего типа оказываются в очень высоких широтах. В таком-то месте и надо допускать былое существование безморозного климата. Но ведь солнце тогда ходило по небу так же, как и сейчас. В высоких широтах оно на зимние месяцы скрывалось за горизонтом. Наступала теплая арктическая ночь. В этой темноте и стояли заросли плауновидных.
Странный ландшафт! Трудно представить себе его. Палеоботаники привыкли к облику еврамерийских плауновидных — лепидодендронов. Это были деревья с раскидистой кроной. Арктические ангарские плауновидные выглядели совсем иначе (рис. 8). Через мои руки прошли тысячи экземпляров ангарских плауновидных, но лишь в редких случаях, да и то у немногих видов, мне удалось наблюдать ветвящиеся стебли. Обычно видишь прямые палки без ответвлений и веточных рубцов. У одних родов и видов палки потолще, у других — потоньше. В общем, чем южнее, тем все толще стволы. В коллекциях из Кузнецкого, Минусинского и Тунгусского бассейнов они до 20–30 см в диаметре. На северо-востоке ствол толщиной 10 см уже редкость. Обычно побеги более тонкие. Но и 10 см в диаметре позволяют предположить, что высота ствола была не меньше 2 м.
 Рис. 8. Реконструкция томиодендрона (слева) и лофиодендрона (Lophiodendron) из раннего карбона Ангариды
Рис. 8. Реконструкция томиодендрона (слева) и лофиодендрона (Lophiodendron) из раннего карбона Ангариды
У еврамерийских плауновидных отношение только неразветвленной части ствола к его толщине у комля составляет от десяти до тридцати к одному. Судя по имеющимся фрагментам, стволы ангарских плауновидных сужались вверх очень постепенно, так что отношение длины ствола к диаметру было не менее двадцати к одному. По листовым подушкам на стволах можно судить о динамике роста ствола. В отдельных его участках подушки сильно сближены и как бы стиснуты, а выше и ниже расположены более свободно. Возможно, так отразились на росте ствола сезоны года или периоды размножения. Сопоставляя эти и некоторые другие факты, можно заключить, что стволы беспрепятственно росли несколько лет. Получалась длинная неветвящаяся палка с пучком листьев наверху и ежиком отсохших листьев ниже по стволу. Можно попытаться представить себе этот странный ландшафт. По берегам рек и озер тянется унылая щетка из палок разной величины. Некоторые завалились. Вода подхватывает их и несет, сбивает кучами в заводях. Местами щетка прерывается зарослями папоротниковидных растений с округлыми листьями-перышками. Остатки таких листьев, еще сидящих на побегах, обычно встречаются в раннекаменноугольных толщах Сибири. Осеннего листопада, наверное, еще не было. Вместе с этими растениями никогда не встретишь ни косточки какого-нибудь четвероногого, ни крылышка насекомого. Тихо было в зарослях.
Когда мы однажды обсуждали эти странные ландшафты с М. И. Грайзером, много лет изучавшим нижнекаменноугольные отложения Сибири, он обратил мое внимание еще на одно обстоятельство. В раннем карбоне Ангариды, считает он, было сухо, и не только на юге — в Минусинском бассейне или в Кузбассе, но и на севере. В бассейне реки Оленек в отложениях низов карбона есть гипсоносные толщи, которые, по всем литологическим канонам, могли откладываться только в сухом (аридном) климате. Можно предположить, что те растения, остатки которых мы находим в немалом количестве в нижнем карбоне Ангариды, селились поближе к воде, а возвышенные места были голыми. Об этом косвенно свидетельствует и тот факт, что даже там, где скапливалось особенно много растительных остатков, дело не доходило до образования угольных пластов, нет даже тонких прослоев угля. С голых возвышенностей, надо думать, непрерывно сносились песок и глина. Накопление же угля возможно лишь тогда, когда песчаные и глинистые частицы не подмешиваются в большом количестве к скоплению растительных остатков.
Здесь надо заметить, что отсутствие угленакопления не было особенностью раннекаменноугольной эпохи вообще. В экваториальной области угленакопление тогда уже шло вовсю, скажем, на Урале, в Подмосковном бассейне, на Шпицбергене и в других местах, хотя главные эпохи образования углей в экваториальной области наступят позже. Появление угленосных толщ в раннем карбоне, причем не только по побережьям морей; но и во внутриконтинентальных впадинах, — важное свидетельство того, что растительность покрывала склоны и сдерживала их размывание, препятствуя сносу песчано-глинистого материала на дно впадин. Только поэтому в болотах на дне впадин могли накапливаться угольные пласты.
В классификации современных климатов не заготовлена ячейка, куда можно поместить условия раннего карбона на Ангарском материке. Поэтому вполне можно говорить о том, что это был по-настоящему «вымерший» климат. Для него еще надо придумать название. Можно сказать, например, так: высокоширотный аридно-безморозный климат. Но лучше, может быть, не обращаться к такой терминологии, поскольку над свойствами этого «вымершего» климата еще предстоит поломать голову. Поэтому лучше воспользоваться другим способом обозначения климатов. Им широко пользовался В. Кеппен, один из крупнейших климатологов нашего столетия. Он говорил о «климате березы» или о «климате фуксии», и это не было стремлением к одной лишь образности языка. Кеппен понимал, что, классифицируя климаты по растениям и растительности, мы сразу получаем легко понимаемые и четко отграничиваемые типы. В такой классификации автоматически учитываются вариации климата по годам, поскольку за долгий срок существования рода растений или определенного типа растительности характеристики климата как бы осредняются. Тем самым отделяются климаты с определенной амплитудой в температуре, влажности и других показателях.
В. А. Красилов использовал тот же прием для климатов геологического прошлого и предложил называть климат Еврамерийской области карбона климатом стигмарии. К сожалению, и этот прием, хотя и наиболее перспективный, пока нельзя использовать для обозначения вымершего ангарского климата. Надо полнее узнать соответствующие роды растений, как следует изучить их распределение в разрезе и на площади. Возможно, что в такой роли в будущем выступят роды Tomiodendron или Lophiodendron, широко распространенные в нижнем карбоне Ангариды.
После всего сказанного об ангарском «вымершем» климате у читателя могут возникнуть вопросы: почему вообще был возможен такой климат, есть ли другие факты, подтверждающие палеоботанические свидетельства, почему произошло крупное похолодание в конце раннего карбона, как отразилось это похолодание по другую сторону от Еврамерийской области, на гондванских материках, и был ли там до похолодания тоже безморозный климат? Все эти вполне естественные и действительно важные вопросы еще ждут обстоятельного ответа. Сейчас можно предложить вместо ответа лишь отдельные соображения, во многом гипотетические.
Прежде всего надо учесть, что безморозный климат в высоких широтах — далеко не уникальная черта раннего карбона. Судя по палеонтологическим свидетельствам, морозные зимы захватывали полярные области лишь в отдельные эпохи документированной истории Земли. Поэтому современная климатическая картина с крупными полярными шапками, покрытыми снегом и льдом, скорее исключение, чем правило в геологической истории. Это значит, что мы должны искать ответа не на вопрос, почему в раннем карбоне или какой-либо иной эпохе не было у Земли ледниковых шапок, а на вопрос, отчего они иногда образовывались и меняли весь климат планеты.
Для последнего оледенения, начавшегося еще в неогеновом периоде, покрывавшего в четвертичном периоде (антропогене) огромные площади в северном и южном полушариях и еще не закончившегося, вопрос так и ставится. Над причинами этого оледенения ученые ломают голову более столетия, так и не достигнув единодушия.
Очевидно, к причинам оледенений надо подбираться не изолированно для каждой эпохи, а сопоставляя черты таких эпох. Кроме того, разумно относиться к оледенениям не как к каким-то посторонним событиям, накладывающимся на рутинную климатическую историю планеты, а как к естественным и закономерным фазам этой истории, ее крайним, а не экстраординарным случаям. Приняв эти установки, можно и оценить ранее выдвигавшиеся гипотезы, и наметить пути дальнейших исследований.
В изучении дочетвертичных климатов, начавшемся еще в прошлом веке, центральное место долго занимали броские климатические события — эпохи оледенений, иссушений климата, сильных потеплений. Картина получалась драматичной, и причины климатических переворотов хотелось искать в необычных происшествиях. Приходили в голову космические катастрофы, путешествие Земли через облака космической пыли, отделение или удаление Луны и пр. В последние десятилетия внимание ученых привлекли сравнительно небольшие климатические колебания. Это изменение интересов можно сравнить с тем, что произошло в истории. Когда-то историки интересовались главным образом войнами, революциями, дворцовыми переворотами и в этом видели основной смысл исторического исследования. Лишь много позже они обратились к повседневной экономической жизни народов. Исследования малозаметных событий гораздо более трудоемки, но они восстанавливают прошлое в его полноте, а не в виде набора единичных впечатляющих событий. Расшифровка повседневной жизни позволяет понять и крупные происшествия.
Существует немало способов реконструировать климатическую обстановку прошлого. Используются признаки осадочных пород и их сочетаний, состав остатков организмов в породах. По изотопному составу кислорода в раковинах можно восстановить температуру воды, в которой жило животное. Еще в начале века было установлено, что у цветковых растений листья с ровным краем чаще встречаются в тропиках, чем в умеренных широтах. Листья с таким же краем, но мелкие характерны для сухого климата.
Благодаря привлечению этих и других показателей климата, с которыми нам еще предстоит столкнуться в последующих главах, удалось выявить непрерывные перемены в климатических условиях в последние геологические периоды. Потепления сменялись похолоданиями, более влажные условия — сухими, сужались и расширялись климатические зоны. Вместо однонаправленных тенденций, гипотетически выдвигавшихся исследователями раньше (утверждали, например, что климат всей Земли становится все суше или все холоднее), стала вскрываться сложная, но небеспорядочная картина.
 Рис. 9. Палеоклиматические кривые для разных районов Земли и отраженные в изменениях разных признаков: а — палеотемпературы поверхностных вод Карибского моря; б — изменение уровня океана под влиянием таяния ледников, установленное по коралловым террасам острова Барбадос; в — палеоклиматическая кривая Европейской части СССР по комплексу данных; г — палеоклиматические колебания, установленные по составу планктонных фораминифер западнее Ирландии; д — процент пыльцы древесных растений в осадках одного из озер Македонии; е — палеотемпературные кривые по изотопному составу кислорода в толще льда Гренландии (по X. А. Арсланову, Н. В. Кинд и др.)
Рис. 9. Палеоклиматические кривые для разных районов Земли и отраженные в изменениях разных признаков: а — палеотемпературы поверхностных вод Карибского моря; б — изменение уровня океана под влиянием таяния ледников, установленное по коралловым террасам острова Барбадос; в — палеоклиматическая кривая Европейской части СССР по комплексу данных; г — палеоклиматические колебания, установленные по составу планктонных фораминифер западнее Ирландии; д — процент пыльцы древесных растений в осадках одного из озер Македонии; е — палеотемпературные кривые по изотопному составу кислорода в толще льда Гренландии (по X. А. Арсланову, Н. В. Кинд и др.)
Наиболее важная черта этой картины — высокая степень синхронности климатических событий в разных частях планеты, глобальный характер не только крупных перестроек, но и небольших перемен. Сначала эта интереснейшая закономерность была установлена для конца четвертичного периода, т. е. для последних десятков тысяч лет. Большую роль здесь сыграли определения возраста отложений с помощью изотопов углерода в органических остатках (радиоуглеродный метод). Еще до этого по остаткам спор и пыльцы во многих районах Земли были установлены небольшие климатические колебания, отразившиеся в смещении растительных зон и осадконакоплении. Эти колебания считали чем-то местным и соответствующие климатические эпизоды в европейской части СССР и, скажем, в Северной Америке называли по-разному. Радиоуглеродные исследования убедительно показали высокую степень синхронности эпизодов на разных материках — в Европе, Америке, Австралии, Африке, Азии (рис. 9).
Эти данные о постоянных климатических переменах и о синхронности похолоданий и потеплений в разных частях планеты заставляют с сомнением относиться к гипотезам, объясняющим оледенения космическими причинами, например, такой, как прохождение Солнечной системы через облако космической пыли. Хотя роль некоторых космических факторов (например, изменение параметров земной орбиты) отрицать рискованно, но едва ли именно космос был непосредственной причиной климатических перемен в истории Земли. Более вероятно допущение, что климат менялся под влиянием палеогеографических перестроек, воздействовавших на циркуляцию атмосферных и водных масс. При этом изменения в одном месте провоцировали отклик в других местах в соответствии с афоризмом «Щелкни кобылу в нос, она махнет хвостом». Некоторые точки Земли особенно чувствительны, и перемены в них могут иметь особенно серьезные последствия.
Сейчас предпринимаются попытки реконструировать климаты далекого прошлого, построив палеогеографические карты и рассчитав циркуляцию течений и атмосферных масс. В нашей стране такую работу недавно предприняли С. С. Зилитинкевич, Д. Д. Квасов и А. С. Монин. Вот что они писали в 1976 г. на страницах журнала «Природа»: «Проведенные расчеты показали, что изменения климата, происходившие в геологическом прошлом, могут быть объяснены «земными» причинами… Главной из них является зависимость климата от изменений расположения суши и моря, а также глубин океана и рельефа суши. Труднее всего объяснить с этой точки зрения быстрые и резкие колебания климата, происходившие в ходе чередования ледниковых и межледниковых эпох на протяжении последних 700 тыс. лет. За это время не произошло существенного перемещения материков. Но, по нашему мнению, сами ледниковые щиты были теми формами рельефа, которые существенно изменяли облик суши. Снижение уровня океана приводило к уменьшению его глубин, а также к осушению шельфов, что значительно изменяло расположение суши и моря. Во время межледниковий оно было благоприятным для начала оледенения умеренных широт, а когда последнее достигало больших размеров, становилось иным — неблагоприятным для его дальнейшего существования. Расчеты изменений климата во время четвертичных оледенений предстоит еще провести в будущем».
В этой цитате важно не пропустить слово «расчеты». Современная климатология — не та наука, где можно строить гипотезы, не прибегая к математическим методам, к расчетам течений, температур и многого другого. «Качественные» (в противовес «количественным») гипотезы сохраняют свой интерес на первых стадиях разработки проблемы, но без математической опоры уже не могут претендовать на истинность. К сожалению, эти расчеты очень сложны просто с математической точки зрения. Впрочем, главная трудность даже не в этом. К расчетам можно приступать, только получив карту распределения материков и океанов и имея представление о рельефе суши и глубинах морей. Хотя при составлении этих карт математика тоже может помочь, но главную роль здесь играют нематематизированные исследования геологического строения разных участков нынешней Земли, распределения в прошлом животных и растений. Расхождения же между геологами, как и между палеонтологами, обычно настолько велики, что многообразие предлагаемых карт может обескуражить кого угодно.
Тем не менее пробные расчеты, выполненные для некоторых эпох, показывают, что, приняв некоторые из предложенных палеогеографических реконструкций, можно хотя бы оценить, каким принципиально мог быть климат. В частности, удалось показать, что нет ничего необычного в климате без полярных шапок и со средней температурой океанических вод около 15°. Про такие времена в уже цитировавшейся статье С. С. Зилитинкевича с соавторами написано, правда с оговоркой, что этот вывод предварительный: «Более интенсивный перенос тепла из низких в высокие широты приводил к тому, что в тропиках температура почти не отличалась от современной, а в полярных районах была гораздо выше». Это как раз то, что надо для принципиального понимания безморозного климата по всей Земле в раннем карбоне и в другие геологические эпохи.
Вынос тепла из экваториального пояса в высокие широты подразумевает систему соответствующих течений. Свободная циркуляция океанических вод должна была отразиться и на распределении морской фауны, климатическая зональность которой должна быть ослаблена. Именно это мы и видим в раннем карбоне. Нельзя сказать, чтобы состав морской фауны этого времени был всюду одинаковым. Палеозоологи выделяют раннекарбоновые области и провинции. И тем не менее общность морских фаун была достаточно высока. Не случайно по остаткам морских моллюсков, плеченогих, кораллов, фораминифер и других организмов удается проследить на огромных пространствах одни и те же стратиграфические подразделения. В Европе нижний карбон делится на турнейский и визейский ярусы. Выше в европейской схеме размещается намюр, относимый уже к верхнему карбону (силезию). В нашей стране карбон делят на три отдела (нижний, средний и верхний), причем нижний намюр выделяют в самостоятельный серпуховский ярус и включают еще в нижний карбон. Каждому ярусу во времени ставится в соответствие одноименный век. Так вот все три яруса нижнего карбона отечественной схемы можно протянуть на всех материках. Наиболее благополучно обстоит дело с турнейским и визейским ярусами. Серпуховский ярус прослеживается уже несколько хуже, но и его отложения можно указать не только в Подмосковном бассейне, где он был выделен, но и, например, на Таймыре, а также и в Австралии.
Поднявшись еще немного вверх по разрезу, мы теряем возможность столь широких корреляций, или они становятся слишком противоречивыми по разным группам морских организмов. Не случайно палеонтологи так долго путались с острогской фауной и даже считали ее пермской. В морях, омывавших Ангариду с севера в послесерпуховское время, морская фауна сначала резко обеднилась, а затем приобрела местный облик. Эту бореальную фауну, появившуюся в среднем карбоне и эволюционировавшую почти до конца перми, иногда называют верхоянской. Происхождение верхоянской фауны неизвестно, а ее своеобразие можно объяснять по-разному. Причину искали в более холодных водах арктических морей или в изоляции бореального бассейна. Возможна и связь обеих причин.
На границе раннего и среднего карбона на больших пространствах временно прекратилось осадконакопление. В геологической летописи образовались пропуски. Например, почти по всей Северной Америке среднекарбоновые отложения (нижняя часть «пенсильванской системы» по схеме Геологической службы США) лежат на нижнем карбоне (миссисипской системе) с крупным перерывом. Вообще лишь в немногих местах планеты можно видеть непрерывную серию слоев верхов нижнего — низов среднего карбона. Интересно, что как раз на серпуховский век и начало среднего карбона приходятся крупные события в истории магнитного поля Земли. В это время несколько раз происходила инверсия магнитного поля, т. е. Южный и Северный магнитный полюсы менялись местами. Палеомагнитологи называют этот интервал разреза дебальцевской зоной (или дебальцевским «супермагнетемом», по новейшей терминологии).
Можно предположить, что на рубеже раннего и среднего карбона в недрах Земли произошло что-то значительное. Это «что-то» вызвало тектонические подвижки с перерывами в осадконакоплении, а с другой стороны — палеомагнитные инверсии. Ограничилась связь арктических вод с экваториальными. Изоляция привела сначала к обеднению фауны на севере, а затем и к появлению верхоянской фауны. Эта же изоляция приполярного арктического бассейна могла превратить его в мощный холодильник, поскольку его воды плохо прогревались полярным солнцем, а приток теплых вод с юга был ограничен. Причинно-следственная цепь раскручивалась все дальше. Морской приполярный холодильник положил конец безморозному климату по всей Ангариде и даже основательно тряхнул тропическую растительность, вызвав здесь «флористическое обеднение» («флористический скачок» Готана). События не прошли незамеченными и в Гондване, но о ней не будем говорить до главы V.
Естественно, что похолодание климата Ангариды не могло не сказаться и на осадконакоплении. Гипотеза о послесерпуховском похолодании, первоначально выдвинутая, как мы видели, исключительно по палеоботаническим материалам, получила неожиданную поддержку от литологов. Об этом стоит рассказать. Как раз в то время, когда начали появляться мысли о том, что раннекаменноугольные ангарские плауновидные свидетельствуют о безморозном климате, возрос интерес к поискам бокситов в Сибири.
Чтобы поиски шли успешно, надо прежде всего отбросить заведомо бесперспективные отложения. К этому времени уже была хорошо известна фитогеографическая схема Г. П. Радченко, который, как помнит читатель, поместил Сибирь раннего карбона далеко от тропиков. Даже более южную Шотландско-Казахстанскую область он связывал с умеренным климатом. Климат Сибири считал умеренно холодным и умеренно влажным. Бокситы в таких условиях не накапливаются. Им нужен безморозный климат. Помня о выводах Г. П. Радченко, один геолог, в докторской диссертации которого помимо прочего рассматривались и перспективы бокситоносности Сибири, с полным основанием заключил: нижнекаменноугольные отложения Сибири можно заведомо исключить из сферы поисков. К счастью, до поисковых партий эта рекомендация или не дошла вообще, или дошла с опозданием. Я сказал «к счастью» потому, что вскоре после защиты диссертации сразу в нескольких местах Сибири и как раз в нижнем карбоне были найдены небольшие залежи аллитов — алюминиевых руд, отличающихся от кондиционных бокситов несколько меньшим содержанием алюминия. Гипотеза безморозного климата Сибири в раннекарбоновую эпоху нашла неожиданное подтверждение, а с другой стороны, и еще более неожиданно, приобрела практический смысл.
Что же было в Сибири после похолодания? Этому посвящена следующая глава. Но прежде чем переходить к ней, не могу удержаться от некоего «моралите».
Когда-то Ж. Кювье выдвинул свою нашумевшую «теорию катастроф», которую его последователи развили до представления о переворотах, преобразовывавших буквально весь земной шар. На смену этой идее после знаменитых исследований Ч. Лайеля пришло убеждение, что на Земле как сейчас, так и раньше в общем было не так уж беспокойно. Хотя то тут, то там случались землетрясения, обвалы, наводнения, извержения вулканов, но ничто не свидетельствует, что раньше они происходили чаще и с большей синхронностью в разных местах, чем сейчас. Ведь никто не призывает эвакуировать жителей из окрестности Везувия даже тогда, когда просыпается не так уж далеко расположенная Этна. Чем дальше, тем все больше всплывало различий между геологической историей разных регионов. Постепенно в сознании большинства геологов утвердилась «мозаичная модель» структуры и развития Земли. Наша планета представлялась огромной мозаикой, участки которой связаны слабо и сама связь их спорадическая. Катастрофизм стал чуть ли не ругательным словом в лексиконе геологов, и от обвинений в катастрофизме старались откреститься.
Тем временем, пока вытравливались остатки и рецидивы катастрофизма из сознания геологов, накапливалось все больше и больше данных о крупных дефектах мозаичной модели. На фоне местной пестроты начинали просвечивать не замечавшиеся ранее широкие межрегиональные связи, выявлялась подозрительная сопряженность во времени весьма разнородных процессов. В самом деле, нужно было время и серьезное усовершенствование стратиграфических шкал, чтобы можно было поставить в связь, пусть гипотетическую, исчезновение плауновидных в Сибири, дебальцевский «супермагнетем», появление верхоянской фауны и перерыв в осадконакоплении между миссисипской и пенсильванской системами Северной Америки. Если все это действительно следы одного события, по-разному проявившегося в разных местах и на разных компонентах геосистем, то можно говорить о некоем глобальном происшествии, пусть не столь драматическом, как представляли себе катастрофы последователи Кювье. Теперь уже дело не в словах. Кто хочет, может назвать гипотетическую перестройку на рубеже раннего и среднего карбона катастрофой. Здесь уже важно не слово, а понимание происшедшего.
Нам еще предстоит познакомиться с подобными событиями в истории Земли, и не буду забегать вперед с рассказом о них. Я только хочу одновременно и сделать вывод из уже сказанного, и сразу обострить внимание читателя к тому, о чем еще пойдет речь. Наверное, надо воспринимать Землю не как мозаику и не как клавиатуру, клавиши которой работают независимо, а как целостную, органично связанную систему. Обычно мы стараемся постичь мир, классифицируя его явления, а затем незаметно навязываем миру свои классификационные ячейки, втискиваем в них какие-то фрагменты действительности, отрывая их от целого. Разойдясь по факультетам и институтам, мы забываем о том, что природу можно разделить соответственно нашим интересам, лишь прибегая к насилию (которое мы зовем абстракцией). Такое размежевание исследователей давно произошло в геологии. Надо ли удивляться после этого, что геологи наблюдают разные проявления одного события и порой не догадываются о том, что их наблюдения надо только свести воедино и получить живую картину происшедшего на самом деле!
Конечно, груз традиций тяжел. Поэтому прослеживание длинных и разветвленных причинно-следственных связей идет медленно. Не случайна и подозрительность ученых к такой работе, ведь они знакомы с попытками связать что угодно с чем угодно и как угодно. Выход растений на сушу объясняли удалением Луны, вымирание динозавров — вспышкой сверхновой звезды. От таких гипотез, с которыми трезвому и вдумчивому исследователю в общем-то просто нечего делать, зарекаться рискованно. Мало ли что насочиняют люди! И все же будущее наук о Земле — не в дальнейшем разобщении данных, сообщаемых разными дисциплинами или касающихся разных регионов. Их будущее — в глубоком и разностороннем синтезе. Главное, что мы должны делать на пути к нему, — это отождествлять то, что раньше разъединяли. Когда-то было подвигом человеческой мысли отождествить в некотором отнюдь не тривиальном смысле падение яблока и движение планет вокруг Солнца. Отождествления, которые еще ждут науки о Земле, будут не менее тривиальными и порой рискованными из-за соблазна необоснованных спекуляций. Чтобы этот риск был минимальным, требуются учет прошлого опыта, широкая эрудиция, разработанная теория исследований. Как ни труден этот путь, иного, по-видимому, нет.
Глава IV
АНГАРИДА — ДРЕВНИЙ МАТЕРИК

Удивительно удачно слово «предрассудок». Какие-то убеждения и чувства остаются незамеченными, неосознанными, предшествуют рассудку, становятся «пред-рассудком». Именно на науку возложена борьба с предрассудками, и кому, как не ученым, в их ученой деятельности надо в первую очередь самим очиститься от предрассудков. А ученые оказываются такими же людьми и так же не замечают своих научных предрассудков, как «простые смертные» не замечают предрассудков житейских.
Не так давно в ФРГ состоялась специальная конференция, посвященная широко распространенному предрассудку естествоиспытателей. Обсуждалось влияние на научные взгляды всего-навсего тех мест, в которых живет и трудится ученый. Странное дело, законы природы едины повсеместно. Для географа, геолога или ботаника должно быть безразлично, живет ли он в Томске или в Рио-де-Жанейро. Его быт будет отличаться, но при чем здесь научные взгляды? Однако так не получается.
Мы привыкли делить историю Земли на эры, периоды, эпохи и века. Хотя девонский период получил свое название от графства Девон в Англии, а казанский век — от города Казани, мы вроде бы не должны считать эти подразделения отрезком местной истории. Тем не менее некоторые авторитетные стратиграфы убеждены, что если бы геология появилась не в Европе, а в другой части света, то членение истории Земли в международной геохронологической шкале было бы совсем иным.
Еще Теофраст в IV в. до нашей эры различал такие формы роста растений, как деревья, кустарники, однолетние и многолетние травы. Для европейца такое деление настолько естественно, что оно сохраняется в учебниках ботаники до наших дней. Тем не менее в тропиках отделить деревья от трав уже не так просто. Ботаники считают банан многолетней травой. Для пальм, драцен и панданусов был введен термин «древовидные однодольные», про них писали, что это и не деревья, и не кустарники, и не травы, а совершенно особые формы роста. Неизвестно, какую классификацию жизненных форм растений штудировали бы нынешние студенты, если бы ботаника становилась на ноги в окружении влажных тропических лесов.
О влиянии окружающей местной природы на мышление естествоиспытателя я вспомнил не случайно. В палеоботанике так до сих пор и не осознано, насколько сильно укоренилось в представлениях о прошлом растительного мира все то, что сначала было обнаружено в Европе и только поэтому вошло в учебники, заняло место в классификациях и воспринимается как нечто самое главное, самое показательное, тогда как все найденное на других материках менее важно или вообще отклонение из правила.
Передо мной учебник биологии для средней школы 70-х годов. В нем говорится о каменноугольном периоде «с его теплым влажным климатом и воздухом, богатым углекислым газом вследствие сильной вулканической деятельности… Пышная растительность сильно изменила состав атмосферы, обогатив ее кислородом, что имело важное значение для развития наземных животных… С конца каменноугольного периода в связи с усиленным горообразованием, охватившим в следующем (пермском) периоде весь земной шар, влажный климат почти повсеместно сменился сухим. В новых условиях древовидные папоротникообразные стали быстро вымирать; лишь в сырых и тенистых местах удержались более мелкие формы. Вымерли и семенные папоротники…».
Да не обидятся на меня авторы учебника, если я скажу, что подобные сведения были в ходу в начале века. Я не имею в виду ошибки, например, такие, как соотношение гибели птеридоспермов с сухим климатом перми (на самом деле птеридоспермы прожили почти до конца мезозоя и местами чувствовали себя неплохо в триасе и юре, не говоря уж о перми, которую можно считать временем их расцвета). Можно бы простить и то, что выдвигаются весьма сомнительные взгляды об изменении атмосферы в карбоне и перми как твердо установленные факты. Среди специалистов есть сторонники этих взглядов, хотя и крайне немногочисленные. Главный дефект учебника в том, что он приписал всей Земле картину, когда-то нарисованную только для Еврамерийской области карбона, т. е. для тропиков тех времен. Сразу видно, что авторы не удосужились заглянуть в учебники палеоботаники. Даже если бы они взяли довольно старый учебник А. Н. Криштофовича, вышедший в 1957 г., то сразу бы убедились, что типичное для Европы не было характерно для всей планеты.
Я недаром вспомнил о начале века. Именно тогда сложилось мнение, что карбон — время теплого и влажного климата, а пермь — время повсеместной аридизации. Такая картина была нарисована европейскими геологами с помощью палеоботаников на европейском же материале и почему-то была распространена на всю Землю, хотя уже тогда было немало фактов, свидетельствовавших, что европейский «аршин» непригоден для Сибири и южных материков. Южным материкам и их растениям посвящена следующая глава, а сейчас мы познакомимся с флорой Ангариды — материка, располагавшегося, как помнит читатель, на месте нынешней Северной Евразии.
Представление о существовании Ангариды упрочилось в умах геологов в конце прошлого — начале нынешнего века после обобщающих работ австрийского геолога Эдуарда Зюсса, автора замечательной сводки «Лик Земли». Растения, населявшие этот материк, были известны и раньше. Небольшие коллекции их описали немецкие палеоботаники Г. Б. Гейниц и Г. Р. Гепперт, но систематически их впервые начал изучать И. Ф. Шмальгаузен. В 1879 г. вышла в свет его монография, озаглавленная «Юрская флора России». Заголовок нуждается в объяснении. Слово «юрская» в нем появилось не случайно. Шмальгаузен действительно думал, что привезенные из Кузнецкого, Тунгусского и Печорского бассейнов коллекции растений не верхнепалеозойские, а юрские. Его подвела ошибка в документации материала. Он не знал, что происходящие из Кузбасса образцы собраны в двух частях разреза, а именно из верхнепалеозойских (пермских) и юрских отложений.
Сибирская флора верхнего палеозоя тогда была почти неизвестна, а более однообразная по всей Евразии юрская флора была уже неплохо изучена. Шмальгаузен узнал в своей коллекции типично юрские растения и, естественно, решил, что и прочие растения коллекции, якобы собранные в тех же отложениях, относятся к юрскому периоду. Правда, в коллекциях из Печорского и Тунгусского бассейнов типично юрских растений не нашлось. Но в остальном флора этих мест была вполне сходна с кузнецкой, и датировать ее другим периодом Шмальгаузен не стал.
Вскоре после выхода в свет монографии Шмальгаузена К. Л. Космовский усомнился в правильности вывода о юрском возрасте угленосной толщи Кузбасса. Он предположил, и был прав, что здесь есть отложения нескольких геологических систем. Шмальгаузен ответил Космовскому раздраженной статьей.
Чуть позже коллекции, собранные И. П. Толмачевым в Кузбассе, попали к видному французскому палеоботанику Рене Зейллеру. Он пришел к выводу, что растения Кузбасса пермские. Эти коллекции до сих пор хранятся в Музее естественной истории в Париже. Мне довелось их видеть. Хранители музея знали только то, что образцы происходят из Сибири. На образцах были наклейки с одним словом — названием ближайшей от обнажения деревни. Надпись «Афонино» ничего не говорила музейным работникам. А ведь это было то самое Афонино, в обнажении у которого вскрываются палеозойские и юрские пласты. Здесь растения обоих периодов были собраны и смешаны, что и породило долголетний спор о возрасте ангарской флоры. В коллекции, попавшей к Зейллеру, такой смеси не было.
Все стало на свои места только в конце 20-х годов. Томский геолог Л. М. Шорохов в 1927 г. обследовал территорию Кузбасса в поисках строительного и флюсового сырья для Кузнецкого металлургического комбината. Младшим рабочим в его партии был
Д. П. Славнин, позже писавший в биографии Шорохова: «Попутно с выполнением основного задания Л. М. Шорохов собирал ископаемую фауну и флору для пополнения коллекций и создания обменного фонда геологического кабинета… Сам Л. М. Шорохов занимался главным образом сбором ископаемых растений. Его интересовал вопрос о существовании мезозойских отложений в Кузбассе».
Л. М. Шорохов передал собранные им коллекции на определение своему земляку, палеоботанику В. А. Хахлову, который подтвердил, что юрские и палеозойские растения приурочены к разным слоям. Статья В. А. Хахлова вышла в 1928 г. В том же году в Кузбасс приехала М. Ф. Нейбург, работавшая в Геологическом музее АН СССР. Она стала отбирать отпечатки и тщательно отмечать слои, в которых найден тот или иной вид. Выводы Шорохова и Хахлова подтвердились, и ангарская флора окончательно отошла к палеозою.
Вывод в общем был правильным. Мы и сейчас относим ангарскую флору Кузбасса к палеозою (карбону или перми). Однако, как ни парадоксально, конкретные доводы палеоботаников позже оказались нацело ошибочными. Кузнецкие растения были отнесены к родам, ранее описанным из верхнепалеозойских отложений Европы и южных материков. Исследования последних лет показали, что на самом деле растения Кузбасса принадлежат совсем к другим родам, не выходившим за пределы Ангариды, а часто и к другим семействам.
Это далеко не первый случай в истории палеонтологии, когда из ошибочных определений ископаемых растений и животных делались верные заключения о возрасте соответствующих толщ пород. Так же было и при датировке гондванской флоры. В чем здесь дело, неясно до сих пор. Можно предложить такую аналогию. Подобно тому как сейчас можно подметить местный колорит ландшафтов разных стран (березовые рощи средней России, саванны Африки, пейзажи с кактусами в Мексике), так и каждая геологическая эпоха имеет некоторые трудновыразимые особенности. Хотя после «великого раскола» ангарская и еврамерийская флоры десятки миллионов лет эволюционировали независимо, все же некоторые изменения совершались в них параллельно. Полной синхронности в изменениях не было, но в широких временных пределах в разных флорах сохранялось что-то общее. В случае ангарской флоры можно сказать, что в ней почти до конца палеозоя сохранялся «палеозойский облик» и определялся он не столько списком родов или семейств, сколько очень общими морфологическими признаками растений и отсутствием всего того, что характерно для мезозойских флор мира.
Территория Ангариды не оставалась постоянной. С севера, юга и запада она временами затапливалась морем и становилась меньше. Иногда моря уходили, и Ангарнда росла. Обычно ее площадь не превышала 10–15 млн. кв. км, что тоже немало. Самостоятельная флора Ангарской области просуществовала более 100 млн. лет. И все же ее растения почти не попали на страницы учебников палеоботаники. Сама ангарская флора обычно упоминается, приводятся списки некоторых ее растений, иногда даются их изображения. Сообщение этих сведений даже стало традиционным в последние 20–30 лет.
Образованный палеоботаник должен знать род Cheirostrobus, описанный по одному образцу, правда, очень хорошей сохранности, из нижнего карбона Шотландии. Это стробил («шишка») какого-то членистостебельного растения. Его побеги так и неизвестны, но, возможно, были как у клинолистников. Из-за своеобразия в расположении спорангиев хейростробус был выделен в особый порядок, который обычно включается в учебники палеоботаники и в палеоботанические сводки.
В ангарской флоре есть растения не менее своеобразные, причем широко распространенные (не какой-нибудь один образец), просуществовавшие миллионы лет и порой определявшие облик растительности всей Ангариды. И все же почти каждый раз, когда мне приходится принимать иностранных коллег и показывать им свои коллекции, они смотрят на такие диковинные для них растения, как эскимос на верблюда, и только удивляются, что почти ничего не слышали о них раньше. Надо ли пояснять, что многие своеобразные ангарские растения вовсе не попали в общую систему растений и выпали из рассуждений о филогении растительного царства. Правда, они долго оставались весьма плохо изученными, но положение почти не изменилось и после их детальных исследований.
О некоторых ангарских растениях и пойдет дальше речь. Я хочу рассказать о них не только для поднятия интереса к ним, но и потому, что на их примере мне сподручнее всего познакомить читателя с повседневной работой палеоботаника. Такое знакомство кажется мне важным, поскольку иначе трудно оценить достоверность фактов, сообщаемых палеоботаниками.
Я стал палеоботаником случайно. В школьном палеонтологическом кружке мне довелось делать доклад об ископаемых растениях. После первого курса университета я работал в Поволжье и Прикамье в палеоэкологическом отряде, руководимом Р. Ф. Геккером. Нам попадались отпечатки пермских растений, и Роман Федорович заметил, что сейчас ими почти не занимаются. Потом в одном из звенигородских оврагов я натолкнулся на подушку четвертичного известкового туфа, переполненного остатками растений, которым посвятил курсовую работу. А затем руководившая моей работой Т. А. Якубовская показала мне бесхозную коллекцию пермских растений, собранных на Печоре еще в 20-х годах и определенных М. Д. Залесским. Это показалось мне действительно интересным. Я решил заняться палеозойскими растениями и вскоре отправился на консультацию к М. Ф. Нейбург, в те годы самому авторитетному специалисту в этой области. Она занималась флорой Ангариды. По ее совету я устроился на практику в экспедицию Аэрогеологического треста и так впервые попал в Тунгусский бассейн в 1956 г.
К тому времени в Тунгусском бассейне поработало несколько палеоботаников. Первым был И. Ф. Шмальгаузен, которому в 70-х годах прошлого века привез коллекцию с Нижней Тунгуски ссыльный поляк А. Л. Чекановский. Потом тунгусские коллекции изучали М. Д. Залесский, В. А. Хахлов, Н. Г. Вербицкая, М. Ф. Нейбург и ее ученица Е. С. Рассказова. Моим главным руководством была небольшая монография по растениям Тунгусского бассейна, опубликованная перед войной Г. П. Радченко и Н. А. Шведовым. К ним попала большая коллекция, собранная уже упоминавшимся Л. М. Шороховым, замечательным геологом, который вскоре трагически погиб на северо-востоке СССР.
Моя первая экспедиция в Тунгусский бассейн была удачной практикой в бытовом отношении. Я научился вьючить оленей, устраивать баню в палатке и ориентироваться в тайге по аэрофотоснимкам. Но с ископаемыми растениями вышло хуже. Как только мы добрались караваном до района работ и стали устраивать базовый лагерь со складом, мой будущий напарник по маршрутам Э. Л. Фишер прибежал с реки и сказал, что там на берегу валяются отпечатки. Его брат, начальник партии В. Л. Фишер, понял, что мне не терпится заняться своим делом, и великодушно освободил от всех хозяйственных работ. Лагерь оборудовали несколько дней и отсюда же совершали первые маршруты, так что у меня было время покопаться в этом первом обнажении. Последнее слово уместно было бы поставить в кавычки. Это было не обнажение, а высыпка плитчатых алевролитов по берегу реки Учами. За несколько дней я выкопал порядочную яму, сидя в которой с подъема до отбоя набрал несколько сот отпечатков, главным образом листьев кордаитов и семян (фиг. VI).
Расчет пополнить коллекцию во время маршрутов не оправдался. Район был сплошной тайгой, и больше обнажений верхнего палеозоя в тот год я не видел. Уже в Москве геолог той же экспедиции А. П. Степанов, сам определявший ископаемые растения, снабдил меня некоторыми своими образцами, и все это я описал в очередной курсовой работе. Я ходил на консультации к Нейбург, пытаясь с ее помощью разрешить сомнения.
Трудности возникли с определением и родов, и видов. Из доступных мне книг я никак не мог понять, чем отличаются два основных рода листьев кордаитов — Cordaites и Noeggerathiopsis, упоминаемые в каждом учебнике. Еще сложнее был вопрос с видами второго рода, якобы преобладающего в Сибири. В книге Радченко и Шведова была приведена таблица, по которой предлагалось определять виды. Ее устройство было до предела простым. Сначала все листья разбивались на две группы в зависимости от того, на каком расстоянии от верхушки располагается линия наибольшей ширины. Следующая графа делила листья по густоте жилкования (более 22 жилок на 1 см поперечника листа в месте наибольшей ширины или менее этого количества жилок). Потом учитывались изгиб жилок, форма верхушки и т. д.
Каждый свой лист я мог пропустить через эту таблицу и получить на выходе подходящее латинское название. Но результаты мне не нравились. В своей коллекции я видел переходы между теми формами, которые по таблице Радченко и Шведова надо было помещать в разные виды. Что было делать с промежуточными формами, я не знал и с этим приставал к Нейбург. Другие вопросы возникали при чтении книги самой Нейбург, опубликовавшей в издании «Палеонтология СССР» описание верхнепалеозойской флоры Кузбасса. В разделе о листьях кордаитов указывалось, что снизу вверх по разрезу закономерно изменяются их размеры и густота жилкования.
Я слышал от А. П. Степанова, что эта закономерность в общем подтверждается и в Тунгусском бассейне, но мои листья как назло не укладывались в картину. По размеру они подходили к верхнепермским видам, а густота жилкования у них была меньше требуемой. Нейбург же почему-то не спешила мне помочь. Она решительно не соглашалась сказать мне, какие же виды я привез с Учами, а, когда я сам решался дать название тому или иному отпечатку, придиралась к моему решению. Однажды она сказала мне нечто совсем странное, когда я показал ей какой-то невразумительный обрывок: «Сережа, почему вы хотите обязательно определить каждый отпечаток? Отложите этот обрывок, пусть полежит. Может, со временем что-то выяснится».
К такому повороту дела я был совершенно не готов. На занятиях по палеонтологии и на большом практикуме по систематике высших растений, который я посещал на биологическом факультете, никогда не бывало, чтобы экземпляр не лез в систему. Если такое случалось, то это означало ошибку в работе. Преподаватель подходил, и все разъяснялось. Я понимал, что мне могли попасться новые виды, но первое же поползновение сделать этот вывод в курсовой работе Нейбург решительно пресекла.
Только потом я начал понимать, в чем было дело. Нейбург прекрасно осознавала, что неурядицы с моими листьями связаны не с неполнотой существовавшей классификации листьев кордаитов, которую надо только дополнить новыми видами, а с серьезными дефектами самого принципа построения классификации. Хотя мой материал был собран лишь в одном обнажении, но его было достаточно много — больше, чем обычно собирали в одном слое обнажения (мой материал происходил из одного небольшого слоя). И на большом материале границы между признанными видами стали размываться, появлялись переходные формы. Самое простое решение в таком случае — выделить переходную форму в отдельный вид, но между этим видом и прежними получаются новые переходные формы, теперь уже с двух сторон. Проблема не решается, а становится еще более запутанной. Это и значит, что принятая классификация дефектна и разрывает то, что мой материал заставлял объединять.
В очередной мой приход Нейбург показала несколько фотографий, полученных от томской аспирантки М. Д. Артамонцевой. Она попробовала изучать микроструктуру листьев кордаитов, снимая с них прозрачные пленки (реплики). Отпечаток покрывался аптечным коллодием, который, подсыхая, повторял микрорельеф отпечатка. Высохшую реплику можно снять с образца и рассматривать микрорельеф под микроскопом при большом увеличении. На фотографиях Артамонцевой были видны прекрасные оттиски клеток кожицы (эпидермы) листа. К тому времени большое систематическое значение структуры эпидермы было показано на примере многих растений, современных и ископаемых. Нейбург дала мне понять, что усовершенствование систематики листьев кордаитов возможно скорее на пути более углубленных исследований микроструктуры, чем за счет дробления видов по форме листа и густоте жилкования. Надо было обращаться к микроструктуре, ей я и посвятил свой диплом.
К сожалению, палеоботанику редко приходится сталкиваться с таким материалом, на котором хорошо видны и общая морфология растения, и детали его микроструктуры. Обычно, чтобы выколотить крупные листья из слоя целиком, порода не должна быть разбита на мелкие плитки. Таковы песчаники, которые плохо передают тонкие структурные признаки листа. Наоборот, алевролиты и особенно аргиллиты (глинистые сланцы) прекрасно передают микроструктуру отпечатка, но часто рассыпаются на мелкие плитки, каждая из которых вмещает только небольшую часть листа. Собрать все обломки листа, конечно, можно, но эта работа очень трудоемка. Во всяком случае геолог, собирающий остатки растений для определения возраста пород, не будет корпеть над мелкими плитками и подбирать из них целые листья. В коллекциях, доставленных геологами, оказывается слишком мало материала, пригодного для детальных микроструктурных исследований. Такой материал палеоботаник должен собирать сам.
После университета я пришел в Геологический институт, где стал работать под руководством М. Ф. Нейбург. В 1959 г. она отправила меня в Тунгусский бассейн собирать листья кордаитов для детальных исследований. Здесь надо пояснить, почему речь все время идет именно об этих растениях. Дело в том, что после исчезновения зарослей плауновидных раннего карбона по всей Ангариде их место заняла совершенно иная флора. Она, собственно, и считается типично ангарской.
Сначала в этой флоре главную роль играли папоротниковидные растения. Их систематическая принадлежность не выяснена до сих пор, так как органы размножения обнаружить пока не удалось, неизвестно и анатомическое строение побегов. Некоторые из этих растений имели округлые листья-перышки с веерным жилкованием, внешне похожие на семенные папоротники («птеридоспермы») европейского карбона (фиг. VII). Другие растения больше похожи на обычные папоротники, но на их листьях никогда не встречаются спорангии. Поэтому палеоботаники склонны включать и эти растения в число птеридоспермов.
Вместе с этими папоротниковидными сначала в небольшом количестве, а чем выше по разрезу, тем все чаще и чаще встречаются ланцетные и линейные листья, прочерченные множеством слаборасходящихся жилок. Похожие листья имели европейские кордаиты — крупные деревья с древесиной, напоминающей древесину хвойных.
Сначала все листья ангарских кордаитов относили к одному виду, но потом Нейбург заметила их изменения по разрезу в Кузбассе. Соответственно стали выделять разные виды, по распределению которых в разрезе можно было делить сам разрез на части и использовать эти данные для расшифровки геологического строения угленосных бассейнов. Нередко в породах нет никаких других остатков (это особенно характерно для Тунгусского бассейна), и листья кордаитов становятся важнейшим средством для привязки изолированных обнажений к сводному геологическому разрезу района. Соответственно правильность геологических построений оказывается в прямой зависимости от качества систематики листьев.
Отправляясь в экспедицию в 1959 г. вместе с коллектором И. И. Лисицей, я уже не опасался, что ничего не смогу собрать. Мы ехали на Нижнюю Тунгуску, по берегам которой и вдоль ее притоков собирали прекрасные коллекции многие геологи и палеоботаники. Но нужно было искать слои с отпечатками, сохранившими микроструктуру поверхности. Клетки эпидермы не настолько мелкие, чтобы для определения качества материала надо было возить с собой коллодий и микроскоп. Обычно можно убедиться в том, что клетки сохранились, с двадцати- и даже десятикратной лупой. Насколько часто встречаются слои с такими отпечатками, было еще неизвестно. К счастью, проблемы найти подходящие слои не возникло. От обилия материала разбегались глаза. Мы сидели на каждом обнажении по нескольку дней, так что под конец экспедиции стали побаиваться, что наши резиновые лодки не выдержат груза.
 Рис. 10. Мужские стробилы (Cladostrobus) paстений, листья которых относят к роду Rufloria
Рис. 10. Мужские стробилы (Cladostrobus) paстений, листья которых относят к роду Rufloria
Свою работу я выполнял, тесно сотрудничая с геологами Аэрогеологического треста, особенно с Г. Н. Садовниковым, некоторое время работавшим в Геологическом институте. Мы шли к одной цели разными путями. При составлении сводного разреза угленосного верхнего палеозоя вдоль Нижней Тунгуски и ее притоков он опирался на данные геологической съемки и на скудные тогда материалы по бурению. Я делал упор на сопоставление частных разрезов друг с другом через Кузбасс, учитывая прежде всего данные по микроструктуре листьев кордаитов, что было делом слишком новым и не вызывало большого доверия геологов. Тем более было важно, что мы с Садовниковым пришли к одним и тем же выводам.
Наше сопоставление тунгусских обнажений друг с другом и с разрезом Кузбасса отличалось от принятого другими геологами. К нашему мнению стали прислушиваться и поверили в необходимость микроструктурного изучения листьев кордаитов много позже, когда Л. В. Глухова, С. Г. Горелова и Л. В. Меньшикова детально проследили смену микроструктурных типов листьев кордаитов в Кузбассе, а М. В. Дуранте обнаружила те же закономерности в Монголии. Одновременно мы с Дуранте проследили эти закономерности в Верхоянье, где переслаиваются континентальные отложения с растениями и морские отложения с фауной. Это дало возможность существенно скорректировать сопоставление сибирских угленосных толщ с международной стратиграфической шкалой.
Нельзя сказать, что изучение кордаитовых листьев Ангариды в основном закончено. До такой систематики, когда палеоботаник может легко определять виды и составлять списки, еще очень далеко. Виды и роды, устанавливаемые по изолированным листьям, могут быть полноценными только тогда, когда их можно хотя бы приблизительно сопоставить с другими частями тех же растений. Тут пока почти нечем похвастаться. Органы размножения ангарских растений, листья которых мы называем кордаитовыми, еще плохо изучены. Но уже достаточно ясно, что эти органы размножения были иными, чем у европейских кордаитов.
В ангарской флоре особенно много своеобразных листьев, относимых к роду Rufloria. У них устьица были собраны в узких желобках, пробегавших между жилками на нижней стороне листа. Недавно удалось связать с верхнепермскими руфлориями мужские спороношения, которые совершенно непохожи на кордаитовые, а скорее напоминают мужские стробилы мезозойских хвойных, хотя пыльца иная (рис. 10, фиг. VIII).
 Рис. 11. Ангарские (А) и гондванские (Г) филлотеки; имея сходные побеги, они относятся к разным семействам
Рис. 11. Ангарские (А) и гондванские (Г) филлотеки; имея сходные побеги, они относятся к разным семействам
Хотя с легкой руки А. Н. Криштофовича мы нередко зовем верхнепалеозойскую растительность Ангариды, особенно ее сибирской части, кордаитовой тайгой, это не значит, что других растений там не было. Второе место после кордаитов в ангарских захоронениях занимают различные членистостебельные, которых палеоботаники в обиходе именуют собирательным словом «хвощи», осознавая, разумеется, что это лишь далекие родственники современного хвоща. Эти растения были отнесены И. Ф. Шмальгаузеном к гондванскому роду Phyllotheca (рис. 11), что сыграло немалую роль в сближении ангарской и гондванской флор, долго считавшихся родственными. Были в ангарских лесах и папоротники.
В целом не будет прегрешением против истины сказать, что мы еще очень плохо знаем ангарскую флору. Ее растения появляются в геологической летописи внезапно, с оборванными родственными связями, как будто это какие-то пришельцы, взявшиеся неизвестно откуда, заселившие все северные внетропические земли и исчезнувшие в конце палеозоя. Мы ничего не можем сказать о потомках типично ангарских сибирских растений. Может быть, этих потомков и не было, а «кордаитовая тайга», пошумев десятки миллионов лет кронами своих деревьев, ушла в небытие, оставив мощные пласты углей и многочисленные палеоботанические проблемы.
Глава V
ПАЛЕОЗОЙСКИЕ АНТИПОДЫ

Если бы путешественники, проехавшие по позднепалеозойской Ангариде, захотели затем познакомиться с Гондваной, им пришлось бы перебраться через океан Тетис. Он отделял Гондвану от северных материков и лежал на месте современного Средиземноморья, горных хребтов Средней Азии и уходил дальше на восток и юго-восток. Очертания этого океана реконструируются по-разному в зависимости от того, признают или нет авторы реконструкций движение материков, т. е. принадлежат ли они к числу «мобилистов» или «фиксистов». На мобилистских реконструкциях, которые уже почти вытеснили фиксистские из специальной литературы, Тетис обычно изображают в виде огромной треугольной вырезки, внедряющейся с востока в массив сползшихся материков (рис. 12). Чтобы быть точным, слово «сползшихся» надо заменить на «еще не расползшихся», поскольку широкий треугольник Тетиса существовал в позднем палеозое, а в мезозое начал закрываться. Индия, первоначально граничившая с Африкой, Мадагаскаром, Антарктидой и Австралией, поехала на север и своей тушей потеснила Тетис, окончательно исчезнувший уже в кайнозое. Тогда на месте его и вспучились высочайшие в мире горы.
Эта история, широко проникшая на страницы научной и популярной литературы, теперь аргументирована самыми разнообразными наблюдениями, среди которых наибольший пиетет вызывают геофизические и региональные геологические. Иногда вспоминают и палеонтологию, но к ее заявлениям прислушиваются лишь тогда, когда она не свидетельствует против геологических и геофизических заключений. В противном случае к палеонтологии относятся как к назойливому насекомому. Ее выступление расценивают как заявление не слишком сильного государства, поддержкой которого при случае можно попользоваться, но протесты при крупной стратегической игре нечего воспринимать всерьез.
 Рис. 12. Океан Тетис до распадения Гондваны; слева — реконструкция расположения гондванских материков до их расхождения, по Э. Д. Баррону, К. Г. Харрисону и В. В. Хэю
Рис. 12. Океан Тетис до распадения Гондваны; слева — реконструкция расположения гондванских материков до их расхождения, по Э. Д. Баррону, К. Г. Харрисону и В. В. Хэю
Между тем палеонтология заслуживает большего внимания. Ее прошлые заслуги в решении геотектонических проблем достаточно велики, чтобы признать за ней способность сообщать сведения, которыми сообразительный геолог пренебрегать не будет. Начнем с того, что без данных палеонтологии не было бы и представлений о Гондване. Если же из нынешних глобальных тектонических схем выбросить Гондвану, то оставшееся сойдет разве что за карикатуру. Ведь Гондване принадлежит не менее трети нынешней суши, а раньше доля Гондваны могла быть и еще более значительной.
В цитате из учебника по биологии, к которому я придирался в предыдущей главе, гондванский материал нашел такое же отражение, как и ангарский. Иными словами, он не нашел никакого отражения. А ведь именно Гондвана еще раньше, чем Ангарида, ниспровергла миф о повсеместной сухости климата в перми и о планетарном влажном и жарком климате карбона. Еще в прошлом веке в верхнем палеозое Гондваны были найдены толщи, настолько похожие на оставленные отступившим ледником, что в их ледниковой природе с тех пор мало кто сомневался. Следы верхнепалеозойского оледенения нашли почти сто лет назад на всех гондванских материках, кроме Антарктиды, а теперь они известны и на ней. История гондванского оледенения еще довольно темна, но ясно, что оно было крупным, неоднократным, длительным.
Растения, которые встречаются в прослоях между ледниковыми отложениями и выше их, тоже не свидетельствуют о жарком и влажном климате, Гондванскую флору обычно считают умеренной или даже холодно-умеренной. В перми гондванский климат не был сухим, поскольку по всему материку после ухода ледников в большем или меньшем количестве откладывались угленосные толщи, причем пласты угля бывают мощнейшими. И все это, к сведению авторов злополучного учебника, было известно чуть ли не сто лет назад.
В проникновении на страницы палеоботанических руководств гондванским растениям повезло больше, чем ангарским. Впрочем, и история их изучения более почтенна, да и до гондванских стран европейским исследователям было больше дела, нежели до недосягаемой Сибири. Первые ископаемые растения, собранные в Индии и Австралии, попали в руки европейских палеоботаников в начале прошлого века. В 1828 г. Адольф Броньяр опубликовал описание рода Glossopteris (рис. 13), который каждому геологу, работающему южнее экватора и в Индии, знаком не меньше, чем нам всем мамонт.
 Рис. 13. Наиболее характерные гондванские растения: слева — лист гангамоптериса (Gangamopteris), справа — лист глоссоптериса (Glossopteris) и его сечение вблизи основания
Рис. 13. Наиболее характерные гондванские растения: слева — лист гангамоптериса (Gangamopteris), справа — лист глоссоптериса (Glossopteris) и его сечение вблизи основания
Глоссоптериевые листья имеют разную форму, но чаще всего они языковидные, обратнояйцевидные или овальные, с острой или притупленной верхушкой. От суженного основания к верхушке идет довольно толстая средняя жилка, от которой отходят боковые жилки, соединяющиеся перемычками-анастомозами.
В 1875 г. Г. Ф. Блэнфорд обобщил сведения о геологическом строении южных материков и Индии и впервые обратил внимание на общность их верхнепалеозойской флоры, которую назвал «глоссоптериевой». Блэнфорд предположил, что от начала перми и до середины третичного периода существовал Индо-Океанский континент. Он соединил нынешние материки, прилежащие к Индийскому океану. Следом за ним М. Неймайр предположил существование Бразильско-Эфиопского континента на месте нынешней Атлантики. Оставалось соединить оба гипотетических материка, что и сделал в 1885 г. Эдуард Зюсс, предложивший термин «Гондвана».
Гондванская флора давно привлекала внимание палеоботаников, занимавшихся ангарской флорой. В Гондване известны листья кордаитового типа, внешне больше похожие на сибирские, чем на европейские. В 1912 г. М. Д. Залесский отнес такие листья из Ангариды к гондванскому роду Noeggerathiopsis. На обоих материках встречаются членистостебельные с листьями, сросшимися в воронку. Сначала их нашли в Австралии и Индии, а в 1879 г. И. Ф. Шмальгаузен отнес к этому же роду ангарские образцы. Постепенно список общих родов рос. В 1897 г. В. П. Амалицкий нашел в верхней перми Среднего Поволжья раковины моллюсков примерно такие же, какие были раньше встречены в Южной Африке вместе с многочисленными скелетами рептилий и амфибий. Амалицкий решил поискать скелеты таких же животных в пермских отложениях Европейской России. Он действительно нашел их, а с ними отпечатки листьев, внешне очень сходных с глоссоптериевыми. Это было уже сенсацией. По-видимому, именно эта находка подстегнула решение Залесского отнести к одним и тем же родам не только кордаиты Ангариды и Гондваны, но и ряд других растений.
Вывод Залесского о близком родстве флор Ангариды и Гондваны получил широкую известность и признание. Геологи и палеонтологи обсуждали пути, по которым могли растения пересекать Тетис. Строились гипотезы о времени соединения и разъединения этих материков. Сам Залесский считал, что сибирская флора приобрела самостоятельность от гондванской только в мезозое.
Первые трещины в гипотезе Залесского появились в 30-х годах. На XVII Международный геологический конгресс, состоявшийся в Москве в 1937 г., приехали палеоботаники, знакомые с гондванской флорой. Они осмотрели ангарские коллекции и заключили, что общность флор есть, но гораздо меньше, чем считал Залесский. Потом последовала война, и палеоботаникам стало не до этого. Залесский, живший в Орле, был насильно вывезен оккупантами в Германию.
Известный палеоботаник, член-корреспондент АН СССР последние дни перед приходом наших войск жил в полуразрушенном доме, в комнате с вывалившейся наружной стеной и добывал средства для пропитания, таская воду в дома с разрушенным водопроводом. Рассудок его помутился, и он не узнавал старых друзей. Вернувшись на родину, он вскоре умер.
В 1954 г. к вопросу о соотношении ангарской и гондванской флор вновь обратилась М. Ф. Нейбург, которая сократила список общих родов и указала, что за этой общностью не обязательно видеть близкое родство флор. Но в ее списке еще оставались такие роды, как Noeggerathiopsis, Phyllotheca и Gondwanidium, считавшиеся весьма характерными для обеих флор. Когда я пришел работать к Нейбург, она продолжала интересоваться этим вопросом и сетовала, что ей вряд ли удастся побывать в какой-нибудь из гондванских стран и самой покопаться в коллекциях.
Разумеется, меня этот вопрос интересовал не меньше. Особенно хотелось посмотреть листья гондванских кордаитов и их микроструктуру. Вскоре после кончины Нейбург я убедился в том, что наши кордаитовые листья относятся к двум родам и что род Noeggerathiopsis должен быть упразднен. Литературные данные о микроструктуре гондванских кордаитов были, но слишком скудные. Надо было смотреть коллекции.
В 1966 г. такая возможность совершенно неожиданно представилась. Я получил командировку в Индию, смог посетить главные палеоботанические лаборатории этой страны и посидеть с коллекциями. К этому времени я уже трудился над картами палеофлористического районирования, о которых шла речь во второй главе, и поэтому мог на месте прояснить множество вопросов, связанных с Индией.
В Индии есть единственный в мире специальный Палеоботанический институт, названный именем его основателя, известного палеоботаника Бирбала Сахни. Сотрудники этого института, находящегося в столице штата Уттар-Прадеш городе Лакхнау, гордятся тем, что основанием института они обязаны Джавахарлалу Неру. Они не скрывают, что поддержка Неру не была просто данью уважения к палеоботанике. Неру и Сахни вместе учились в Англии и были близкими друзьями.
Я плохо представлял себе индийские палеоботанические лаборатории, где какие коллекции можно посмотреть, и рассудил, что больше всего времени надо провести в Лакхнау. Это было ошибкой, поскольку классические коллекции верхнепалеозойских растений Индии, описанные чешским палеоботаником О. Фейстмантелем, современником И. Ф. Шмальгаузена, хранятся не в Лакхнау, а в Калькутте. Там я тоже побывал, но времени было в обрез. Неделю я провел в лаборатории профессора Д. Д. Панта в Аллахабадском университете, работающего с небольшой группой учеников, настоящих энтузиастов.
О результатах своей поездки в Индию я уже подробно писал в популярной книжке «Из истории растительных династий», вышедшей в 1971 г., и боюсь, что читавшие ее коллеги осудят меня за повторение. Поэтому остановлюсь только на главных результатах, чтобы подробнее рассказать о последующих исследованиях. Результаты эти были для меня неожиданными. Индийские коллеги показали мне все те растения, которые считались общими, ангаро-гондванскими. Вся эта общность стала рассыпаться, как только я начал класть образцы под бинокуляр и изучать тонкие, неважные детали жилкования, микроструктуру и другие не отмеченные в описаниях признаки. У некоторых листьев кордаитов можно было подозревать присутствие желобков между жилками, как у ангарских руфлорий. Оказалось, что такая картина наблюдается только в основании листа, где устьичные желобки имитируются узкими междужильными промежутками. Так называемые кордаитовые листья сопровождаются в Индии совсем другими семенами, и, кроме того, можно видеть переходные формы между этими листьями и теми, которые относят к роду Gangamopteris. Возникло подозрение, кордаиты ли это вообще. Иными оказались и членистостебельные, относимые к роду Phyllotheca.
Даже не эти вскрывшиеся различия растений, прежде казавшихся родственными, привели меня к выводу о разительном различии ангарских и гондванских флор. Гораздо важнее было посещение каменноугольного бассейна Ранигандж, классического места сборов гондванских растений.
От Калькутты в Ранигандж идет экспресс. Вечером мы сошли на станции Баракар. Это название ничего не говорит большинству читателей. Но для меня было удивительно видеть его на станционном здании. Ведь так называется одно из стратиграфических подразделений индийского верхнего палеозоя, в котором еще в прошлом веке были собраны многочисленные ископаемые растения. В их числе была и Barakaria, два вида которой были обнаружены Нейбург в Кузбассе и Печорском бассейне, а затем мной в Тунгусском бассейне. Так что слово «Баракар» на перроне вызвало у меня не менее богатые ассоциации, чем у любителя пива дорожный указатель на Пльзень. Сопровождающие меня геологи разыскали на пристанционной площади предоставленный нам «джип». Он и доставил нас в маленькую гостиницу у подножия Панчетских холмов.
Работая на разрезах угленосного верхнего палеозоя Ангариды, невозможно не встретиться с отпечатками листьев кордаитов. Чуть ли не в каждом слое найдешь хоть один обрывок, а порой листья кордаитов столь многочисленны, что порода становится тонкослоистой. (Такие скопления ископаемых остатков В. А. Красилов удачно назвал «листовыми кровлями».) В раниганджских карьерах я увидел совсем иную картину: растительных остатков тоже немало, есть и «листовые кровли», но все это — глоссоптериевые листья. Разница настолько бросилась в глаза в первом же карьере, что вся многолетняя дискуссия о соотношении ангарской и гондванской флор сразу же потеряла смысл и интерес. Обсуждать стало почти нечего. О каком родстве флор можно говорить, когда общие элементы, даже если они и есть, тонут на совершенно различном фоне. Невольно вспоминались индийский гость в «Садко» и Афанасий Никитин, добравшийся до Индии. То, что купцы ездили из страны в страну, конечно, свидетельствует о связях, но появление на базаре чужеземца, когда толпа пялит на него глаза, лишь лучше подчеркивает независимость стран и народов.
Несколько дней мы ездили по карьерам, осматривали разрезы, собирали отпечатки. Некоторые карьеры были частными и приходилось спрашивать разрешения администрации на посещение. Разрешения выдавались охотно, и, больше того, обычно нам выделяли рабочего с ломом и киркой, который выворачивал плиты. Хотелось найти хоть одно растение из тех, которые считались общими для Ангариды и Гондваны, но, кроме бесчисленных глоссоптериевых листьев, ничего не попадалось. Непривычный облик гондванской флоры выступал тем ярче, что и все вокруг было чужим. Мы привыкли видеть в открытых карьерах экскаваторы и ревущие самосвалы. Здесь же была бесконечная вереница носильщиц с огромными корзинами на головах, в корзинах — уголь и пустая порода. В старых отвалах копались ребятишки и взрослые из соседних деревень в поисках кусков угля для домашних очагов. По вечерам дым от очагов застилал все вокруг, и мы ехали домой как в тумане да еще в кромешной тьме. Днем, переезжая из одного карьера в другой, мы объезжали деревни. Глинобитные лачуги стояли стена к стене вдоль узких улочек. По ним «джип» проехать не мог.
Поездка в Ранигандж всего на несколько дней дала мне больше, чем сотни статей. Оставалось только удивляться, насколько обманчивой может быть картина, складывающаяся из изучения не только литературы, но даже музейных коллекций. Все получалось как в книгах по истории. Как ни старается историк оживить картины прошлого, он не может не исказить пропорции. Ренессанс предстает перед нами как эпоха, освещенная творчеством Петрарки и Леонардо да Винчи. Только проехав по дорогам Италии тех времен, мы могли бы почувствовать, как далеки были творения гениев от повседневной жизни городов и деревень.
То же и в естественных науках. Редкие находки привлекают внимание, описываются в деталях в специальных статьях и занимают почетное место в музейных витринах. Кто будет забивать энтомологические музейные коллекции бесчисленными тараканами, шныряющими чуть ли не в каждом доме? В прекрасном Индийском музее в Калькутте я видел немало глоссоптериевых листьев, а вместе с ними лежали и многочисленные другие растения. Разве придет сразу в голову, что эти «другие» — чрезвычайная редкость? Читая литературу или сидя в музее, можно было понять, что глоссоптериды преобладали в гондванской флоре, но прочувствовать и осознать, насколько они преобладали, было нельзя.
Возвращаясь в Москву, я предвкушал, как познакомлю коллег с результатами. Мне казалось, что проблема соотношения ангарской и гондванской флор решена раз и навсегда. Через несколько лет нашу страну приехал директор института в Лакхнау К. Р. Суранге. Он долго сидел над коллекциями ангарских растений, увидел «листовые кровли» с кордаитами, смотрел микроскопические препараты и по окончании работы присоединился к моим выводам. Проблема в самом деле казалась закрытой. Но недаром говорят, что получение ответа на важный вопрос может всего лишь означать неправильную постановку вопроса, Обычно оказывается, что не учтено что-то весьма существенное. Вскоре я это ясно понял.
Среди коллег, слушавших мои доклады о соотношении индийских и ангарских флор, были не только специалисты по отпечаткам растений («макрофоссилиям»), но и палинологи. Они-то и не разделили мой оптимизм. Надеюсь, читатель помнит, что палинология — наука, изучающая споры и пыльцу растений. Оболочки спор и пыльцы (их часто называют собирательным термином «миоспоры») встречаются в огромных количествах в палеозойских, мезозойских и кайнозойских, особенно угленосных, отложениях. Обычно палинологи не уверены в том, какими растениями продуцировались миоспоры, которые поэтому описываются под самостоятельными родовыми и видовыми названиями. Мне бы следовало сразу поинтересоваться, насколько миоспоры подтверждают мои выводы, сделанные по макрофоссилиям, и уж затем звонить на всех перекрестках. Палинологи сразу указали мне, что звонить преждевременно. Ведь если взять списки миоспор из верхнего палеозоя Ангариды и Гондваны, в том числе и Индии, то в них можно найти очень много общего, особенно в родовом составе миоспор (фиг. IX).
Мало того, те общие морфологические типы миоспор, которые известны на обоих материках, появляются тут и там в одинаковой последовательности. Это особенно ярко видно на тех миоспорах, которые относят к пыльце голосеменных. Сначала появляются пыльцевые зерна с одним воздушным мешком, окружающим тело пыльцы широким ободом. Затем появляется пыльца с двумя воздушными мешками, к которой еще выше добавляется сходная двухмешковая пыльца, имеющая ребристое тело. Почти одновременно появляется и ребристая пыльца без мешков, относимая к роду Vittatina. Правда, последней пыльцы мало в Индии, но много на других гондванских материках. Самое неприятное было в том, что такая последовательность отмечена не только на ангарском и гондванском материале, но и в Еврамерийской (тропической) области. Вполне естественно, что палинологи использовали сходство в последовательности типов пыльцы для сопоставления соответствующих толщ по их возрасту.
Попытка заинтересовать палинологов возникшим противоречием не удалась. У всех есть собственные заботы. У меня их тоже хватало, и поначалу казалось, что придется заняться розыском корней противоречия в отдаленном будущем. Случилось же так, что я снова отправился в Индию по приглашению К. Р. Суранге. Заниматься макрофоссилиями уже не имело большого смысла, тем более что специалисты по ним моих выводов не оспаривали. Было решено взяться за сравнение миоспор. В Лакхнау нет разделения специалистов на палинологов и тех, кто занимается только макрофоссилиями. Я таким преимуществом не обладал и, поскольку времени на подготовку почти не было, отправился в Лакхнау с весьма легковесным палинологическим багажом, в чем и признался Суранге, когда изложил ему свои планы. Тем не менее Суранге планы одобрил. Его сотрудники — Суреш Шривастава, К. М. Леле и X. К. Магешвари — несколько дней с утра до вечера внедряли в меня азы практической палинологии. Пройдя ускоренный курс обучения, я засел за микроскоп.
Главное, с чем предстояло разобраться, — это с соотношением форм мешковой пыльцы разных палеофлористических областей, и прежде всего Ангариды и Гондваны. Запомнить диагностические признаки нескольких десятков родов, выделенных для такой пыльцы, не так уж сложно. Несравненно сложнее научиться различать сами диагностические признаки. Обычно пыльца, извлеченная из породы, сплющена в тонкую лепешку, да часто еще и со сдвигом. Понять, как и с какой стороны прикрепляется мешок, какие пупырышки расположены на оболочке снаружи, а какие изнутри, как изменяется тонкая структура мешка вокруг зерна, далеко не просто. Иногда на реконструкцию общего прижизненного облика единственного зерна уходят многие часы. Приходится работать с самыми большими увеличениями и, постепенно меняя фокус, делать «оптические срезы» зерна в разных плоскостях, а потом мысленно совмещать картины, получавшиеся для разных плоскостей. Мои учителя, особенно Леле и Магешвари, были прекрасными мастерами таких реконструкций и, что было еще важнее для меня, терпеливыми педагогами,
Я жил в гостевых комнатах прямо в институте и мог приходить в лабораторию в любое время, а покидать ее только для сна и еды. Меня снабдили литературой и многочисленными препаратами (некоторое число препаратов я привез с собой). Очень медленно картина стала проясняться. Обнаружились различия между формами миоспор, казавшимися общими для Ангариды и Гондваны. В таких случаях можно было утверждать, что общность родовых названий пыльцы не свидетельствует о том, что и роды родительских растений были одними и теми же. С таким параллелизмом форм, не подкрепленным родством, палеонтологи знакомы давно. Он проявлялся и в макрофоссилиях верхнего палеозоя. В конце концов я пришел к выводу, что именно параллелизм ответствен за общность миоспор Ангариды, Гондваны и отчасти Еврамерийской области. Этот вывод, впрочем, страдал изрядной гипотетичностью. Мои индийские коллеги в общем были с ним согласны, но тоже понимали, что решающие доказательства еще предстоит получить.
Как приятно было бы написать, что возникшая с миоспорами проблема была столь же успешно закрыта, как и ранее существовавшая проблема общности ангарских и гондванских макрофоссилий! Однако до «закрытия» проблемы еще далеко. Правда, с тех пор удалось привязать некоторые типы миоспор, почти неотличимых под микроскопом, к совершенно разным группам растений. Например, выяснилось, что двухмешковая пыльца с ребристым телом в гондванских
палинологических комплексах продуцировалась растениями с глоссоптериевыми листьями.
Пыльца совершенно того же облика, встречающаяся огромными массами в верхнепермских отложениях европейской части СССР, принадлежит совсем другим растениям — пельтаспермовым птеридоспермам, о которых пойдет речь в главе «Корни мезофита». Такую же пыльцу находили и в мужских органах размножения некоторых гондванских хвойных растений мезозоя. Короче говоря, присутствие такой пыльцы не дает возможности сказать, какая группа растений жила в данном районе. Это все равно что пытаться по одному слову судить о фразе. Многие формы миоспор так и остаются не привязанными к исходным растениям, известным по макрофоссилиям. Кроме того, вполне возможно, что от каких-то растений вообще не сохранялось в ископаемом состоянии ничего, кроме миоспор.
Особенно непонятным остается сходство в последовательности типов миоспор в разных палеофлористических областях. И для объяснения этого сходства особенно важно знать, одновременно или нет происходили смены основных типов в разных местах. Тут-то мы и натыкаемся на проблему более чем столетней давности.
Периоды и более мелкие подразделения международной геохронологической шкалы были сначала установлены на европейском материале. По мере того как геологи знакомились с другими материками, с разрезами, отличающимися от европейских, геохронологическая шкала дополнялась и изменялась. Иногда стратиграфические шкалы, составлявшиеся для удаленных от Европы регионов, удавалось слить с международной шкалой. Разные региональные шкалы существуют и сейчас, но с большей или меньшей точностью их можно сопоставить с общепринятыми подразделениями. Пожалуй, труднее всего пришлось стратиграфам тогда, когда они попытались привязать к стандартным подразделениям верхнепалеозойские толщи Гондваны. Довольно долго обсуждался вопрос, принадлежат эти толщи к палеозою или мезозою.
Еще Броньяр отметил сходство между родом Glossopteris и листьями, известными в юрских отложениях Европы и позже выделенными в род Sagenopteris. Мнение, что гондванская глоссоптериевая флора принадлежит мезозою, разделялось многими исследователями до 60-х годов прошлого века. Однако затем все постепенно пришли к убеждению, что эта флора палеозойская. Любопытно, что палеоботанические аргументы в пользу этой точки зрения позже оказались ошибочными. Например, О. Фейстмантель описал в глоссоптериевой флоре розетки листьев, очень похожих на листву карбоновых еврамерийских каламитов (древовидных членистостебельных, дальних родственников современных хвощей). Были сопоставлены и кордаитовые листья Гондваны и Европы. Теперь от всех этих сопоставлений отказались, но вывод был сделан и по счастливой случайности в главном оказался правильным. В палеозойском возрасте глоссоптериевой флоры теперь никто не сомневается.
Для современного стратиграфа привязка к международной геохронологической шкале с точностью до эры (в данном случае палеозойской) вызывает улыбку. Он требует гораздо более точной привязки. Гондванский верхний палеозой в этом отношении редкий случай. Мы не имеем представления, на каком уровне надо проводить границу карбона и перми в гондванских разрезах. Расхождения между стратиграфами все еще очень велики. Слои, которые одними геологами сопоставляются с нижней Пермью, другие геологи относят к среднему карбону. Дело в том, что фауна и флора Гондваны слишком своеобразны. Девонские и раннекаменноугольные морские фауны Гондваны достаточно сходны с европейскими, чтобы уверенно различать по ним отдельные ярусы (века). Но уже в раннем намюре положение начинает меняться: степень своеобразия гондванской фауны быстро возрастает и прямые сопоставления с Европой становятся невозможными. Напомню, что с того же уровня становится невозможным сопоставлять с европейскими разрезами и ангарские толщи. Географическая дифференциация на севере и на юге шла одновременно, причем у наземных растений тут и там она развивалась быстрее, чем у морских животных.
При всем своеобразии гондванской флоры и фауны среди современных исследователей преобладает мнение, что глоссоптериевая флора целиком относится к перми. Хотелось бы пояснить, почему стратиграфы пришли к этому выводу, но я не в силах этого сделать, и не потому, что не знаком с соответствующей литературой, а потому, что не могу обнаружить логики в рассуждениях стратиграфов. Их аргументы таковы. Во-первых, говорят они, в морских отложениях, залегающих среди толщ с глоссоптериевой флорой, есть раковины плеченогих (брахиопод), которые относятся к родам, известным в перми северного полушария. Во-вторых, в палинологических комплексах, сопровождающих глоссоптериевую флору, преобладают роды миоспор, характерные в северном полушарии для перми. В-третьих, появление глоссоптериевой флоры знаменует собой новый этап в эволюции гондванского растительного покрова, а такой важный рубеж должен соответствовать рубежу между периодами.
Я внимательно изучал эти и другие аргументы, которыми не буду утомлять читателя, но признать их обоснованными не смог при всем желании. Беседы со специалистами по гондванской стратиграфии не столько рассеяли мои вопросы, сколько укрепили сомнения. Пусть читатель судит сам. Действительно, между брахиоподами северного полушария и Гондваны есть общие роды; верно и то, что эти роды на севере определенно пермские. Более того, они верхнепермские. В лучшем случае они заходят в морских разрезах Сибири в верхи нижней перми, а дальше вниз не идут. Слои с этими же родами в Австралии и Южной Америке невозможно отнести к верхней перми, поскольку они лежат ниже отложений, которые удалось довольно уверенно отнести к нижней перми (причем к низким ее частям) по находкам раковин головоногих моллюсков, к счастью примерно одинаковых на разных материках.
Если северные и южные брахиоподы действительно относятся к одним и тем же родам (и здесь не строит козни параллелизм раковин), то приходится предполагать, что роды эти сначала появились в гондванских морях, а затем отправились на север. Но тогда ничто не обязывает нас считать, что они появились в Гондване именно в перми, а, скажем, не в среднем или в позднем карбоне. Видимо, для палеонтологов сработал эпитет «пермский», который издавна привязывался к сибирским раковинам. Допустить находки этих раковин в другой (нижней) части перми, но все же перми палеонтологи согласились, но «опустить» эти «пермские» роды в карбон показалось чем-то слишком рискованным.
О том, чего стоят палинологические данные, я уже говорил. Появление сходных миоспор в северных и гондванских разрезах пока не удалось подкрепить привязкой этих микроскопических оболочек к материнским растениям, а в некоторых случаях ясно установлены различия растений, продуцировавших весьма сходные миоспоры. Однако сходство латинских названий в палинологических списках продолжает гипнотизировать стратиграфов.
Третий аргумент еще слабее. Границу карбона и перми в международной стратиграфической шкале проводят по появлению определенных групп ископаемых организмов. Многие считают, что появление этих групп было важным событием в истории жизни и даже, если угодно, новым этапом в эволюции. Но из этого никак не следует, что появление какой-то иной группы в другом месте Земли должно было произойти на том же рубеже. Появление глоссоптериевой флоры безусловно было важным событием в гондванской истории, но из одного этого никак не следует, что надо приурочивать это событие к границе каменноугольного и пермского периодов.
Я не исключаю, что в отнесении всех отложений с глоссоптериевой флорой к перми немалую роль сыграла ошибка, о которой теперь мало кто помнит. Осенью 1976 г. вместе с лионским палеоботаником Ж. Баралем я смотрел коллекции в Горной школе в Париже. К этому времени администрация школы легкомысленно решила расстаться с огромными палеонтологическими коллекциями, концентрировавшимися в музее школы о прошлого века. Правда, коллекции не выбрасывали на помойку, а сложили в ящики и лотки под витринами. Предполагалось передать их другим учреждениям. Работа была проделана без излишних предосторожностей, и возникла путаница в образцах. Поэтому в поисках нужных коллекций приходилось пересматривать лоток за лотком и не по этикеткам, а по внешнему виду вылавливать образцы, знакомые по давно опубликованным иллюстрациям. В ходе этой работы я неожиданно наткнулся на образцы, по виду происходившие из европейского верхнего карбона, но имевшие наклейки с совершенно иным адресом. На них было написано, что это образцы, собранные геологом Лапьером в бассейне Тете в Мозамбике.
Подозреваю, что судьба этих образцов могла бы быть довольно печальной. Никакой документации при них не было, а в музейных описях они не значились. Впрочем, и сами описи были в полнейшем беспорядке, так что образцы при случае могли просто ликвидировать как бесхозные. Между тем интерес лапьеровской коллекции немалый, и история, происшедшая с ней, любопытна. Коллекция Лапьера была изучена в 70-х годах прошлого века известным французским палеоботаником Р. Зейллером. Он обнаружил отпечатки растений, тогда уже хорошо известных в верхнекарбоновых (стефанских) отложениях Европы. Без особых колебаний он отнес вмещающие породы к стефанскому ярусу карбона и опубликовал результаты в одном из геологических журналов. Находка типично стефанских растений на юге Африки, среди многочисленных местонахождений глоссоптериевой флоры, была крайне важной. Если Африка в позднем карбоне была заселена растениями европейского типа, то оставалось отнести слои с глоссоптериевой флорой к перми. В том, что глоссоптериевая флора — палеозойская, Зейллер к тому времени уже не сомневался. Пермский возраст получался методом исключения. Более того, можно было предположить, что Гондвана до конца карбона была заселена той же флорой, что и Европа. Значит, время разделения растительного покрова Земли на области — пермь. Такой вывод и был сделан Зейллером в специальных статьях.
Поскольку Зейллер был одним из наиболее авторитетных палеонтологов конца прошлого — начала нынешнего века, его заключение не могло не произвести впечатление на стратиграфов. Коллекция Лапьера долго не привлекала внимания палеоботаников, хотя о выводах Зейллера помнили. Лишь в 40-х годах к ней снова обратился португальский палеоботаник К. Тейшейра, обрабатывавший остатки гондванских растений из Анголы. На одном из международных конгрессов Тейшейра выступил с докладом, где повторил мысли Зейллера. В короткой дискуссии по докладу принял участие видный голландский палеоботаник и крупный специалист по стратиграфии верхнего палеозоя В. Йонгманс. Он скептически отнесся к идеям Зейллера, поддержанным Тейшейрой, и высказал предположение, что с коллекцией Лапьера произошла путаница в самом начале. Совершенно невозможно поверить, чтобы чисто европейский комплекс растений был найден в недрах Гондваны.
Если бы Йонгманс видел описанные Зейллером образцы, у него бы не возникло сомнений в правильности выдвинутого предположения. И вид породы, и облик отпечатков были таковы, что не возникало сомнений: Зейллер действительно описал европейские образцы, кем-то ошибочно этикетированные как африканские. К тому же с тех пор в районе Замбези неоднократно работали геологи и привозили коллекции растений, но все это были обычные гондванские растения. В геологическом разрезе района просто не осталось места, в которое можно втиснуть вопреки всем установленным закономерностям образцы, чуть не затерявшиеся в Горной школе.
Итак, Зейллер жестоко ошибся. Лишились обоснования его доводы о пермском возрасте гондванской флоры. Но эта трещина в фундаменте давно успела скрыться под новыми постройками. Наверное, стоило бы проследить по литературе, как конкретно повлияли взгляды Зейллера на отнесение глоссоптериевой флоры к перми. Впрочем, едва ли это изменит сложившуюся уже привычку.
Читатель может спросить, а почему я так возражаю против мнения о появлении глоссоптериевой флоры в начале перми? Какие есть основания относить это событие к карбону? Надо честно признаться, что и эти основания весьма шаткие. Я не доказываю, что надо относить раннюю глоссоптериевую флору к карбону, а лишь хочу подчеркнуть, что проблему зря считают решенной. Если же мы откажемся от настолько же привычного, насколько и необоснованного решения, то придется ставить вопрос снова. Придется искать пути к его решению. Таких путей может быть несколько. Во-первых, надо шире привлечь определение возраста гондванских пород по радиоактивным изотопам. Такие анализы уже делались, и, кстати, первые полученные результаты указали на позднекарбоновый возраст некоторых слоев Австралии с глоссоптериевой флорой. Во-вторых, надо более тщательно проследить палеоклиматическую историю обеих внетропических областей — Ангариды и Гондваны. Судя по новейшей истории климатов Земли, похолодания и потепления происходят в обоих полушариях с поразительной синхронностью. Если удастся восстановить с достаточной полнотой всю последовательность похолоданий и потеплений как в Ангариде, так и в Гондване, то можно будет сопоставить осадочные толщи обоих материков. В-третьих, в позднем палеозое было несколько заметных палеомагнитных событий, когда происходил повторный обмен местами между Южным и Северным магнитными полюсами. По остаточной намагниченности горных пород можно сопоставлять разрезы, и этот метод тоже может помочь при сопоставлении гондванских разрезов с международной геохронологической шкалой.
Перечисленные пути не есть что-то новое и неожиданное для стратиграфа. По ним пробовали двигаться, хотя и в небольшом масштабе, часто останавливаясь раньше времени. Когда в 1972 г, появилось сообщение, что радиоизотопный (в данном случае рубидиево-стронциевый) анализ указал на позднекарбоновый возраст слоев с ранней глоссоптериевой флорой в Австралии, то австралийский палинолог Б. Балм написал специальную заметку о важности полученного результата и о том, что представление о появлении этой флоры в перми может быть ошибочным. Анализ следовало повторить и соответствующий комплекс растительных остатков описать, чего так никто и не сделал за прошедшие с тех пор годы. Палеомагнитная шкала перми до сих пор содержит крупные прорехи, неизученные интервалы разреза, без наполнения которых невозможно уверенно опираться на палеомагнитные данные при сопоставлении верхнепалеозойских толщ.
Еще предстоит разобраться и в палеоклиматической истории как Ангариды, так и Гондваны. Здесь намечены лишь отдельные вехи, из которых еще рано складывать полную последовательность даже самых важных климатических эпизодов. Важные сведения о соотношении палеоклиматических событий в Ангариде и Гондване можно было бы получить, сравнивая богатство последовательных флор и фаун, тщательно отмечая моменты, когда на север и на юг проникали теплолюбивые формы из экваториального тропического пояса. И эти исследования, которые могут выявить эпохи потеплений, еще предстоит провести.
Отсутствие надежной привязки верхнепалеозойских отложений Гондваны к международной геохроиологической шкале не просто частный пробел в наших знаниях. Поздний палеозой — интереснейшая и важнейшая глава в летописи Земли. Это было время климатических перестроек, крупных оледенений, горообразования, вулканизма. В позднем палеозое произошли события, во многом определившие последующую историю как Земли в целом, так и жизни на ней. Особенно важное значение имеет расшифровка позднепалеозойской истории Земли для решения широкого круга проблем, связанных с предполагаемым перемещением материков. Если материки двигались (а сейчас в этом мало кто сомневается), то надо восстановить их траектории, а для этого надо начинать со времен, когда они еще не успели разойтись и когда Гондвана была единым континентом. Это и было в позднем палеозое. В былом единстве Гондваны сейчас почти не приходится сомневаться, но, как именно составить ее из расползшихся по земной поверхности материков, не известно. Гипотетических реконструкций Гондваны накопилось уже немало, и выбрать из них одну истинную так и не удается. Необходимое условие выбора — детальное знание позднепалеозойской истории, которую нельзя расшифровать, пока не удастся предложить надежное сопоставление стратиграфических шкал Гондваны и северных территорий.
Глава VI
ПАССАЖИРЫ ИЛИ ПОСТОЯЛЬЦЫ?

Из прослушанных на студенческой скамье курсов самые тоскливые воспоминания вызывает у меня история геологии. Было невероятно скучно выслушивать поминальник имен, какие-то нелепые и наивные концепции средневековья, ошибочные теории недавнего прошлого. Да и что нам, будущим геологам, было до того, что какой-то француз Эли де Бомон создал никому уже не нужную контракционную гипотезу. История геологии представала перед нами в виде скучного рассказа о том, как глуповатые и необоснованные гипотезы заменялись истинными и теперь уже незыблемыми теориями. От своих однокашников с других факультетов университета я слышал такие же жалобы.
Мое отношение к истории науки начало меняться после долгих бесед с известным исследователем Байкала В. В. Ламакиным, с которым мы жили по соседству и вместе ходили пешком в институт. Он заявил, что находит в старой литературе больше, чем в новейших поступлениях в нашу библиотеку. Хотя это и было некоторым преувеличением, но побудило меня внимательнее относиться к работам далеких предшественников. Интерес к истории геологии подогрел и Ю. Я. Соловьев, занимавшийся историей палеогеографии в России и обнаруживший, что А. П. Карпинского по чистому недоразумению считают основателем отечественной палеогеографии. Вскоре и я стал обнаруживать, что идеи, возникшие, как считали коллеги, совсем недавно, весьма обстоятельно обсуждались еще в прошлом веке. Далекие предшественники стали быстро умнеть в моих глазах.
Сам по себе розыск предшественников современных идей — довольно азартное занятие. Восстановление приоритета доставляет историку науки не меньшее удовольствие, чем новое наблюдение в экспедиции или лаборатории. Нередко история науки становится для исследователя и источником ценных, но забытых идей. Однако высший смысл изучения старой литературы в ином. С какого-то момента начинаешь понимать, что наши нынешние взгляды не вершина творения научной мысли, а преходящий этап, что пройдут годы, и самые прочные убеждения могут подвергнуться сомнению, будут отброшены и забыты. И чем сложнее проблема, тем меньше шансов у господствующих взглядов удержаться надолго и тем более навсегда.
Обо всем этом я заговорил не случайно. Знание истории науки, ее уроков могло бы смягчить дискуссии и сделать их более плодотворными по одной из любопытнейших проблем, касающихся истории Земли, — проблеме перемещения континентов, или мобилизма. Еще не так давно, чуть больше десяти лет назад, в наших геологических кругах к мобилистам относились как к людям со странностями, занятным чудакам, которые вопреки очевидности исповедуют архаичные и никому не нужные взгляды. В лучшем случае к мобилизму относились как к «полезному для своего времени заблуждению». В книге по истории геологии, вышедшей в 1956 г., так и написано: «Появление даже таких в основном неверных представлений, как гипотеза перемещения материков Вегенера… принесло некоторую пользу науке, поскольку авторы этих представлений подвергли критике слабые места ранее господствовавших взглядов… и обратили внимание исследователей на некоторые факты, требовавшие объяснения…»
Идея горизонтальных перемещений земных масс на колоссальные расстояния «чрезвычайным насильством внутреннего подземного действия» обсуждалась во времена Ломоносова и даже раньше. К ней не раз обращались в прошлом веке, но восприняли всерьез лишь после книги А. Вегенера «Возникновение континентов и океанов». Известный геолог и климатолог, впоследствии трагически погибший в экспедиции в Гренландии и окруженный неким романтическим ореолом, провел идею движения материков через всю историю Земли. До сих пор публикуются составленные им палеогеографические карты, иллюстрирующие, как первичный гигантский материк Пангея стал расползаться в послепалеозойские времена на части, которые лишь в последние геологические эпохи сложились в современную картину распределения материков и океанов. Гипотеза Вегенера была обоснована обширным геологическим и палеонтологическим материалом. Его книга была переведена на многие языки и встретила широкое сочувствие, особенно среди геологов, работавших на «гондванских» материках. Потом интерес к мобилизму стал угасать, прежде всего среди европейских и американских геологов, так как предложить удовлетворительный физический механизм перемещения материковых масс не удавалось.
В 60-х годах отношение к гипотезе дрейфа материков снова изменилось. Накопились данные по остаточной намагниченности горных пород, по которой можно восстановить положение полюсов. Данные по каждому материку примерно совпадали, а расхождения можно было объяснить неточностью метода. Данные же по разным материкам давали настолько противоречивые сведения о положении полюсов, что для их согласования приходилось допускать крупные смещения в относительном положении материков в геологическом прошлом. Попытки списать это противоречие на ошибки измерений успокаивали противников мобилизма, но не убеждали сторонников. В случае же так называемых полосовых магнитных аномалий на океаническом дне на ошибки измерений уже сослаться было нельзя.
 Рис. 14. Сильно упрощенная схема перемещения материковых плит (по А. А. Гангнусу)
Рис. 14. Сильно упрощенная схема перемещения материковых плит (по А. А. Гангнусу)
Эти аномалии были обнаружены при магнитной съемке дна океанов. По обе стороны от срединно-океанических хребтов располагаются последовательные полосы с чередующимся знаком полярности. Ф. Вайн и Д. Мэтьюз предположили, что в этих полосах записаны последовательные фазы базальтового магматизма. Остывающая магма запечатлевала полярность магнитного поля и отодвигалась в сторону от оси хребта. Поколения магматических пород записывали смену в полярности геомагнитного поля, как на магнитофонной ленте. Появилась гипотеза, что в районе срединно-океанического хребта выходят глубинные породы, которые постепенно растекаются по обе стороны от хребта (рис. 14), Океаническое дно расширяется и соответственно разъезжаются материки, сидящие на вязкой подстилке — астеносфере. Плиты, из которых складывается земная кора, двигаются, расходятся, сталкиваются, дробятся, местами засасываются одна под другую, перекрываются, сминаются в горные цепи. Такая модель развития Земли была названа тектоникой плит.
Вскоре «тектоника плит» получила подкрепление. В конце 60-х годов удалось решить проблему глубоководного бурения. Океанское буровое судно «Гломар Челленджер» пробурило первые профили через Атлантику. Выяснилось, что чем ближе к Срединно-Атлантическому хребту, тем меньше мощность рыхлых осадков, ложащихся на вулканический фундамент, и все моложе породы в основании рыхлого осадочного чехла. В растекание океанического дна поверили многие противники мобилизма. Началось повальное увлечение составлением схем былого расположения материков. Неизменными элементами этих схем стали Гондвана и соединение материков, ныне разделенных Атлантическим океаном. В остальном схемы различаются очень сильно. Особенно досталось Мадагаскару, который на разных схемах «ездит» вдоль всего восточного побережья Африки. Индостанский полуостров то прислоняют к Африканскому Рогу, то отгоняют от него на тысячи километров. На одних картах Евразию раскалывают океаном, отделяющим Сибирь от Восточно-Европейской платформы. Другие карты сохраняют нынешнее положение этих частей Евразии.
«Тектоника плит» вызвала бурную дискуссию. Одни безоговорочно ее поддержали, другие решительно отрицали. Появились дочерние концепции, многочисленные варианты основных идей. Возродился интерес и к гипотезе расширения Земли, которая обещает снять некоторые противоречия между мобилизмом и фиксизмом. «Мобилистский бум» реабилитировал чудаков-мобилистов. К проблеме движения материков не осталась равнодушной и широкая публика. Оказывается, даже людям, далеким от науки, совсем не безразлично, живут ли они на устойчивых или путешествующих континентах. Тем более постепенно втянулись в дискуссию те, в чьих руках есть факты «за» и «против» дрейфа материков. Именно так случилось с палеонтологами. Одна из недавних годичных сессий Всесоюзного палеонтологического общества была посвящена теме «Палеонтология, палеобиогеография и мобилизм». И вот что примечательно. На этой сессии прямо высказались в пользу мобилизма 13 докладчиков из 20 вообще высказывавшихся на эту тему. Двадцатью годами раньше, в 1955 г., на такой же сессии общества, кстати тоже посвященной палеобиогеографии, мобилизма коснулись лишь три докладчика и все высказались против перемещения материков в решительных выражениях.
Если бы это явление было единичным, то оно вряд ли бы заслуживало внимания историка геологии. Однако оно далеко не единично. Просматривая палеонтологическую литературу последних десятилетий, можно прекрасно видеть, как многие палеонтологи, сначала убежденные «фиксисты», становятся не менее убежденными мобилистами. Например, в 1960 г. американский палеоботаник Д. Аксельрод опубликовал статью под громогласным заголовком: «Ископаемые флоры свидетельствуют об устойчивых, а не о дрейфующих материках». Немецкий палеоклиматолог М. Шварцбах тщательно проанализировал примерно тот же круг фактов и сформулировал противоположный вывод, воспользовавшись теми же словами. «Ископаемые флоры, — написал он, — свидетельствуют о дрейфующих, а не об устойчивых материках». В 1970 г. Аксельрод опубликовал статью, в которой безоговорочно присоединился к мобилизму, в частности к тектонике плит, и почему-то ни словом не обмолвился о своей же статье десятилетней давности и никак не объяснил причин изменения позиции.
Подобные перемены в настроениях хотелось бы объяснить поступлением каких-то новых материалов. Однако в самой палеонтологии такого не происходило. Новые материалы, конечно, поступали, но к дискуссии о перемещении материков они мало что добавляли. В основном люди меняли взгляды на ранее известные материалы. Старое представлялось в новом свете. Решающим, несомненно, был экспорт идей из геологии, из тектоники плит.
Можно упрекнуть Д. Аксельрода и других палеонтологов, «изменивших» фиксизму, в непоследовательности и недостаточно внимательном анализе имеющихся в их распоряжении фактов. Но можно повернуть дело иначе и задать себе следующие вопросы: если когда-то эти палеонтологи ошибались, то где гарантия, что они не ошибаются и теперь? И еще: кто доказал, что палеонтологические факты толковались сначала ошибочно, а потом верно? Может быть, все как раз наоборот? Возможно и такое, что палеонтологические факты вообще можно толковать и так и этак. Стало быть, они не обладают в дискуссии правом голоса. Если ископаемые фауны и флоры за уши притянуты к предмету спора, то не стоит ли палеонтологам вовсе воздержаться от коалиции, будь то с фиксистами или с мобилистами?
Если такие вопросы возникнут у тектониста и он захочет в них самостоятельно разобраться, то палеонтологическая литература быстро его разочарует. Во многих сотнях статей и целых книгах, посвященных теме «Ископаемые организмы и мобилизм» найдутся лишь единичные и весьма скупые соображения о том, каковы должны быть палеонтологические факты, чтобы их можно было привлекать для обоснования или опровержения мобилизма.
Обычно палеонтологи, выступающие за перемещение материков, обращают внимание на три группы фактов. В первую группу входят все случаи распространения одних и тех же ископаемых животных и растений на противоположных берегах океана. Когда в триасовых отложениях Антарктиды нашли кости листрозавра — рептилии, до этого известной в Индии, Южной Африке и Южной Америке, то в научных изданиях и даже в газетах замелькали сообщения, что получено замечательное доказательство объединения материков южного полушария и Индии в некогда существовавшую Гондвану и последующего расползания ее частей.
В прошлой главе уже говорилось о поразительном сходстве палеозойских растений во всех частях Гондваны — факт, на который опирался и Вегенер. Можно допустить, что не раз и делали, постоянство в расположении гондванских материков и сушу на месте нынешних южных океанов. По ней, дескать, и путешествовали листрозавры и легко расселялись растения. Тогда, правда, придется допустить, что эти растения и рептилии совершенно не зависели от географических поясов и могли жить на всех широтах — от Северного тропика до Южного полюса. Опровергнуть такое предположение трудно, но не легче его и доказать. Подавляющее большинство палеонтологических фактов, привлекаемых в пользу мобилизма, относится именно к этой группе. Нашли по обе стороны от Атлантики одинаковые морские раковины. Вот и доказательство, что Африка соединялась с Америкой. Ясно, однако, что морские раковины никоим образом не могут дать такого свидетельства. Впрочем, и с наземными организмами, например растениями, дело не обстоит так просто, ведь их споры или семена могли путешествовать через морские преграды. Еще Дарвин интересовался вопросом, насколько вероятен перенос семян птицами через океаны. Он счищал грязь с птичьих лап, находил в ней семена, проращивал их и тем самым доказывал, что птицы — важное транспортное средство в расселении растений. Было показано, что мелкие семена могут переноситься на лапках и некоторых насекомых (например, саранчи). Известно, что мелкие зверюшки, спасаясь от наводнений, путешествуют на вынесенных реками бревнах. Нередко реки несут в моря целые плавающие острова из торфяной залежи со всем населением — растительным и животным. Причалит такой остров к другому материку и завезет туда целое сообщество иноземцев.
Итак, эта группа фактов указывает на неестественную разобщенность фаун и флор, разорванность ареалов родов и видов, которую хочется убрать, осушив разделяющие их океаны. Такие факты действительно могут косвенно свидетельствовать о времени возникновения океанических впадин, но далеко не всегда эти свидетельства надежны. Действительно, вообразим себе палеонтолога далекого будущего. Он найдет кости лошадей, собак и кошек по обе стороны нынешнего Атлантического океана и сделает вывод, что в XX в. Атлантики не было. Хотя организмы геологического прошлого не путешествовали на кораблях и самолетах, но, как мы видели, у них были свои транспортные средства, пусть и не регулярные.
Итак, эта группа фактов, наиболее популярная среди палеонтологов-мобилистов, дает лишь, косвенные свидетельства. Именно эти факты легче всего допускают встречное толкование, и именно они чаще всего сбивают людей с толку. Убедительнее кажется другая группа фактов. Их суть лучше пояснить на конкретном примере. В отложениях мелового периода Северо-Восточной Бразилии и в одновозрастных толщах той части Экваториальной Африки, которая обращена к Атлантике, встречаются очень сходные раковины мелких ракообразных (остракод). Примечателен не сам по себе общий состав видов остракод, а то, в каком окружении они захоронялись. В нижней части геологического разреза они приурочены к осадкам сравнительно небольшого пресноводного бассейна. Выше по разрезу на обоих континентах отложения этого бассейна сменяются одинаковыми соленосными толщами с характерным и довольно редким составом солей. Еще выше и в Бразилии, и в Африке лежат морские отложения с одними и теми же раковинами аммонитов. Эта аммонитовая фауна резко отличается от известной в Африке же, но севернее (в Марокко). Еще выше по разрезу аммониты становятся одними и теми же по всей приатлантической части Африки.
Палеонтологи Р. Реймент и Э. Тейт рассказывали про все это на Международном геологическом конгрессе в 1972 г. В начале мелового периода, считают они, Бразилия почти вплотную примыкала к Экваториальной Африке. Пресноводный бассейн, в котором жили остракоды, захватывал смежные части обоих континентов. Потом с юга внедрилось море с аммонитами, что ознаменовало начало расхождения материков. Еще позже морской бассейн расширился и соединился с другим бассейном, располагавшимся на севере Африки, Образовалось подобие нынешнего Атлантического океана. В последние годы было проведено сравнение комплексов спор и пыльцы из отложений, описанных Рейментом и Тейтом. Оказалось, что и здесь можно видеть удивительное сходство бразильских и африканских (габонских) комплексов, и тоже не единичных, а образующих одну и ту же последовательность, причем отличающуюся от того, что известно в других местах Африки и Южной Америки.
Изложенное отличается от первой группы фактов тем, что на разных материках сходны не только организмы, но и вмещающие осадки и сходны следы не каких-то отдельных событий, а вся их последовательность в районах, ныне разделенных тысячами километров океана. Можно, конечно, и здесь допустить, что одинаковые события происходили в Бразилии и Африке независимо (или зависели от какой-то общей причины) и что из-за сходства разрезов и окаменелостей совершенно не обязательно двигать материки. Все же такое допущение кажется менее вероятным.
Слова «менее вероятным» употреблены здесь не из-за внешней осторожности. Приходится помнить, что восстановление давно прошедших событий всегда основано на неполном знании фрагментарных документов и что многие следы утрачены навсегда. Делать категорические выводы в таких условиях не только рискованно, но и недопустимо. Впрочем, об этом более подробно пойдет речь в
главе IX.
Наконец, есть третья группа фактов, доставляемых главным образом палеоботаниками и ставящих противников мобилизма в особенно трудное положение. Наверное, поэтому они стараются о таких фактах не вспоминать. Ископаемые растения дают много ценнейших свидетельств о климатах прошлого, в чем читатель уже успел убедиться.
Движение материка вдоль географических параллелей может не повести к существенному и направленному изменению его климата. Такое движение по палеоклиматическим данным уловить трудно или невозможно. Иное дело, когда материк поползет поперек параллелей и будет пересекать климатические зоны. Тогда будет направленно меняться его растительность и становиться все более теплолюбивой или холоднолюбивой. Наблюдения по одному материку недостаточны, поскольку изменение его климата можно связать и с перемещением полюсов при устойчивом относительном положении материков. Иное дело, если климаты на разных материках будут меняться несогласованно, так что наблюдения по одному материку потребуют сдвижения полюсов в одном направлении, а по другому материку — в другом. Впрочем, и этого недостаточно, Надо еще убедиться, что климат менялся именно из-за пересечения ползущим материком географических параллелей, а не по иной причине. Всем известна зависимость климата не только от широты, но и от побочных факторов — течений, холодных или теплых, горных хребтов, крупных водоемов и пр.
 Рис. 15. Современное распространение мангровых зарослей; пунктиром показаны Северный и Южный тропики
Рис. 15. Современное распространение мангровых зарослей; пунктиром показаны Северный и Южный тропики
Все это в общем учитывали участники дискуссии о мобилизме. Достаточно было палеоботаникам указать на ненормально широкое распространение глоссоптериевой флоры Гондваны (от Южного полюса до северной части Индии), как сразу появились объяснения, как могли океанические холодные течения остужать Гондвану на севере, а теплые — подогревать у полюса. Палеоботаники вместе с литологами отметили смещение к северу климатического экватора в позднем палеозое и в ответ быстро получили схему возможной циркуляции водных и атмосферных масс, ответственной за аномалию.
Мнение, что из-за асимметричного расположения материков и океанов на поверхности Земли, как и из-за течений и горных хребтов, климатические зоны не идут параллельно широтам и вообще не обнаруживают полной симметричности, совершенно справедливо. На юге тропики быстро сменяются холодным антарктическим климатом, а на севере за тропиками следует широкая полоса субтропиков и умеренно теплого климата. Отсутствие строгой параллельности современных климатических зон не раз фигурировало в качестве контраргумента, когда сторонники мобилизма использовали причудливые очертания палеоклиматических областей для подкрепления мобилистских идей. Однако ни сторонники, ни противники мобилизма не учли важного обстоятельства, а именно того, что асимметрия современных климатических зон почти не затрагивает экваториальной полосы теплого, безморозного климата без существенных сезонных колебаний.

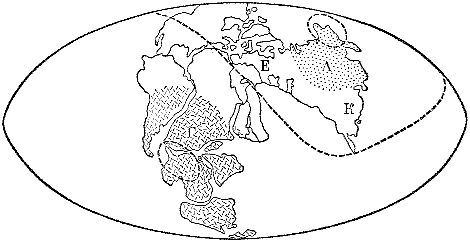 Рис. 16. Расположение главных палеофлористических областей в позднем палеозое при современном положении материков (наверху) и на мобилистской реконструкции (внизу); А — Ангарское царство; Е — Еврамерийская область, К — Катазиатская область; Г — Гондванское царство
Рис. 16. Расположение главных палеофлористических областей в позднем палеозое при современном положении материков (наверху) и на мобилистской реконструкции (внизу); А — Ангарское царство; Е — Еврамерийская область, К — Катазиатская область; Г — Гондванское царство
Такой климат особенно характерен для тропических дождевых лесов и для океанических побережий, окаймленных полосой мангровых зарослей (рис. 15). Полоса этого климата (условно назовем его экваториальным) местами сужается или расширяется, в нее заходят горные системы, на вершинах которых климат может быть суровым. И нигде экваториальный климат не отходит от географического экватора больше чем на 30–35° по меридиану. Напрашивается вывод, что полоса экваториального климата искажается воздействием местных факторов, но сместиться так, чтобы лечь целиком по одну сторону экватора, не может. Полагают, что строгая приуроченность экваториальной полосы к географическому экватору контролируется астрономическими причинами: положением земной оси по отношению к эклиптике, солнечной радиацией.
Сейчас уже накоплено немало данных о растительности экваториальной полосы в последние геологические эпохи. Выяснилось, что эта полоса сужалась и расширялась в эпохи похолоданий и потеплений, так и оставаясь более или менее симметричной по отношению к экватору. Напрашивается мысль проследить историю зоны экваториального климата в геологическом прошлом и сопоставить данные по разным материкам. Такая работа была проделана. В позднем палеозое экваториальному климату соответствовали уже упоминавшиеся еврамерийская и катазиатская флоры. Они занимали всю Северную Америку, Гренландию, почти всю Европу, север Африки. Далее на восток область распространения экваториальной флоры внезапно сужалась в несколько раз и в Средней Азии едва достигала тысячи километров в поперечнике, а потом в Китае и Юго-Восточной Азии снова расширялась в несколько раз (рис. 16).
Даже не это странное очертание в виде песочных часов — главная особенность позднепалеозойского экваториального пояса. Любопытнее то, что весь он лежит в пределах современного северного полушария и лишь небольшими выступами заходит за экватор на юг. Можно попытаться объяснить обе аномалии течениями и циркуляцией атмосферных масс (хотя этого пока никому не удавалось), но тогда важно не забыть еще об одном парадоксе. Прослеживая северную границу экваториального климатического пояса за время от начала среднего карбона до середины мела, можно видеть, что она несколько колеблется, спускаясь на юг или поднимаясь к северу, так и не покидая сравнительно узкой полосы порядка тысячи километров в поперечнике. Впрочем, важна не ширина этой полосы, а то, что движение северной границы не было направленным (рис. 17).
 Рис. 17. Положение северной границы экваториального пояса в разные эпохи — от позднего палеозоя до середины мела (за 250 млн. лет)
Рис. 17. Положение северной границы экваториального пояса в разные эпохи — от позднего палеозоя до середины мела (за 250 млн. лет)
Аналогичная южная граница ведет себя совсем по-иному. Лучше всего это видно на меридиане Индии. В позднем палеозое Индия принадлежала Гондване с ее внетропическим, временами даже холодным климатом. В конце мезозоя Индия уже была заселена типично тропической растительностью. Если нанести на карту положение южной границы экваториального пояса на долготе Индии, то мы увидим ее последовательное смещение на юг на несколько тысяч километров. Такая же картина получится и на меридиане, проведенном вдоль Африки или Южной Америки. Экваториальная полоса заняла современное положение уже в кайнозое. Если принять вывод, что экваториальная полоса контролируется прежде всего астрономическими причинами, то понять описанную историю ее северной и южной границ без обращения к перемещению материков становится невозможным.
У человека, скептически относящегося к палеоботаническим свидетельствам, может возникнуть вопрос: а насколько надежны реконструкции палеоклиматов по ископаемым растениям? Может быть, те признаки, на которые опираются палеоботаники, зависят от климата лишь у современных растений и имели к нему косвенное отношение в далеком прошлом?
Думаю, что любой отдельно взятый признак растения не дает окончательного доказательства определенного типа климата. Тем более нельзя ручаться, что раньше признаки растений не могли быть иначе связаны с климатическими условиями, чем сейчас. Например, в позднем палеозое во флорах Ангариды и Гондваны преобладали растения с цельными, нерасчлененными листьями (глоссоптериевые в Гондване, кордаиты в Ангариде). В экваториальном же поясе, т. е. в местах распространения еврамерийской и катазиатской флор, преобладали растения с сильно расчлененной листовой пластинкой — различные папоротники и птеридоспермы («семенные папоротники»). Соотношение цельных и расчлененных листьев в позднем палеозое в разных климатических зонах было, таким образом, противоположным тому, что мы видим в современной растительности.
В чем здесь дело? На этот вопрос нет ответа. Конечно, можно привести разные вспомогательные гипотезы и, например, напомнить, что у ангарских кордаитов нередко бывает мелкозубчатый край на верхушке листа, а у еврамерийских кордаитов такого пока неизвестно. На это соображение можно возразить,
что в Еврамерийской области ранней перми зубчатый край свойствен некоторым клинолистникам, а у ангарских раннепермских клинолистников край цельный.
Вообще палеоботаникам следовало бы всегда помнить о том, что экологический смысл большей части признаков растений совершенно неизвестен. Мы не знаем, почему растения именно так, а не иначе реагируют на климат; часто не знаем, связано ли какое-то изменение признаков у растений разных флористических областей с климатом или с иными факторами.
В главе II уже говорилось, что приуроченность многолетних маноксилических стволов к областям безморозного климата физиологически непонятна. Для нынешних тропиков характерна каулифлория, когда цветки не сидят на ветвях дерева, а прикрепляются прямо к главному стволу. Примерно тысяча видов, имеющих каулифлорию, живет в нынешних тропиках, а во внетропических зонах такие растения редки (к ним относится, например, «волчье лыко»). У некоторых еврамерийских плауновидных карбона шишки (стробилы) сидели прямо на стволе, и это послужило дополнительным аргументом для отнесения Еврамерийской области к тропикам. Чем же вызывается каулифлория, мы так и не знаем. Замечено, что значительная доля растений с каулифлорией опыляется летучими мышами, и даже высказывались предположения, что именно способ опыления ответствен за появление цветков на стволах. Очевидно, для палеозойских плауновидных, не нуждавшихся в опылении, надо искать другое объяснение. Р. Потонье полагал, что каулифлория помогала защищать органы размножения от тропических ливней. Кто возьмется доказывать или оспаривать эту гипотезу?
Случай с климатической интерпретацией каулифлории, маноксилических стволов и края листа (цельного или расчлененного) довольно типичен. Как правило, мы даем признакам растений (и вообще организмов) то или иное экологическое толкование, основываясь не на знании действительного приспособительного значения признака, а лишь на частом совпадении признака и какого-то фактора среды. Мы не знаем, какие преимущества имеют растения с цельными листьями в тропическом лесу, а просто видим эмпирическое совпадение двух явлений во многих случаях. Установление совпадений, не подкрепленных знанием более глубоких причин, видимо, не стоит называть закономерностью, а лучше сопоставить с народными приметами. Жители средней полосы России знают, что ясное летнее утро с обильной росой — к хорошей погоде, а почему так, не имеют представления (думаю, что и познания метеорологов по части толкования народных примет не слишком велики, иначе мы с меньшим сарказмом относились бы к прогнозам погоды, шокирующим иной раз и самих синоптиков). Ботаник, связывающий те или иные признаки с климатом, поступает примерно так же: опирается на внешние приметы и лишь гадает о причинах.
В немногих случаях палеоботаник, восстанавливающий климат по структуре растений, может опереться на более солидные закономерности. Таково строение древесины в многолетнем стволе. Зависимость прироста древесины от климатических факторов и внутренних ритмов растения изучена неплохо. Если клетки древесины не складываются в слои прироста (годичные кольца), то можно довольно уверенно заключить, что ствол рос в условиях очень выравненного климата с высокими среднегодовыми температурами (выше 15°). Клетки камбия, производящие древесину ствола, чутко реагируют на сильные сезонные колебания температуры и влажности и обязательно откликаются на сезонный листопад. Их функция почти подавляется постоянными низкими температурами, даже плюсовыми. Не случайно на океанических островах высоких широт, где климат целый год очень ровный, но холодный (всего несколько градусов тепла, а морозов не бывает), деревья не растут вообще. Зная все это и найдя толстый древесный ствол, в котором будут плохо заметны или вовсе неразличимы слои прироста, палеоботаник может уверенно заключить, что климат был не только с минимальными сезонными колебаниями (безморозный), но и очень теплый, близкий к тропическому.
Для отложений последних геологических эпох надежные сведения о климате можно получить, просто анализируя состав родов и видов, остатки которых удалось надежно определить. Спускаясь вниз по геологическому разрезу, мы должны становиться все более осторожными, поскольку виды и тем более роды могли менять свои климатические привязанности. Не исключено и то, что распространение некоторых растений контролируется сейчас не только климатом, как может показаться при поверхностном анализе «примет», а, скажем, фотопериодом — длиной дня и ночи в течение года. Если растение способно жить в широком диапазоне среднегодовых температур, но строго зависит от фотопериода, то ошибка палеоботаника почти неизбежна. Найдя остатки растения, свойственного сейчас умеренным широтам из-за их фотопериода, а не из-за среднегодовых температур, палеоботаник сделает вывод, что климат был умеренным, хотя температуры, в которых жило раньше это растение, были под стать тропическим. Я не исключаю, что такие ошибки делаются не так уж редко. Например, в палеогеновых (эоценовых) отложениях вблизи Лондона есть толща глин («лондонских») с большим количеством растительных остатков, особенно семян. Многие семена принадлежат типично тропическим растениям, но есть и роды, характерные сейчас для внетропических флор. Палеоботаники считают, что климат этих мест в эоцене не был тропическим, хотя и был безморозным.
Для эоцена Европы имеются данные геохимических палеотемпературных измерений, и они указывают на высокие, по существу тропические среднегодовые температуры омывавших Англию морей. Думается, что геохимическим данным следует доверять больше, чем родовому составу ископаемых растений. Сейчас палеотемпературные измерения по изотопному составу изотопов кислорода и по соотношению кальция, магния и стронция в морских раковинах накопились уже в большом количестве, и их надежность подтверждена на многочисленных современных животных. Зная температуру вод, омывающих сушу, можно составить представление и о климате побережья, а затем экстраполировать полученные данные в глубь суши. Замечу, что установить направление климатических изменений, будь то в пространстве или во времени, по комплексам растительных остатков много легче, чем сказать, какие конкретно были на данном участке климатические условия. Палеоботаник часто может указать, что климат похолодал, но не может сказать, о скольких градусах идет речь.
Все эти сложности необходимо принимать во внимание, поскольку они исключают подход к палеоклиматическим реконструкциям как к некоей механической операции: зафиксировал несколько первых попавшихся признаков и вывел температуру и влажность. Приходится учитывать все множество собственно палеоботанических данных, сопоставлять их с наблюдениями над вмещающими породами, смотреть связь комплексов растительных остатков в смежных частях разреза и в соседних районах. Если так подходить к палеоклиматам, то получаются реконструкции достаточно надежные, чтобы с ними считались и противники и сторонники мобилизма. Отвести согласованный комплекс палеоклиматических свидетельств уже нельзя только потому, что отдельные компоненты анализа не вполне надежны. Все те палеоклиматические данные, которые приводились выше в поддержку гипотезы мобилизма, основаны именно на таком сложном комплексе признаков. Поэтому к ним целесообразно относиться если и не с полным доверием, то с уважением.
Хотелось бы напомнить и о других палеоботанических фактах, которые успели отрастить бороду в ожидании внимания со стороны противников мобилизма. Прежде всего это палеоботаническая летопись Индии. Фиксисты единодушны в том, что Индия для них самый крепкий орешек. Внетропический и, может быть, даже холоднолюбивый характер индийской флоры позднего палеозоя и тропический характер меловой и кайнозойской ее флоры прекрасно согласуются именно с перемещением Индии из высоких южных широт на север, причем никаким перемещением полюсов здесь помочь нельзя. И характер индийской флоры, и ее связь с отложениями, оставленными ледниками, не раз пытались объяснить тем, что оледенение было горным и растения жили высоко над уровнем моря. Фиксистам не раз напоминали, что в толще с глоссоптериевой флорой есть по крайней мере два морских прослоя. Комплексы спор и пыльцы в отложениях, занимающих как морские, так и ледниковые толщи, примерно одни и те же. Не могла индийская флора жить на горах. Возражения серьезные, а ответа на них не дают.
Индийский палеоботаник Б. Сахни еще в 30-х годах подметил необычно резкую границу между индийской частью Гондваны и смежными палеофлористическими областями. Последующие исследования лишь подтвердили наблюдения Сахни. На крайнем востоке Индии, где Брахмапутра делает крутой поворот и сбегает с Гималаев, найдены типично гондванские растения. В нескольких сотнях километров к востоку, в китайской провинции Юньнань, тоже известны верхнепалеозойские растения, уже чисто катазиатские, тропические. Восточноиндийская флора росла совсем близко от тропического океана Тетис, если считать, что Индия всегда сидела на своем месте. Никакого влияния Тетиса на эту флору не видно, хотя, судя по анализу верхнепалеозойских речных долин, реки текли из области глоссоптериевой флоры именно в направлении Тетиса и на их пути не было мощной горной системы, которая могла бы закрывать Индию от теплого дыхания океана.
Можно указать и другие места, где палеофлористические границы очень резкие и для обычной переходной зоны между областями почти не остается места. На крайнем юго-востоке Монголии пермская флора типично ангарская, а в соседнем Китае, у выступающей на север излучины Хуанхэ, — чисто катазиатская, тропическая. Еще ближе сошлись ангарская и катазиатская флоры в Приморье. Здесь между ними осталось не более 200 км. Разница же флор огромная, Чтобы понять суть таких резких границ, представьте себе, что вы садитесь на поезд в Москве, сначала едете мимо березовых рощ, потом поспали, а примерно у Тулы открыли глаза и увидели за окном бананы и пальмы.
Важно не забывать о том, что палеоботаник преимущественно имеет дело с ассоциациями растений, живших в низинах, лишь немного приподнятых над уровнем моря. Только в таких низинах велась в позднем палеозое и мезозое дошедшая до нас геологическая летопись континентов. Поэтому связывать различие флор с тем, что они жили на существенно разных высотах, в большинстве случаев не приходится. И тогда напрашивается мысль объяснить резкие флористические границы тем, что блоки земной коры сдвинулись и поместили рядом местонахождения растений, когда-то живших вдали друг от друга.
Идея о связи резких флористических границ с горизонтальными тектоническими перемещениями не раз приходила в голову палеоботаникам. Однако было бы преждевременным отклонять другие объяснения. До сих пор палеофлористические границы остаются очень плохо изученными. Мало собрать списки растений и нанести на карту местонахождения растительных остатков. Надо восстановить всю палеогеографическую обстановку пограничных территорий и проследить ее изменения за длительный промежуток времени. В некоторых случаях резкие границы явно никак не связаны с горизонтальными тектоническими движениями. Раньше уже упоминались «лондонские глины» с тропическими растениями. Там же, в Англии, в нескольких сотнях километров северо-западнее, есть иные местонахождения, растения которых уже не назовешь тропическими и даже субтропическими. Их долго считали более молодыми, но сейчас скапливается все больше данных, что вмещающие их толщи примерно одновозрастны «лондонским глинам». Почему Англию в эоцене пересекала столь резкая палеофлористическая граница, сказать пока невозможно.
Напоследок расскажу о недавнем палеоботаническом исследовании. Оно, надо полагать, доставит меньше всего радости тем; кто решил не сдавать фиксистских позиций. В 1973 г. экспедиция наших геологов начала работы в Исландии, которая сидит прямо на Срединно-Атлантическом хребте. Если «растекание» океанов от срединных хребтов действительно существует, то можно ожидать тот же процесс и в Исландии, где его наблюдать гораздо удобнее, чем на морском дне. Для реконструкции новейшей истории острова (а докайнозойских пород тут нет вообще) важно было прежде всего как следует разобраться в возрасте слагающих остров вулканических пород — покровов лав и туфовых толщ.
К счастью, в вулканогенно-осадочных отложениях острова встречаются растительные остатки, которыми занялся палеоботаник М. А. Ахметьев. Он нашел много горизонтов с растениями и, кроме того, привез образцы для спорово-пыльцевого анализа, в которых палинологи нашли достаточно богатые комплексы спор и пыльцы. Когда палеоботанические документы были размещены в геологическом разрезе, а затем был составлен стратиграфический профиль через весь остров, то картина получилась такой. Вдоль меридиональной оси острова, совпадающей с осью Срединно-Атлантического хребта, слоев с растительными остатками не оказалось вовсе. Чем дальше в стороны от этой линии, тем все более мощным был разрез и все древнее были слои с растениями, лежавшие в основании разреза. Иными словами, получилась точно та же картина, что и нарисованная в результате глубоководного бурения (рис. 18). Только теперь для датировки пород использовались остатки не морских организмов, а наземных растений.
Интересными оказались и биогеографические связи территории Исландии. М. А. Ахметьев заметил, что наиболее древние комплексы растений близки к одновозрастным комплексам Гренландии и Северной Америки. Чем выше по геологическому разрезу, тем все большую роль играют европейские растения. Современная флора Исландии уже по существу европейская. Это наблюдение, может быть, указывает, что расширение Атлантики в обе стороны не было сбалансированным.
Палеоботанические данные уже не раз использовались для реконструкции истории Атлантического океана. О гондванских флорах мы уже говорили. В. А. Вахрамеев сравнивал флоры, жившие по берегам Атлантики в мезозое, и пришел к выводу, что возрастание различий европейских и американских флор в меловом периоде можно связать с образованием здесь океана, Д. Хабиб изучал палинологические комплексы в меловых отложениях севернее Багамских островов и обнаружил в морских осадках такое огромное количество спор и пыльцы (более 40 тыс. зерен на грамм осадка) и такое их разнообразие, которое возможно только в том случае, если берег находился совсем рядом. Эти данные прекрасно согласуются с мобилистским прочтением истории Атлантики, но при большом воображении их можно истолковать и с фиксистских позиций. Истолковать же историю Исландии, не обращаясь к гипотезе раздвигания блоков земной коры в обе стороны от срединного хребта, по-видимому, невозможно.
 Рис. 18. Последовательность флор, населявших Исландию с начала миоцена (по М. А. Ахметьеву)
Рис. 18. Последовательность флор, населявших Исландию с начала миоцена (по М. А. Ахметьеву)
Читатель, может быть, упрекнет меня за непрерывные нападки на фиксистов, за град упреков в их адрес. Человек, знакомый с дискуссией о движении материков, может быть, добавит, и будет прав, что и мобилисты не лучше обращаются с неприятными для них фактами. В свое оправдание скажу только, что я в данном случае выступал как палеоботаник и попрекал фиксистов в невнимании именно к палеоботаническому материалу.
Глава VII
КОРНИ МЕЗОФИТА

В книге П. Эрлиха и Р. Холма «Процесс эволюции» есть выражение, которое мне сразу очень понравилось, и я его потом не раз использовал. Они разбирают известный «биогенетический закон» Э. Геккеля, гласящий, что онтогенез повторяет филогенез, т. е. каждый организм в индивидуальном развитии проходит стадии эволюции предков. Эрлих и Холм пишут, что на недостатки этой идеи, заставляющей каждый организм снова взбираться на свое филогенетическое древо, указывают почти все современные исследователи, а она все же «занимает видное место в биологической мифологии». Перед нами действительно биологический миф, это не просто ошибка, а скорее заблуждение от недостатка компетентности, прочно удерживающееся в поколениях. Мифический «биогенетический закон» лишь один пример, число которых можно легко умножить. Приходится признаться, что в науке мифы играют немалую роль.
Я не берусь объяснять причину их возникновения и поразительной устойчивости. Возможно, причина отнюдь не в инертности и не в неумении критически относиться к прочитанному и услышанному, а просто в самой сложности науки. Ученый не в состоянии переосмыслить и проверить каждое наблюдение и каждое обобщение. Что-то приходится брать на веру, полагаясь на компетентность коллег. Броские, но упрощенные обобщения особенно быстро распространяются и легко запоминаются, Когда потом оказывается, что на самом деле все сложнее, то такое усложненное представление уже труднее распространяется за пределы круга компетентных специалистов. Заблуждение, старая точка зрения не вытесняется новым, усложненным взглядом. Примерно так произошло и с «биогенетическим законом». Современные представления о соотношении онтогенеза и филогенеза уже не уложить в простенькую формулу. Естественно, что они не смогли вытеснить «биогенетический закон», к тому же якобы подтверждаемый таким замечательным и всем известным фактом, как зачатки жаберных щелей у человеческого зародыша.
О некоторых мифах уже шла речь выше (например, о климатах карбона и перми, описываемых в школьном учебнике биологии). О других пойдет речь в этой и следующей главе. Читатель помнит, что в становлении геологии и биологии немалую роль сыграло то, что основной материал для наблюдения сначала был европейским. Все европейское долго казалось, а отчасти кажется и до сих пор типичным, закономерным, а все за пределами Европы — или дополнением к правилу, или исключением из него.
Еще в прошлом веке были выделены палеозойская, мезозойская и кайнозойская эры, причем решающую роль в их выделении сыграла палеозоология. Палеозой — время трилобитов, панцирных рыб, древнейших четвероногих. Мезозой — время динозавров, ихтиозавров, аммонитов и белемнитов. Кайнозой — время млекопитающих. Со времен Дарвина и Лайеля в книгах изображают международную стратиграфическую шкалу, где слева приводят периоды, эпохи и века, а справа перечисляют именно эти основные группы организмов. Что касается растений, то о палеозое принято говорить как о времени споровых растений — папоротников, древовидных каламитов и плауновидных, о мезозое — как об эре голосеменных, кайнозой считают эрой покрытосеменных.
Впрочем, еще в прошлом веке возникла неувязка в сопоставлении эр животной и растительной жизни. Европейская пермь делится на две части. В ранней перми растительный мир еще сохранял многие черты предшествующего каменноугольного периода. В поздней перми главную роль играли хвойные, внешне сходные с триасовыми, т. е. мезозойскими. Споровых растений в поздней перми Европы очень мало, и среди них нет ни каламитов, ни древовидных плауновидных. Поздняя пермь по растениям казалась гораздо ближе к мезозою, чем к ранней перми. Не менее ярко несоответствие на рубеже мезозоя и кайнозоя. Если флора раннего мела еще типично мезозойская, то в позднем мелу Европы господствовали покрытосеменные, так что облик захоронений растительных остатков типично кайнозойский.
Так появилось убеждение, что деление истории Земли по эволюции животных и растений различно и что крупнейшие этапы эволюции растений на полпериода опережают этапы в эволюции животных. Отсюда следовали важные выводы. Во-первых, надо вводить самостоятельные эры для растений. Их назвали палеофитом, мезофитом и кайнофитом. Во-вторых, поскольку этапы эволюции растений меняются раньше, чем у животных, в смене первых можно было видеть причину, а в смене вторых — следствие. В цепях питания первыми меняются растения, затем меняются травоядные, а за ними и плотоядные животные. На перераспределение цепей питания и уходит полпериода. Известный палинолог Р. Потонье даже провозгласил соответствующий закон «эволюционного несогласия».
Закон «эволюционного несогласия» был сформулирован сравнительно недавно, в 50-х годах, и лишь подвел некоторый итог взглядам, сложившимся задолго до этого, в 60 — 70-х годах прошлого века. В нынешних статьях и книгах, в том числе и учебниках, можно прочесть примерно такое же изложение истории органического мира. Если же внимательно ознакомиться с фактическими материалами, то чуть ли не все в этой картине оказывается неверным, причем материалов для исправления ошибок было вполне достаточно еще 20–30 лет назад. В этой главе я постараюсь развенчать мифы о событиях на рубеже палеозоя и мезозоя, а в следующей главе мы поговорим о том, что произошло на рубеже мезозоя и кайнозоя.
Начнем с того, что считать палеозой временем расцвета и господства споровых растений можно было лишь тогда, когда все растения с папоротниковой листвой принимались за настоящие папоротники и когда не были известны ангарская и гондванская флоры. Теперь хорошо известно, что очень многие палеозойские растения с листвой папоротникового типа, характерные для европейского карбона и ранней перми, имели семена и принадлежат к «семенным папоротникам» (птеридоспермам). Само слово «папоротники» в их названии сохраняется по традиции, подобно тому как один из лишайников мы по-прежнему зовем оленьим мхом. В Ангариде и Гондване в отличие от Европы уже со среднего-позднего карбона главную роль играли голосеменные, а роль споровых была второстепенной. Поэтому если бы мы членили историю растений применительно к Сибири, то эру голосеменных пришлось бы начинать со среднего карбона. С другой стороны, для мезозоя Сибири особенно характерно большое количество папоротников — их здесь не меньше, чем голосеменных. Вообще для Сибири становится неверным характеризовать палеозой как время споровых, а мезозой — как время голосеменных.
Я убежден, что выделять некие планетарные эры в эволюции растений — довольно безнадежное предприятие. Невозможно подстричь под гребенку единых этапов все растительное население Земли. Правящие династии растений менялись от одной области к другой, и все подгонять под европейский стандарт так же неуместно, как привязывать историю европейских народов к смене династий китайских императоров. Поэтому сами понятия «палеофит», «мезофит» и «кайнофит», если их сохранять, наполняются существенно разным содержанием, когда мы говорим о Сибири, Европе или Индии.
Еще более сомнителен вывод, что эволюция растений опережает эволюцию животных. Цепи питания — важный компонент эволюции органического мира. Наверное, многие эволюционные события прямо или косвенно связаны с тем, кто кого ел. Только я не думаю, что мы можем ставить в прямую связь широкое расселение покрытосеменных в середине мелового периода и вымирание аммонитов и белемнитов в конце мела. Если это звенья одной цепи питания, почему-то разматывавшейся несколько десятков миллионов лет, то неплохо было бы указать хотя бы главные элементы этой цепи. Тогда можно всерьез говорить и о гипотетическом законе «эволюционного несогласия».
Обсуждая общие закономерности эволюции органического мира, исследователи часто опирались на некие обобщенные списки растений и животных, составленных чуть ли не для всей Земли в целом, отвлекаясь от тех конкретных ассоциаций животных и растений, с которыми мы встречаемся в отдельных биогеографических областях. До сих пор никто как следует не сопоставил соотношение ассоциаций наземных травоядных позвоночных и комплексов растительных остатков того же района. Мы пока не имеем представления о том, какими растениями питались палеозойские и мезозойские травоядные и какие хищники им досаждали. С другой стороны, можно предполагать, что диета травоядных, относившихся к одному семейству и близкородственным родам, но жившим в разных палеофлористических областях, различалась достаточно сильно. Скажем, травоядные позвоночные поздней перми и раннего триаса, жившие в Гондване и Западной Ангариде, — довольно близкие родственники, но питаться им приходилось совсем разными растениями. Поэтому разумно предположить, что представители одной систематической группы травоядных могли питаться разными растениями не только в пространстве, но и во времени, т. е. существенно менять диету без большого ущерба для здоровья.
Многие из тех, кто писал о мезозойском облике позднепермской флоры и относил ее поэтому к мезофиту, не учитывали источника этого представления. Оно возникло после изучения позднепермской флоры Западной Европы. Верхнепермские растительные остатки захоронены здесь в медистых сланцах мощностью всего в несколько десятков сантиметров. Большей частью это остатки четырех родов хвойных, ни один из которых не известен в триасе. Нередко встречается один вид птеридосперма Lepidopteris, другие виды которого характерны для триаса. Остальные растения медистых сланцев редки, плохо изучены, и ничего определенного об их связях с мезозойскими растениями сказать нельзя.
Упрощенными оказались и представления о возрасте флоры медистых сланцев Европы. Они принадлежат «цехштейну» (средневековые рудокопы назвали так толщу морских отложений, составляющих верхнюю часть европейской перми). После того как были изучены микрофауна и палинологические комплексы цехштейна, оказалось, что он (а тем самым и вся европейская верхняя пермь) соответствует лишь верхним частям верхней перми в других областях. Короче говоря, если придерживаться обычных взглядов на объем верхней перми (учитывая и расхождения во взглядах), то так или иначе получается, что флора цехштейна, раньше олицетворявшая верхнепермскую флору вообще, всего лишь некая частная флора, распространенная только в Европе и только в конце перми, к тому же составленная родами или вымирающими в конце перми, или неизвестно как связанными с мезозойскими родами. Только один вид цехштейновой флоры действительно связывает ее с мезозоем.
В который уже раз мы сталкиваемся с тем, что новые исследования, вместо того чтобы дополнять и подправлять устоявшиеся взгляды, подтачивают их и наконец обращают в миф. Мы многое узнаем попутно, а от главного вопроса, на который хотели получить ответ, оказываемся дальше, чем думали в начале работы. Достаточно уверенно можно говорить только о совершенных в прошлом ошибках. Главная уже упоминалась. Изучая эволюцию растительного мира, нельзя механически суммировать списки растений, найденных на разных материках, как нельзя выдавать частную флору за олицетворение всего, что было в разных местах Земли. Надо изучать становление мезозойской флоры по конкретным областям, прослеживая истоки каждой области в палеозое. Надо помнить о том, что эволюция растений протекала в экосистемах, судьба которых контролировалась различными факторами, в том числе и небиологическими. Геологические перестройки открывали и перекрывали миграционные тракты. Морские бассейны и горные цепи имели для эволюции растений такое же значение, как государственные границы для истории народов. Открытые границы ведут к усилению связей, интеграции экосистем. Закрытые границы порождают обособление. Эволюция по разные стороны от границы протекает более автономно. Нельзя забывать и об изменениях климата.
О расшифровке корней мезозойской флоры в палеозое не может быть речи, если нет надежной систематики растений. Случаев, когда палеоботаники попадались на внешнем сходстве неродственных растений, не перечесть. Примеры, когда обманчивое сходство растений приводило к неверным фитогеографическим выводам, уже приводились. Не меньше недоразумений происходило и при анализе исторической преемственности флор.
В перми Гондваны известны членистостебельные, листья которых срослись по несколько штук в ланцетные лопасти. Похожие растения нашли в триасе Европы и сделали вывод, что европейская триасовая флора генетически связана с гондванской позднепалеозойской. Может быть, это и так, но органов размножения гондванских членистостебельных с такими листьями мы до сих пор не знаем. Без этого рискованно заключать и об исторической преемственности. В гондванском палеозое есть род Buriadia. Его ветки совсем как у хвойных, и их сначала отнесли к европейскому триасовому роду Voltzia, а когда отнесли к самостоятельному роду, то все равно оставили в семействе вольциевых. Вот якобы и еще одно подтверждение родства обеих флор. Теперь мы знаем, что органы размножения буриадий и вольций совершенно различны, а сходство их побегов мало что значит для выяснения флористических связей. Вывод очевиден: для сравнения флор, для суждений об их происхождении нужно детальное изучение растений, надо получить достаточно полные реконструкции растений, знать их анатомию, органы размножения.
К сожалению, выполнить все эти требования слишком непросто. Поэтому убедительные данные о происхождении мезозойской флоры накапливаются медленнее, чем хотелось бы. Мы вынуждены довольствоваться отдельными штрихами сложной картины, причем на каждый штрих уходят годы работы. Сужу об этом по собственному опыту.
С проблемой происхождения мезозойской флоры я столкнулся в общем-то случайно, еще занимаясь систематикой сибирских кордаитов. Мой университетский однокашник В. А. Селиверстов принес мне коробочку с несколькими отпечатками, найденными в верхнепермских отложениях в Вязовском овраге недалеко от Оренбурга. Отпечатки были покрыты угольной корочкой, из которой, судя по всему, можно было приготовить препараты кутикулы и изучить клетки кожицы. Похожие отпечатки тоже из Южного Приуралья незадолго до этого прислали М, Ф. Нейбург саратовские геологи. Тогда я составил опись образцов. Нейбург дала заключение о возрасте, и отпечатки предполагалось выбросить. На них почти не было видно жилок, а поэтому точное определение казалось невозможным. Почему-то образцы саратовцев остались лежать в посылочном ящике, и я вспомнил о них, только познакомившись с селизерстовскими отпечатками. На них тоже жилки почти не были видны, хотя сохранность угольной корочки была хорошей.
То, что при хорошей сохранности угольной корочки жилки плохо пропечатались на породе, показалось мне не случайным. Когда-то Нейбург поручала мне рисовать отпечатки триасовых птеридоспермов, у которых было то же самое. Я проклинал все на свете, пытаясь разглядеть направление жилок при освещении — косым светом то с одной стороны, то с другой. Не такой уж вроде бы существенный признак, хорошо ли пропечатались жилки. Скорее его надо относить за счет условий сохранности, а не своеобразия растений. Возможно, что я не придал бы этому признаку значения, если бы не сделал препаратов кутикулы с селиверстовских образцов. Получились препараты хорошо, можно было разглядеть на кутикуле каждую клетку, каждое устьице. Устьица оказались такими же, как у упомянутых триасовых птеридоспермов. Возникло подозрение, что это растения одной группы.
Коллекции от геологов, работавших в Южном Приуралье и в других районах европейской части СССР, продолжали поступать. В моем распоряжении были и коллекции, ранее полученные Нейбург от геологов. Собирался порядочный материал, с каждым годом все более интересный. Главный его смысл был в том, что он характеризовал флору Западной Ангариды, т. е. ее части, расположенной западнее Урала, и мог быть надежно привязан к стандартной стратиграфической шкале ярусов Перми. Это было важно для правильной датировки других пермских флор Ангариды, в том числе сибирских, чей возраст служил источником нескончаемых дискуссий.
К этому времени возня с кордаитовыми листьями, которым я уже посвятил почти 10 лет, успела изрядно надоесть. Появилась возможность заняться флорой, стратиграфически очень важной, но изученной еще очень плохо. Правда, первые образцы с отпечатками растений из перми Приуралья изучал еще А. Броньяр и немногие другие европейские палеоботаники. Часть из этих образцов привез из России известный английский геолог Р. И. Мурчисон, проехавший до Урала и установивший в результате путешествия пермскую систему. Он и назвал ее по тогдашней Пермской губернии. Потом отдельные коллекции описывал И. Ф. Шмальгаузен, а уже в нашем веке — М. Д. Залесский. Последний опубликовал в 1927 г. прекрасный атлас «Палеозойская флора уральских пределов Ангариды», где изобразил лучшие из собранных до него образцов, в том числе и из частных коллекций. Некоторые из них были собраны в Приуралье при разработке медных руд (медистых песчаников). Впоследствии Залесский вместе со своей сотрудницей Е. Ф.Чирковой объехал многие местонахождения и описал часть коллекций.
Нельзя сказать, чтобы западноангарская флора была совсем плохо изучена, когда я собрался ею заняться. Число описанных видов было довольно большим — много десятков. Воспользоваться же этими описаниями было почти невозможно: они были короткими, иллюстрации — скудными, а главное, не изучались детали жилкования, строение кутикулы, органы размножения растений. Более того, наиболее важные образцы оказались давно потерянными (в частности, почти вся коллекция Залесского погибла во время войны), а на оставшихся образцах то не было угольной корочки, то отпечатки плохо сохранились.
Коллекции, переданные геологами и хранящиеся в моих шкафах, были достаточно обширны, но и они не внушали полного доверия. Часто геологи собирают не те отпечатки, которые наиболее интересны, а то, что попалось на глаза и привлекло внимание. Когда я смотрел одну коллекцию, привезенную из окрестностей Кунгура, то обратил внимание, что в ней нет листьев кордаитов. Решив, что в этом кроется интересная закономерность, я поделился своими мыслями с человеком, собравшим образцы. «Да что вы! — услышал я в ответ. — Там было много кордаитов, но я не брал их! Целых листьев не нашел, а обрывки такие скучные!» Короче говоря, надо было поехать по главным местонахождениям самому.
В целом поездки были удачными, удалось разыскать многие классические местонахождения растений. В некоторых случаях в розысках помогли старожилы. Один из них прекрасно помнил приезд М. Д. Залесского и повел меня туда, где собирал растения «профессор из центра». Довелось побывать и там, где вел раскопки В. П. Амалицкий в поисках гондванских позвоночных и где он нашел листья, принятые им за глоссоптериевые. На Малой Северной Двине, там, где раньше неподалеку стояла деревня Соколки, сохранился гигантский раскоп, где поместился бы большой многоквартирный дом. Жители окрестных деревень до сих пор помнят рассказы отцов и дедов, на которых произвели впечатление и найденные скелеты «ящеров», и щедрая оплата нелегкого труда, и бесплатно раздававшаяся рабочим мазь от комаров. К сожалению, найти в этом раскопе ископаемые растения нам не удалось (правда, в нем выросла отменная современная смородина).
Коллекции собрались огромные — многие тысячи отпечатков из десятков местонахождений. Только усевшись за эти коллекции, я понял, какая трудная работа предстоит. Изучение материала затянулось и продлится еще не один год. Как ни приятно иметь дело с материалом хорошей сохранности, но корпеть над ним надо куда дольше, чем над отпечатком, на котором ничего не видно, кроме оттисков жилок. С иным образцом приходится заниматься неделями. Поэтому сейчас рано делать определенные выводы. Зато можно более осмысленно сформулировать некоторые проблемы, в том числе и проблему происхождения мезозойской флоры.
То, что изучение западноангарской флоры поможет разобраться в происхождении мезозойской флоры, стало понятно довольно быстро. Из Вязовского оврага, откуда были образцы В. А. Селиверстова, в первую же поездку в 1965 г. я привез много образцов. Помимо все тех же листьев с неясным жилкованием и хорошей угольной корочкой удалось найти и интересные органы размножения. Внешне они похожи на небольшой гриб: тонкая ножка, а на конце шляпка. Край шляпки фестончатый, а поверхность с радиально расходящимися от центра ребрами. Из угольной корочки, составляющей шляпку, получился неплохой препарат кутикулы. Под микроскопом можно было хорошо видеть устьица, такие же как и на селиверстовских листьях. В Вязовке нашлись и листья, угольная корочка которых была тонкой и на свет можно было видеть черные тяжи жилок. Они шли плотным пучком вдоль оси листа, а затем расходились к краям. На иных экземплярах можно было рассмотреть оттиски жилок, отпечатавшихся на породе. Там были какие-то дополнительные складки, их можно было принять за соединения между жилками.
Возникло предположение, что именно эти листья В. П. Амалицкий, а затем М. Д. Залесский приняли за глоссоптериевые. Я вспомнил, что в Палеонтологическом музее АН СССР им, Ю. А. Орлова, где хранятся найденные Амалицким скелеты, есть и привезенные им отпечатки растений. Предположение в общем подтвердилось. Отпечатки были действительно очень похожи, но на образцах Амалицкого не было угольной корочки, чтобы окончательно отождествить листья с Малой Северной Двины и Вязовского оврага. Во избежание недоразумений пришлось обозначать вязовские листья другим родовым названием. В качестве такового я выбрал Tatarina — по татарскому ярусу, в отложениях которого листья были найдены.
Палеоботаники часто вынуждены использовать разные родовые названия для листьев, у которых известно или нет строение кутикулы. Нередко листья, неотличимые по форме и жилкованию, принадлежат разным систематическим группам (например, цикадовым и беннеттитам), и установить это можно только по клеточному строению кожицы, отпечатавшемуся на кутикуле. Листья с неизвестным строением кожицы могут принадлежать любой из групп, но как-то надо их называть. Для таких листьев вводят особый род, который не относят ни к одной из групп. Поскольку отпечатки, собранные Амалицким и описанные Залесским сначала как Glossopteris, а затем как Pursongia, могли иметь разную кутикулу, в том числе и отличающуюся от той, что я видел у вязовских листьев, было рискованно называть последние Pursongia. Это название можно было оставить для всех отпечатков сходного облика и лишенных остатков кутикулы.
Я уже говорил, что по строению кутикулы татарины Вязовки оказались похожими на триасовые птеридрспермы. Более важным было то, что эти триасовые птеридоспермы — семейство пельтаспермовых — имели такие же, как в Вязовке, грибообразные органы размножения. Их еще в 30-х годах детально описал известный английский палеоботаник Т. М. Гаррис. Под шляпкой «гриба» прикреплялись семена, по опадении которых оставались рубцы. В этом месте кутикула прерывалась отверстиями. Уже потом я обнаружил оттиски округлых рубцов от опавших семян и у органов размножения татарин.
Большое количество этих растений в те же годы привезли из скважин Севера европейской части СССР геологи А. Г. Олферьев и В. Р. Лозовский. Местами порода была превращена в сланец из-за огромного скопления листьев татарин. Тут же встречались и «шляпки» (рис. 19). Количество татарин в отложениях татарского яруса (последнего яруса перми) было примерно таким же, как кордаитов в перми Сибири. Ясно, что это была главная группа растений в конце перми по всей Западной Ангариде. В те же годы И. А. Добрускина занималась триасовыми птеридоспермами и пришла к выводу, что пельтаспермовые гораздо более характерны для триасовых флор, чем можно было полагать раньше. Таким образом установилась первая достоверная прямая связь между пермской и триасовой флорами, подтвержденная и изучением микроструктуры листьев, и знанием женских органов размножения.
Одновременно стала ясна ошибка и Амалицкого, и Залесского, и последующих палеоботаников, связывавших пурсонгии (и, стало быть, татарины) с гондванскими глоссоптериевыми.
 Рис. 19. Облиственный побег верхнепермской татарины (Tatarina) и связываемые с такими растениями женские органы размножения (Peltaspermum) слева и пыльца (Vittatina) в кружке; длина линейки 1 см
Рис. 19. Облиственный побег верхнепермской татарины (Tatarina) и связываемые с такими растениями женские органы размножения (Peltaspermum) слева и пыльца (Vittatina) в кружке; длина линейки 1 см
Связь пермской флоры Западной Ангариды потянулась не в палеозой Гондваны, а в мезозой северных материков, поскольку в триасе пельтаспермовые особенно характерны для Европы, Северной и Средней Азии, Китая. Их остатки находили и в Гондване, но тоже только в триасе. Так что можно полагать, что они проникли сюда с севера. Древнейшие достоверные остатки пельтаспермовых птеридоспермов сейчас известны в кунгурском ярусе (последнем ярусе нижней перми) Среднего Приуралья. Круг предшественников мезозойской флоры, обнаруженных во флоре Западной Ангариды, не ограничивается пельтаспермовыми. Другие растения были известны и И. Ф. Шмальгаузену, и М. Д. Залесскому, но они почему-то не разглядели в них предтеч мезозоя. Впрочем, Шмальгаузен вообще мало занимался этой флорой, да и коллекции у него были небольшими. Что же касается Залесского, то он, по-видимому, был целиком поглощен идеей о близком родстве ангарской и гондванской флор. Кроме того, расшифровка общности западноангарских и мезозойских растений была невозможна без достаточно детальных исследований. Внешне же языковидные листья пурсонгий-татарин гораздо больше похожи на глоссоптериевые, чем на сильно рассеченные папоротникообразные листья триасовых пельтаспермовых. Примерно то же случилось и с другими растениями, которые, как я склонен считать, дали начало мезозойским родам.
В западноангарской флоре эти растения можно проследить до отложений кунгурского яруса. Глубже их корни теряются. В отложениях предшествующего, артинского яруса остатков растений уже не так много, а еще ниже по разрезу историю западноангарской флоры можно проследить только по палинологическим данным, т. е. по остаткам спор и пыльцы. Пока, к сожалению, это только принципиальная возможность, которую трудно реализовать. Мы уже знаем состав спор и пыльцы в докунгурских отложениях Западной Ангариды, но не знаем, каким растениям принадлежали те или иные формы спор и пыльцы.
Некоторые палинологи пытались решить эту задачу, сравнивая пермскую пыльцу с пыльцой современных растений. Скажем, у многих современных хвойных пыльца снабжена воздушными мешками, чаще всего двумя. Такую же пыльцу находят и в пермских отложениях, так и называя ее пыльцой хвойных. Такой путь более чем рискованный, поскольку палеоботаникам известно несколько групп растений (помимо хвойных), обладавших пыльцой с двумя мешками. Это различные птеридоспермы и растения с глоссоптериевой листвой.
Первоначально я надеялся
проследить с помощью палинологии хотя бы историю пельтаспермовых птеридоспермов. Их пыльца была изучена у триасовых представителей, и можно было надеяться, что и в перми она была того же облика — в виде французской булки. Я попросил палинологов обработать несколько образцов, в которых было особенно много листьев татарин. Мне принесли пробирки с выделенной из породы пыльцой. Тщетно я просматривал под микроскопом препарат за препаратом. Пыльцы в них было очень много, и самой разнообразной, но ожидавшейся не было. Зато было особенно много пыльцы с многочисленными ребрами (род Vittatina) и мешковой с ребристым телом. Оставалось предполагать или то, что пыльца моих пельтаспермовых не сохранялась в ископаемом состоянии (такие растения с крайне неустойчивой к захоронению пыльцой известны), или что она была другого облика, чем в триасе. Мысль, что татарины принадлежат не пельтаспермовым, а другим растениям с иной пыльцой, я не допускал, поскольку знал их женские органы размножения и строение кутикулы листьев. Вскоре А. В. Гоманьков, с которым я разделил изучение западноангарской флоры и который занялся пельтаспермовыми, обнаружил, что иногда под «шляпками» их органов размножения огромными массами скапливается пыльца типа Vittatina. Трудно поверить, что так поведет себя пыльца чужого растения. Почти одновременно как будто решился вопрос и с мешковой пыльцой, имеющей ребристое тело. Я копался с непонятным обугленным остатком, найденным вместе с татаринами и похожим на какую-то сильно деформированную, сплющенную шишку. Разглядеть в обугленной лепешке ничего не удавалось, и я решил обработать химически небольшой кусочек, чтобы понять, что это такое. После обработки окислителем и щелочью с кусочком произошло нечто странное. Он превратился в студенистую белесую массу, буквально набитую однородной пыльцой с двумя мешками и отчетливой ребристостью тела. Помимо массы пыльцы нашлись мелкие обрывки листовой кутикулы, такой же, как на листьях татарин. Дальше можно было не ломать голову над строением обугленной лепешки: это был копролит (окаменелый навоз) какого-то животного, объедавшего мужские органы размножения татарин и прихватывавшего листья (фиг. X).
Кто бы мог подумать, что животное, пасшееся в татариновых зарослях и оставившее аккуратную кучку, тем самым окажет услугу палеоботанике? Жаль только, что мало было этих животных и поэтому в наших коллекциях после тщательного их пересмотра нашлось лишь два других копролита, изучение которых еще впереди. Опущу дальнейшие перипетии исследования и сразу сделаю вывод: мы с А. В. Гоманьковым сейчас склонны считать, что растения с татариновыми листьями имели оба типа ребристой пыльцы, т. е. с мешками и без мешков, а также неребристую двухмешковую пыльцу.
Пыльца этих типов известна по всему разрезу перми и в небольшом количестве встречается в верхах карбона. Впрочем, из этого мы не заключаем, что пельтаспермовые появились в карбоне. Известно, что такую же пыльцу имели и другие растения, в том числе и некоторые гондванские глоссоптериевые. Там находили ребристую пыльцу с мешками, проникшую в семезачатки семян или еще не выпавшую из мужских спорангиев. Хитро все переплелось. Находим похожие побеги в разных местах, а потом оказываются разными органы размножения. Потом находим похожую пыльцу, и снова оказывается, что под одинаковой внешностью кроются разные растения. В который уже раз получают палеоботаники уроки осторожности.
В недостатке осторожности повинны все палеоботаники. Мне тоже случалось сильно обжигаться и попадаться на обманы внешнего сходства. Поэтому я не вправе первым бросать в кого-то камень. Если я сейчас выскажу упрек некоторым палинологам, то не потому, что сам безгрешен, а только потому, что этот упрек уже высказывали многие до меня,
Я имею в виду многочисленные указания в палинологической литературе на находки в палеозойских отложениях родов, характерных для мезозоя и даже кайнозоя. Ту самую ребристую безмешковую пыльцу, которую сейчас можно довольно уверенно связать с пельтаспермовыми, палинологи сравнивали с пыльцой современной группы покровосеменных, почти не известной широкой публике (знают только принадлежащую покровосеменным лекарственную эфедру). О хвойных я уже говорил. В перми Сибири находят пыльцу на первый взгляд того же типа, что и у мезозойских цикадовых, гинкговых и беннеттитовых. Соответственно в палинологических статьях порой пишут, что в перми Сибири жили эти группы растений, предшественники мезозоя. При этом делают сразу несколько ошибок. Во-первых, такую же пыльцу, как и у перечисленных групп, имели и пельтаспермовые птеридоспермы, которые тоже просятся в компанию. Во-вторых, пермская пыльца Сибири на самом деле совсем иная, чем у этих мезозойских групп.
Было показано, что за безмешковую пыльцу палинологи часто принимают оторвавшиеся и сложившиеся в лодочку мешки одномешковой пыльцы растений с листвой рода Rutloria. В такие же лодочки сворачиваются и оставшиеся без мешков тела. Наконец, та же пыльца, еще не созревшая, тоже складывается в лодочку. Три типа сохранности одной и той же пыльцы описываются как нечто совсем разное, да еще относятся к группам, характерным для мезозоя. В результате у мезозойской флоры отрастают ложные корни.
До сих пор речь шла только о Западной Ангариде как об источнике мезозойской флоры. Читатель может подумать, что это был какой-то сравнительно небольшой (в масштабе планеты) рассадник растений, которые при подходящих условиях расселились в мезозое и заполнили всю Землю. Едва ли дело происходило так, В литературе не раз выдвигались подобные точки зрения на происхождение и крупных систематических групп растений (например, покрытосеменных, о которых пойдет речь в следующей главе), и целых флор. В качестве главного источника флоры мезозоя выдвигались сибирская пермская флора и западноевропейская цехштейновая флора. Думаю, что на самом деле все было иначе. Каждая из флористических областей перми сделала больший или меньший вклад в растительный покров мезозоя. Кроме того, мезозойские группы растений, прежде чем осесть на своих местах, могли совершить далекие путешествия, основывая вторичные центры расселения.
Для реконструкции этого сложного процесса надо хорошо знать филогению основных мезозойских групп растений. Тут главный пробел в наших знаниях. В типично мезозойских флорах, населявших Землю от конца триаса примерно до середины мела, главную роль играют гинкговые, лептостробовые, или чекановскиевые (о них будет рассказано в следующей главе), цикадовые, беннеттиты, различные хвойные и папоротники, местами — птеридоспермы. Происхождение всех этих групп остается полностью неизвестным. Предполагают, что цикадовые произошли от каких-то палеозойских птеридоспермов, а гинкговые от кордаитов. Мезозойские хвойные филогенетически связывают с палеозойскими. Все эти и другие гипотезы пока подкрепляются не конкретными находками переходных форм, а некими абстрактными схемами, показывающими, как могли эволюционировать отдельные органы.
Может быть, в будущем мы будем полнее знать филогению основных мезозойских групп, а пока к проблеме происхождения мезозойской флоры приходится подходить иначе. Мы можем проанализировать, в каких пермских флорах больше всего представителей тех групп, которые особенно характерны для мезозоя, и каким образом соотносятся палеофлористические области перми и мезозоя.
При таком подходе картина получается следующей. Начнем с Ангариды. Типично ангарские пермские флоры, известные в Сибири и составленные преимущественно кордаитами и членистостебельными, едва ли сделали серьезный вклад в мезозойскую флору. Правда, здесь встречаются отдельные растения, которые сравнивают с мезозойскими родами, но в других местах Земли соответствующих растений гораздо больше. Иное дело — Западная Ангарида, о которой уже много говорилось. Эта территория лежала тогда в низких широтах, на краю тропической зоны, причем климат был примерно таким же, как в современном Средиземноморье, или более теплым. Судя по почти полному отсутствию углеобразования, в этих местах было довольно сухо. Неподалеку стояли Прауральские горы. Считается, что предгорные области с теплым и не слишком влажным климатом, большим разнообразием экологических условий особенно благоприятны для быстрой эволюции растений. Западная Ангарида хорошо отвечает перечисленным требованиям.
В последние годы накапливается все больше данных, что того же типа флора не только заселяла Приуралье и соседние с ним части Восточной Европы, но и уходила в Казахстан, Среднюю Азию и далее на восток, в Китай. Видимо, Ангарида была окружена поясом (назовем его Субангарской областью; рис. 20), в котором аборигены смешивались с пришельцами с севера и юга. На опубликованных в палеоботанической литературе палеофлористических картах эта область пока отсутствует, а очерчена лишь ее западная часть, получившая название Восточно-Европейской области (в поздней перми) и Уральско-Казахстанской области (во второй половине ранней перми).
 Рис. 20. Фитогеография середины перми (куньгурский, уфимский и казанский века): 1 — Ангарская область; 2 — Субангарская область (кружками показана и примесь субангарских растений в других флорах); 3 — Катазиатское царство; 4 — Атлантическое царство (включая цехштейновую флору); 5 — Северо-Американское царство
Рис. 20. Фитогеография середины перми (куньгурский, уфимский и казанский века): 1 — Ангарская область; 2 — Субангарская область (кружками показана и примесь субангарских растений в других флорах); 3 — Катазиатское царство; 4 — Атлантическое царство (включая цехштейновую флору); 5 — Северо-Американское царство
Гипотеза о существовании Субангарской области складывалась у меня постепенно из разрозненных наблюдений. В 1971 г. я был в Алма-Ате и смотрел коллекции палеоботаника К. 3. Сальменовой, собранные в Прибалхашье и Джунгарии. Хотя сохранность растительных остатков была довольно плохой (на растительных остатках нет угольной корочки, и поэтому нельзя приготовить препараты кутикулы), некоторые растения разительно напоминали приуральские. Возможность связи юго-востока Казахстана с Приуральем подтверждалась и некоторыми палинологическими данными по пермским отложениям района Джезказгана и Южного Казахстана (Чуйской впадины и Малого Каратау). Спорово-пыльцовые комплексы этих мест сходны с приуральскими. Затем ташкентский геолог А. С. Масумов прислал мне коллекцию, собранную в Чаткальском хребте. Снова обнаружились растения, до этого считавшиеся характерными для Среднего Приуралья. Потом К. 3. Сальменова показала мне прибалхашские отпечатки рода Phylladoderma, типичного для европейской части СССР. Этот же род В. И. Бураго нашла в верхней перми Приморья.
Неожиданной была коллекция из Афганистана, собранная душанбинским геологом В. И. Дроновым вблизи Кабула (фиг. XI). Здесь смешивались растения, характерные для верхней перми Поволжья и Прикамья, и явные пришельцы из Китая. Последнее меня не удивило, поскольку комплекс катазиатских растений был еще в 50-х годах найден английским палеоботаником Р. Вагнером в верхней перми на востоке Турции. Позже чехословацкий геолог П. Чтироки прислал мне фотографии растений, собранных им в Ираке и ошибочно отнесенных к гондванским родам и видам. Это тоже были катазиатские растения. Наконец, надо упомянуть коллекцию, собранную еще в начале 30-х годов шведским геологом Г. Бекселлом в хребте Наньшань восточнее коленообразной излучины Хуанхэ. Здесь выше отложений с катазиатской флорой были собраны растения, отнесенные шведским палеоботаником Т. Г. Галле к видам, характерным для перми Сибири. Из этого был сделан вывод, что вся сибирская ангарская флора моложе катазиатской. К коллекции Бекселла, оставшейся неописанной, никто не возвращался более 40 лет. Потом в Москве побывали шведские ученые Э. Норин и Б. Болин, которые пообещали прислать фотографии образцов Бекселла. Определения Галле оказались ошибочными, но дать новые по фотографиям было рискованно. Надо было изучать сами образцы. В 1977 г. М. В. Дуранте, до этого исследовавшая верхнепалеозойскую флору Монголии, поехала в Швецию. Ее работа продолжилась в Москве, куда была прислана часть образцов. Исследования коллекции Бекселла еще продолжаются, но уже можно сделать вывод, что и здесь есть немало растений, которые мы привыкли считать западно-ангарскими.
Точно оконтурить Субангарскую область пока невозможно. Трудно расшифровать ее соотношение с соседями как с севера, так и с юга. Пермская история районов с субангарской флорой крайне запутана. Ситуация осложнилась и тем, что в это время, примерно в начале поздней перми, катазиатские растения нашли дорогу на запад Евразии. Другие растения шли им навстречу (например, в верхней перми Дарваза найдены некоторые цехштейновые растения). С севера приходили сибирские растения, а с юга, видимо, гондванские. В верхнепермских отложениях Сибири и Приморья попадаются листья, очень похожие на глоссоптериевые. В некоторых местах получались удивительные смешанные флоры. Так, в палинологических комплексах верхней перми Соляного кряжа австралийский палинолог Б. Балм нашел странную смесь западноевропейских, приуральских и гондванских родов. Еще предстоит разобраться с соотношением субангарской и североамериканской флор. Судя опять же по палинологическим данным, между ними было немало общего.
Субангарская флора — неожиданно обнаружившееся белое пятно на палеофлористических картах перми. Его закрытие — одна из главных задач палеофлористики палеозоя на ближайшие годы. Не требуется пояснять, что одновременно удастся многое понять и в происхождении мезозойской флоры.
Южнее субангарской флоры располагались Катазиатская область (на востоке Евразии) и Атлантическая область (с флорой цехштейна). Обе они могли сделать некоторый вклад в мезозойскую флору. Для первой характерно большое количество растений с листьями, похожими на цикадовые. Может быть, здесь был центр происхождения цикадовых. В цехштейне много хвойных, и, хотя ни один род не перешел в триас, можно наметить филогенетические связи между цехштейновыми и триасовыми хвойными. Много хвойных и в Северной Америке.
Для понимания происхождения мезозойской флоры много могут дать детальное изучение и точная датировка растений Сибири, которые пришли здесь на смену кордаитам и пермским членистостебельным. Впервые эти растения были собраны в Кузбассе М. Ф. Нейбург и в Тунгусском бассейне Л. М. Шороховым еще в 30-х годах. Кузнецкие растения сразу отнесли к триасу, а дискуссия о возрасте тунгусских растений затянулась. Сначала их относили то к перми, то к триасу, а затем единодушно остановились на триасовом возрасте. Когда в 50-х годах началось детальное изучение отложений с тунгусскими растениями (из так называемой корвунчанской серии), выяснилось, что остатки животных (ракообразных, двустворчатых моллюсков) и спорово-пыльцевые комплексы в нижней части этих отложений скорее пермские, чем триасовые. Палеоботаник и стратиграф Г. Н. Садовников пришел к выводу, что корвунчанская серия состоит из двух частей. Нижняя относится к самым верхам перми, а верхняя — к триасу. Это решение кажется мне вполне разумным, хотя и нуждающимся в дополнительном обосновании. Многие палеоботаники придерживаются иного мнения. Они считают, что корвунчанская серия целиком принадлежит триасу. В подтверждение указывают на мезозойский облик корвунчанской флоры (фиг. XII), на резкую смену состава как самих отложений, так и растительных остатков на нижней границе корвунчанской серии.
Все это так. Действительно, вулканогенная корвунчанская серия резчайшим образом отличается от подстилающих угленосных отложений. Я согласен и с тем, что флористическая граница на этом рубеже тоже чрезвычайно резкая. Однако из этого никак не следует, что перестройки и в режиме осадконакопления, и в растительном покрове должны были произойти именно на рубеже перми и триаса. Они могли произойти и раньше, и позже. Мы должны сначала сопоставить разрезы Тунгусского бассейна с эталонными разрезами верхов перми и низов триаса, а уж потом решать, когда, где и что случилось.
Сопоставить разрезы можно по остаткам животных и растений. Животные довольно единодушно высказались в пользу пермского возраста нижней части корвунчанской серии. С растениями получилось сложнее. Правильно, что в корвунчанской серии есть мезозойские роды. Но будем точными. Эти роды хотя и мезозойские, но никогда не считались характерными для нижнего триаса. Они обычно считались верхнетриасовыми и даже юрскими. Корвунчанскую серию никто не относит к верхнему триасу или юре. Стало быть, все равно допускается, что некоторые роды спускаются в Сибири ниже по разрезу, чем в других местах Земли. Если же разрешается им спуститься до нижнего триаса, то почему надо закрывать им дорогу в пермь? При этом в эталонных разрезах верхов перми, т. е. в Западной Ангариде, облик флоры уже мезозойский. Здесь господствуют пельтаспермовые птеридоспермы — листья татарин, о которых много говорилось. Кстати, и в нижней части корвунчанской серии есть прослои с большим количеством татарин. В Западной Ангариде кордаиты полностью исчезают не на рубеже перми и триаса, а раньше. Поэтому связывать исчезновение кордаитов в Сибири с границей перми и триаса пока нет оснований.
Если возраст корвунчанской флоры действительно частично: пермский, то и эта флора может быть включена в число предшественников мезозоя. Впрочем, возможно, что Сибирь была не первичным, а вторичным центром расселения мезозойских растений, как бы временным лагерем в середине маршрута. На это предположение наводят, во-первых, все те же пельтаспермовые, которые появились в Субангарской области намного раньше, чем в Сибири, а во-вторых, почти полное отсутствие преемственности корвунчанской флоры и ее предшественницы в самой Сибири.
Я вообще склонен думать, что смена палеозойских флор на мезозойские происходила на фоне расширявшейся миграции растений, ранее сидевших по своим областям. Еще в середине перми началось оживленное движение по периферии Ангариды вдоль Субангарской области. В начале поздней перми тронулись в путь катазиатские растения, из Китая покатились на запад, добрались до Турции, Ирака и, видимо, пошли еще дальше. В самом конце перми (если раннекорвунчанская флора еще относится к перми) началось заселение Сибири новой флорой, пришедшей на смену кордаитам.
В триасе этот процесс продолжался. В раннем триасе чуть ли не по всему миру распространились своеобразные плауновидные плевромейи, сначала описанные в Европе, а потом в окрестностях Рыбинска, на Мангышлаке, в Приморье, на северо-востоке СССР. Недавно Г. Н. Садовников нашел огромные скопления их остатков на восточном Таймыре. Близкие растения найдены в Средней Сибири, Индии и Австралии. Где они появились, мы не знаем. Возможно, что их родиной была опять же Западная Ангарида, где в верхней перми встречаются похожие плауновидные.
Необычайно широкое распространение плевромей лучше, чем что-либо иное, подчеркивает процесс разложения, распада флористических областей, сложившихся в позднем палеозое. Некоторые фитогеографические рубежи продолжали существовать и позлее, но теперь они не охранялись так строго, как раньше. Трудно сказать, привело ли исчезновение преград между областями к ломке местных флор из-за многочисленных пришельцев, или общее изменение палеогеографической обстановки привело как к ломке преград, так и к изменению флор. Где здесь причины, где следствия, мы пока не знаем, да и сама эпоха смены палеозойских флор на мезозойские изучена еще очень плохо. Здесь обширное поле деятельности не только для палеоботаников, но и для стратиграфов. Сопоставление разрезов пограничных отложений между пермью и триасом еще остается весьма дискуссионным. Без этого трудно расшифровать все случившееся на рубеже палеозоя и мезозоя,
В заключение остановлюсь на одном методическом вопросе. Граница перми и триаса давно привлекает внимание палеонтологов. Связанная с ней смена органического мира долго считалась внезапной и повсеместной. Найти земное объяснение столь крупным переменам казалось невозможным, и на ум стали приходить космические причины, например взрыв по соседству с Солнечной системой какой-то сверхновой звезды. Ее жесткая радиация, дескать, уничтожила палеозойский органический мир и пощадила лишь некоторых счастливцев, потомки которых и заселили мезозойские моря и континенты.
Я не собираюсь оспаривать эту и подобные гипотезы. Просто они кажутся мне неинтересными. Неизвестно, что надо делать, чтобы их опровергать или доказывать. Никто не знает, взрывались ли сверхновые в окрестностях Солнечной системы, а если да, то как они могли влиять на земную жизнь. Критики гипотезы о взрыве сверхновой уже указывали, что кратковременная радиация не могла породить многоступенчатую смену органического мира на границе перми и триаса, где разные группы животных и растений меняются в сложной и пока плохо расшифрованной очередности. И не только в этом дело. Думается, что к помощи сверхновой звезды надо обращаться не тогда, когда мы лишь начинаем разбираться с последовательностью событий, а позже — когда последовательность восстановлена, положена на хороший палеогеографический фон, когда мы восстановили смену экосистем и увидели, что все происходило вопреки привычным земным причинам. Последним убежищем и будет взрыв сверхновой. Сейчас же это то, что наши мудрые предки назвали asylum ignorantiae — убежищем невежества, к которому, как сказано в Логическом словаре Н. И. Кондакова, «прибегают, поскольку ленятся или не хотят глубже исследовать спорный вопрос».
Путей для исследований отнюдь не мало, и, уж конечно, они не исчерпаны. Смену морских фаун на границе перми и триаса связывали с перестройкой экосистем, вызванной обширными регрессиями морей, уходом их с континентов и осушением шельфов (так считал, например, наш видный палеонтолог В. Е. Руженцев). В случае наземного органического мира можно тщательно изучить связь миграций флор и фаун с перестройкой физико-географических барьеров. Словом, надо еще много потрудиться, чтобы отчаяться в отыскании земных причин и удариться в абстрактные космические домыслы.
Глава VIII
«ОТВРАТИТЕЛЬНАЯ ТАЙНА»

Я не знаю, все ли области науки доступны для популяризации. Наверное, не все. Иногда мне кажется, что и популяризация палеоботаники — дело безнадежное. О чем бы ни повел палеоботаник речь, ему придется так или иначе употреблять множество слов, незнакомых широкой публике. Чем-то их заменить невозможно, а без них совершенно немыслимо рассказать о самом важном и действительно интересном. Короче говоря, чтобы читать популярную книгу по палеоботанике, надо освоить некий новый язык со словами «спорангий», «птеридоспермы», «карбон», «Гондвана». У кого хватит терпения все это держать в голове?
Все же я взялся за эту книгу и вот подбираюсь к ее концу. Меня ободряет то, что среди ее читателей будет достаточное количество географов, геологов и ботаников, а среди них и те, кто интересуется ископаемыми растениями, но не в силах следить за огромной палеоботанической литературой. Полагаю, что та глава геологической летописи, о которой сейчас пойдет речь, интересует ботаников больше всего. Я собираюсь говорить о происхождении и расселении покрытосеменных, или цветковых.
Для человека, неплохо знакомого с палеоботаническими документами, происхождение покрытосеменных не составляет какой-то экстраординарной проблемы. Да, мы не знаем их предков и не знаем, как они возникли. Что ж из того, что в этом особенного? Много ли крупных групп растений, происхождение которых известно? Если отвечать честно и не выдавать шаткие и валкие домыслы за хорошо обоснованные гипотезы, то таких групп не окажется вовсе. Что мы знаем о происхождении цикадовых, гинкговых, хвойных (и вообще голосеменных), папоротников, членистостебельных, мхов, разных групп водорослей? А почти ничего не знаем.
И все же ситуация с покрытосеменными несколько особая. Это самая крупная группа современных растений. О значении покрытосеменных для человечества нет нужды говорить. Покрытосеменные — это хлеб, овощи, фрукты, топливо, строительная древесина, цветы на окнах и в цветочных магазинах, лекарства в аптеках. Классификация покрытосеменных — не отвлеченная научная проблема. Разобраться же в предложенных классификациях и отобрать наиболее удачную и практически наиболее удобную без понимания эволюции всей группы почти невозможно.
Поразительно разнообразие покрытосеменных. Достаточно вспомнить ряску и дуб, пшеницу и кактус. Много ли общего между прекрасным цветком магнолии и скромным ржаным колосом? Откуда взялось это невероятное разнообразие, как понять его? Все это волнует ботаников уже несколько столетий. С тех пор как в ботанику и биологию вообще проник исторический метод исследования, возникла, да так и не исчезла проблема происхождения покрытосеменных. Быстрое появление покрытосеменных в недавние геологические эпохи еще Ч. Дарвин назвал «отвратительной тайной» — abominable mistery. Замечу, что прилагательное abominable в толковом словаре еще поясняется словами «противная» и «ненавистная».
Ситуация и в самом деле какая-то противная. Представим себе, что некто хвалится познаниями в радиотехнике. Его просят починить что-то совсем обычное — радиоприемник или телевизор, и оказывается, что в них-то он ничего не понимает. С историческим методом в ботанике получается примерно то же. Сказано немало высокопарных слов об огромном значении историзма в систематике и морфологии растений, но мы все еще бессильны расшифровать происхождение и эволюцию самых распространенных высших растений.
Впрочем, словам Дарвина уже сто лет (он их написал в письме ботанику Дж. Д. Гукеру в 1879 г.). Может быть, с тех пор все изменилось? Многие ботаники оптимистически скажут, что изменилось немало и что мы сейчас гораздо лучше представляем себе и предков, и эволюционные пути покрытосеменных. Кто-то, может быть, расскажет про новейшие гипотезы. Некоторым из них посвящены толстенные книги. Кто-то напомнит об огромной литературе из тысяч статей и монографий на эту тему. Если продуктивность научной работы исчислять количеством и объемом опубликованных работ, то прогресс действительно будет потрясающим. Однако потрясение от прогресса мгновенно улетучится, как только мы начнем знакомиться с выводами, к которым приходят исследователи. Разнобой взглядов потрясет читателя гораздо больше, чем любой из выводов.
Я не собираюсь утверждать, что мы так и остались на уровне незнания столетней давности. Наши знания с тех пор изрядно увеличились. Ботаники несравненно лучше знают теперь строение и функционирование покрытосеменных, их образ жизни и распределение по лику планеты. Выдвинуты некоторые весьма правдоподобные гипотезы о том, какие могли существовать филогенетические отношения между отдельными родами, семействами и порядками. Что же касается самого важного — исторического отношения крупных групп покрытосеменных, то здесь прогресс ничтожен. Активность исследователей в отношении всех важнейших вопросов больше напоминает не организованное шествие, а толчею в магазине. К какому-то прилавку вдруг выстраивается огромная очередь. Выстояв несколько часов за дефицитным товаром, люди с удивлением узнают, что стояли зря: товара нет и не будет, а продают какую-то чепуху. В начале 50-х годов были описаны органы размножения глоссоптерид из гондванской флоры Южной Африки. Прямо в основании листа на ножке сидят овальные органы, в которых, как показалось сначала, мужские спорангии сочетаются с семенами. Обоеполые цветки — характернейшая черта покрытосеменных. Органы размножения глоссоптерид — тоже обоеполые, что большая редкость среди голосеменных. Читатель, может быть, помнит, что глоссоптериевые листья имеют сетчатое жилкование, и в этом отношении они, стало быть, тоже похожи на листья покрытосеменных. Родилась надежда, что именно гондванские глоссоптериевые — то недостающее звено в филогении, которое связывает голосеменных с покрытосеменными.
Статья была опубликована в одном из наиболее авторитетных палеонтологических журналов. Главный редактор журнала понимал возможный резонанс сообщения. Поэтому рукопись статьи была показана нескольким видным палеоботаникам. Их комментарии были опубликованы вместе со статьей. Мнение было, почти единодушным: доказательств, что у глоссоптерид действительно были обоеполые органы размножения, не представлено. Материал, который описан и изображен, слишком плохой сохранности. Общая структура органов размножения непонятна. Особенно сомнительны те структуры, которые были приняты за мужские спорангии. Словом, критика была не просто резкой, а почти уничтожающей и… бесполезной. Доверчивых и не слишком внимательных читателей оказалось немало. Гипотетические интерпретации стали широко известны, а предостережения палеоботаников прошли незамеченными. В ботанической литературе появились статьи, где глоссоптериды прямо назывались предками покрытосеменных, а Гондвана — их родиной.
Поразительные вещи происходят порой в науке. Я готов всерьез отстаивать взгляд, что механизм возникновения и распространения многих научных убеждений не многим отличается от того, что происходит со сплетнями. Кто-то что-то видел или читал, толком не разобрался, заявил во всеуслышание. Ему сразу возразили, все сказанное убедительно опровергли, а присутствовавшие чего-то недослышали. Вот и поползли пустые и досадные разговоры.
 Рис. 21. Глоссоптериевый женский орган размножения, как его представила Э. Пламстед (наверху), по современной реконструкции Т. Делевориаса и Р. Гулда (внизу)
Рис. 21. Глоссоптериевый женский орган размножения, как его представила Э. Пламстед (наверху), по современной реконструкции Т. Делевориаса и Р. Гулда (внизу)
Я не хочу винить кого-то в публикации статьи. В общем была соблюдена осторожность и в выводах, и в частных интерпретациях образцов. Некоторые наблюдения были действительно интересными. Например, было очевидно, что найдены какие-то органы размножения, и они в самом деле прикреплялись к основанию листа. На некоторых образцах были хорошо видны семена. Ограничиться бы этими сообщениями, и все было бы хорошо. Впрочем, теперь поздно давать советы «задним числом».
За прошедшие с тех пор годы число находок глоссоптериевых органов размножения сильно увеличилось (рис. 21). Недавно нашли их полностью минерализованными, так что можно рассмотреть каждую клетку. От прежних интерпретаций давно ничего не осталось. Никаких обоеполых органов размножения у глоссоптериевых не было. Мы знаем, какие у них были семена и мужские спорангии и как они сидели. На сплетню, которая давно гуляет по свету, все это не повлияло.
В статье, вышедшей в 70-х годах, когда ошибка была давно ясна, читаем: «Glossopteridales, по мнению всех исследователей, хотя и не являются прямыми предками покрытосеменных или, во всяком случае, не всех покрытосеменных, но очень близки к ним». В этой цитате выражение «по мнению всех исследователей» подобно словам «все говорят» в каждой порядочной сплетне.
Проблема происхождения покрытосеменных знает немало слухов. В 50-х годах появилось сообщение, что в каменноугольных отложениях нашли пыльцу покрытосеменных. Это были любопытные оболочки почти квадратного очертания с ясными порами по углам. Сходство с пыльцой действительно было. Палинологи быстро выпустили воздух из раздувшейся сенсации. Оказалось, что эти оболочки принадлежат каким-то морским водорослям. И все же несколько лет назад один видный ботаник подошел ко мне во время перерыва совещания и спросил, какие есть новости о пыльце покрытосеменных в карбоне. Слух оказался живучим.
В 1955 г. М. Ф. Нейбург описала странное растение, найденное Е. С. Рассказовой в пермских отложениях Печорского бассейна (фиг. XIII, XIV). На оси сидят мясистые конусовидные органы, несущие на конце щетку булавовидных придатков. Между придатками Нейбург нашла еще не опавшее семя. Видимо, статья Пламстед, вышедшая незадолго до этого, повлияла на интерпретацию печорского растения (Нейбург назвала его «войновския»). Почему-то Нейбург решила, что булавовидные придатки — мужские спорангии, а значит, войновския — обоеполый орган размножения. И опять в литературе заговорили о том, что найден возможный предок покрытосеменных.
Я принимал на веру вывод Нейбург о строении войновскии, пока известный английский палеоботаник Т. М. Гаррис не стал меня с пристрастием допрашивать, какие есть доказательства, что булавовидные придатки действительно мужские спорангии. Я ничего не мог ответить. Просто так считала Нейбург. Все же пришлось войновскией заняться более обстоятельно. Оказалось, что Гаррис спрашивал не зря. В булавовидных придатках ничего общего со спорангиями не оказалось. Это были всего лишь видоизмененные чешуи, сидевшие между семенами. Такие чешуи известны у многих голосеменных.
Подобные совершенно явные недоразумения составляют значительную, но не львиную долю литературы по происхождению и расселению покрытосеменных. Больше всего в ней работ, где исследователи пытаются понять случившееся в далеком прошлом, глядя на современный растительный мир. Они изучают современные цветки и пытаются понять, как эволюционировали их части, как произошли плодолистики, тычинки, венчик. Наблюдая современные жизненные формы, они спорят о том, с деревьев или трав началась эволюция покрытосеменных. Обсуждается, какими были листья первых покрытосеменных — цельными, как у лавра, или расчлененными, как у винограда. По тому, как расселены ныне по поверхности Земли разные группы покрытосеменных, ботаники пытаются расшифровать их центры происхождения и пути расселения.
Палеоботанические материалы в этой литературе фигурируют лишь от случая к случаю. Пусть не сердятся на меня ботаники, если я скажу, что поверить в выводы, которые они делают, в большинстве случаев невозможно. Причина моего неверия очень простая. Как правило, выводы о филогении, сделанные по разным признакам, вступают в противоречие. Скажем, среди ботаников популярна идея, что травянистые формы происходят от древесных, что цветки с большим числом плодолистиков, тычинок и лепестков дали начало цветкам с малым числом частей, что пыльца с тремя бороздами примитивнее многобороздной и т. д. Если попытаться расположить в филогенетической последовательности конкретные роды или семейства, то окажется, что разные свидетели примитивности и прогрессивности дают несогласованные показания. Складывающиеся ряды форм можно «читать» филогенетически по-разному. Такого, чтобы показания всех признаков совпали, не случается почти никогда. (Слово «почти» я вставил из осторожности. Думаю, что без него можно и обойтись.)
В новейших сводках по филогении покрытосеменных самыми примитивными считают растения, приближающиеся по общему облику к современным магнолиевым. Порядок магнолиевых стоит у корня филогенетического древа, ветви которого составляют все прочие порядки числом около сотни. Покопавшись в литературе, можно найти и другие мнения. Некоторые ботаники считают, что из магнолиевых нельзя вывести однодольных, что крупные одиночные цветки, не собранные в соцветия, признак не примитивности, а прогрессивности. Указывают, что по биохимическим признакам магнолиевые — весьма высокоразвитая, а отнюдь не примитивная группа. Правда, другие биохимики с этим не соглашаются.
Что делать при таких разнотолкованиях? Как убедиться, кто прав и кто не прав? Ботаники ищут выхода из противоречий в расширении круга анализируемых признаков. К сожалению, увеличение числа признаков ведет лишь к умножению противоречий. Новые признаки не согласуются друг с другом и со старыми. Я не буду вдаваться в теоретические рассуждения, как быть в подобных случаях, как разрешать филогенетические противоречия. Кое-что на этот счет сказано в следующей главе. Сейчас лишь замечу, что решение филогенетических проблем без обращения к палеонтологической летописи, по-видимому, принципиально невозможно.
В ботанической литературе широко распространено мнение о чрезвычайной неполноте палеонтологической летописи. Поэтому, дескать, ископаемые растения — разрозненные и мало о чем говорящие документы. В одной из сводок по эволюции и классификации покрытосеменных так и написано, что геологическая летопись, польза которой «весьма ограничена для прояснения родственных отношений между разными группами покрытосеменных, не более полезна в выявлении их предка».
В тех палеоботанических примерах, с которыми мы уже познакомились в этой главе, роль палеоботаники действительно довольно неприглядна. Скорее она была источником путаницы, а не знания. К счастью, не все палеоботанические материалы того же достоинства. Палеоботаника сейчас располагает очень интересными фактами, которые, правда, не всегда известны ботаникам. Особенно важные сведения были получены в последние 10–15 лет. Почему-то скорость их распространения куда более скромна, чем скорость сообщений об обоеполых органах размножения глоссоптериевых и войновский.
Думаю, что причина этого прежде всего в том, что новые данные плохо согласуются с наиболее признанными убеждениями. Они заставляют пересмотреть всю систему взглядов на происхождение и расселение покрытосеменных, Я недаром выделил разрядкой слово «система». Проблема происхождения и расселения покрытосеменных — сложное событие. Для его реконструкции надо учитывать широкий круг фактов, доставляемых не только ботаникой и палеоботаникой, но и науками о Земле — стратиграфией, палеогеографией, тектоникой, геохимией и др. Пренебрежение этими фактами может направить все исследование по неверному руслу.
В 1927 г. в Москве вышла книга М. И. Голенкина «Победители в борьбе за существование». Потом она дважды переиздавалась, что редко случается в научной литературе. В этой книге большое внимание было уделено распространению покрытосеменных в середине мелового периода. Еще в прошлом веке было замечено, что количество листьев покрытосеменных в верхнемеловых отложениях значительно больше, чем в нижнемеловых. Некоторые верхнемеловые захоронения растений иногда почти лишены других растительных остатков, а отпечатки покрытосеменных образуют листовые кровли. В нижнемеловых отложениях эти отпечатки встречаются, но гораздо реже. Внезапное увеличение остатков покрытосеменных в середине мела и породило ту «отвратительную тайну», о которой писал Дарвин в письме Гукеру.
В гипотезах, объяснявших этот факт, никогда не было недостатка. Большей частью они исчезали столь же незаметно, как и появлялись. Гипотезе Голенкина повезло гораздо больше. Ее сразу заметили, и ее сторонников немало до сих пор. Сочувственное изложение идей Голенкина можно найти даже в учебниках.
Голенкин предположил, что покрытосеменные стали «победителями в борьбе за существование» по нескольким причинам, как зависящим, так и не зависящим от них самих. Одним из решающих преимуществ покрытосеменных перед голосеменными и споровыми, населявшими сушу до позднего мела, Голенкин считал способность выносить яркий солнечный свет и сухость воздуха. Эти преимущества не имели большого значения до определенного момента. Раз покрытосеменные появились еще в раннем мелу и особенно не выделялись, а потом внезапно распространились по всей Земле, значит, им что-то помогло. Не могли же частные и местные факторы сработать сильно и одновременно по всей Земле!
Опираясь на некоторые косвенные соображения, Голенкин предположил, что до середины мела Земля была укутана плотными облаками, так что солнечные лучи редко пробивались к земной поверхности. В этой обстановке и появились первые покрытосеменные. Тогда они не имели серьезных преимуществ перед тенелюбивыми и влаголюбивыми голосеменными и споровыми растениями. В середине мела по каким-то космическим причинам облачный покров планеты сильно поредел. Яркие солнечные лучи хлынули на Землю. Вместе с освещенностью возросла и сухость климата. Большинство споровых и голосеменных растений, господствовавших до этого, не смогли приспособиться к революции климата и света. Они вымерли и освободили жизненное пространство покрытосеменным, «детям солнца».
Я не проводил специальных изысканий, чтобы выяснить, насколько заинтересовала гипотеза Голенкина палеоботаников. Может быть, кто-нибудь и высказывался о ней с сочувствием. Во всяком случае это не было правилом. Показательно, что в капитальной сводке А. Н. Криштофовича «Палеоботаника», вышедшей посмертным изданием в 1957 г., книга Голенкина упоминается в списке литературы, а в главе о меловых флорах о его гипотезе не говорится ни слова. Разумеется, отсутствие ссылок можно объяснять по-разному. Бывает ведь и заговор молчания против выдающейся идеи. Я с трудом верю, что в случае гипотезы Голенкина было умышленное игнорирование. Просто она не заинтересовала профессионалов-палеоботаников. О ней все знают и остаются равнодушными. Еще меньше ее популярность за рубежом.
Популярность гипотезы Голенкина поддерживается преимущественно в ботанических кругах, но я ни разу не видел, чтобы ее анализ сопровождался внимательным отношением к материалам исторической геологии. Рассматриваются покрытосеменные вообще, говорится о меловом периоде в масштабе всей Земли. Нет ни рассмотрения конкретных, известных палеоботаникам меловых флор, ничего не говорится об условиях их существования, как они видятся специалистам.
При том равнодушии, которое выказывают палеоботаники к гипотезе Голенкина, может быть, и неуместно много о ней говорить. Все же я решил разобрать ее более подробно, поскольку она показательна с методологической точки зрения. При этом я вовсе не ставлю в вину Голенкину незнание фактов, ставших известными позднее. Такого упрека заслуживают лишь те, кто поддерживает идею Голенкина до сих пор.
Главный дефект гипотезы Голенкина в том, что она приписывает всему миру то, что характерно только для Европы. За ее пределами события в середине мела выглядят иначе. В некоторых местах листовые кровли с покрытосеменными встречаются в верхних горизонтах нижнего мела. В других местах Земли, наоборот, покрытосеменные преобладают в захоронениях только в верхних горизонтах верхнего мела, а до этого главную роль играют хвойные.
Сейчас, когда стратиграфия меловых отложений стала достаточно детальной и поэтому датировки толщ с растительными остатками приобрели определенность, картина расселения покрытосеменных представилась совсем иной, чем в 20-х годах. Если опираться не только на отпечатки листьев, но и на остатки пыльцы покрытосеменных, то картина получится еще менее драматичной. В некоторых районах количество
покрытосеменных действительно резко увеличивается на определенном уровне, но в разных районах соответствующие рубежи оказываются приуроченными к разным интервалам мелового периода.
Судя по длительности эпох мелового периода, процесс завоевания суши покрытосеменными затянулся на многие миллионы лет. Не будем забывать и о том, что до сих пор самые распространенные деревья на Земле — сосна и ель, т. е. голосеменные, а не покрытосеменные растения. Таким образом, разрушен главный компонент прежних взглядов — внезапность и одновременность победы покрытосеменных по всей Земле. Даже в масштабах геологического времени его нельзя считать быстрым. Палеоботаники давно отметили, что флоры в каждом регионе меняются как бы толчками. На большом участке геологического разреза мы наблюдаем примерно одного типа флору, затем на коротком отрезке состав флоры меняется, и снова наступает период спокойствия. Масштабы перемен могут быть различными. Иногда флоры меняются быстро и почти целиком, так что между старой и новой флорами не остается общих видов. В этом отношении флористические границы внутри мела, на которых в том или ином месте Земли покрытосеменные вытеснили предшественников, ничем не отличаются от прочих флористических границ. С некоторыми мы сталкивались в прошлых главах.
Поставив меловые события в один ряд с другими флористическими событиями в истории Земли, мы должны, как мне кажется, не подбирать каждой границе в каждом районе особое объяснение и уж тем более не спешить за помощью к космическим причинам, а попытаться понять общие причины, приводящие к сменам флор. Думаю, что в случае расселения покрытосеменных сработали обычные земные механизмы: перестройки барьеров, смена миграционных маршрутов, климатические перемены и пр.
Меловой период богат крупными событиями в истории лика Земли. Еще в конце юры произошел раскол Гондваны на две части. Западная Гондвана, охватывающая Африку и Южную Америку, отделилась от Восточной Гондваны (Индии, Австралии и Антарктиды). Примерно на рубеже юры и мела, как полагают некоторые тектонисты, Индия стала отходить от Антарктиды. В первой половине мела распад Гондваны продолжался, Очередь дошла до Западной Гондваны, которая стала делиться на Южную Америку и Африку (рис. 22). Этот процесс подробно прослежен по меловым отложениям, сохранившимся в приатлантических частях обоих материков (об этом рассказывалось в главе VI). В позднем мелу образовался Протоатлантический океан. Чем дальше, тем больше хирел великий средиземноморский океан Тетис. Индия отделилась от Мадагаскара и отправилась на север, чтобы войти в новый материк — Евразию. В конце мела Индия уже лежала в тропиках. Антарктида, наоборот, поехала на юг, и ее климат становился все холоднее.
 Рис. 22. Взаимное положение Южной Америки и Африки в раннем мелу (К), середине палеогена (раннем эоцене — EI), начале неогена (раннем миоцене — Ml) и сейчас (в голоцене — Н) по реконструкции Т. ван Анделя, Д. Тиде, Д. Г. Скле Итеру и В. В. Хэю
Рис. 22. Взаимное положение Южной Америки и Африки в раннем мелу (К), середине палеогена (раннем эоцене — EI), начале неогена (раннем миоцене — Ml) и сейчас (в голоцене — Н) по реконструкции Т. ван Анделя, Д. Тиде, Д. Г. Скле Итеру и В. В. Хэю
Может быть, я слишком уверенно расписываю все эти события. В истории гондванских материков и в маршрутах их путешествий (если вообще их допускать) еще немало неясного. Пусть не описанные события, а другие, тоже достаточно крупные, все же так или иначе происходили на Земле в меловое время. С тем, что в мелу происходили серьезные палеогеографические перестройки, никто спорить не будет. И при фиксистском, и при мобилистском прочтении истории приходится допускать, что в это время перестраивалась вся система циркуляции водных, а с ними и атмосферных масс. Должны были меняться и ландшафты, особенно если материкам приходилось ползти поперек параллелей. Ломались зоны растительности, а с ними и экосистемы.
Все эти события отнюдь не были катастрофическими, хотя в отдельных регионах перестройки совершались быстро. Желающие могут пофантазировать, как в обстановке смуты и ломки экосистем покрытосеменные подняли головы, стали одолевать соперников — споровых и голосеменных — и вытесняли их из ландшафтов.
Я не берусь сам рассуждать на эти темы, поскольку мы знаем еще слишком мало. И вообще сейчас здесь надо поменьше рассуждать, а более тщательно собирать и анализировать сохранившиеся документы. Сведения об ископаемых флорах по всем видам растительных остатков, особенно по пыльце и спорам, надо связать с конкретным палеогеографическим фоном, составлять подробные палеофлористические карты, много карт, для всех районов, где сохранились подходящие отложения. Такие исследования ведутся, хотя и в ограниченном масштабе. До последнего времени большая часть карт использовала фиксистские реконструкции палеогеографического фона. Теперь надо на все посмотреть и с мобилистских позиций. В отношении покрытосеменных такую попытку недавно сделали американские исследователи ботаник Р. Рейвн и палеоботаник Д. Аксельрод (он, если помнит читатель, когда-то резко протестовал против мобилистских идей).
Задача реконструкции меловых событий в растительном покрове Земли осложняется тем, что приходится опираться на палеогеографические реконструкции, которые сами зависят от интерпретации палеоботанических данных. Достоверность схемы расположения континентов подлежит палеоботаническому экзамену. Геологические и палеоботанические (и вообще палеонтологические) данные еще предстоит свести в единую непротиворечивую систему. Только тогда можно будет более уверенно судить о том, где были центры формирования отдельных групп покрытосеменных, как и когда шло их расселение. Не выполнив этой работы хотя бы начерно и не построив палеофлористических карт всего мира хотя бы для каждого века мелового периода (таких карт пока нет), нет смысла увлекаться гипотезами о конкретных причинах того, что произошло в растительном покрове Земли в те далекие времена.
Гипотеза Голенкина — не единственная, которой не было суждено выдержать проверку фактами. Новые материалы заставляют усомниться и в других гипотезах, особенно касающихся места и времени возникновения покрытосеменных. Зашатались и гипотезы об облике первых покрытосеменных. Я не хочу сказать, что есть ответы на все эти вопросы. Можно говорить лишь о сокращении числа возможных вариантов ответа на них.
Я уже говорил о том, что многие ботаники пытались найти ответы чуть ли не на все вопросы, глядя на современный растительный мир, и что исходной формой покрытосеменных чаще всего считают древесные растения с крупными одиночными цветками, напоминающими цветки магнолий. Построив на этой основе предполагаемую филогению покрытосеменных, ботаники попытались отыскать и родину этих растений. Оказалось, что семейства и роды, стоящие в нижней части предполагаемого филогенетического древа, не расселены равномерно по всей Земле, а сконцентрированы в отдельных районах. Самое большое их скопление отмечено в Юго-Восточной Азии. Ее-то и нарекли предполагаемой родиной покрытосеменных.
До последнего времени эти взгляды нельзя было непосредственно проверить по палеоботаническим данным. К тому же палеоботаников и ботаников долго сбивали с толку остатки якобы покрытосеменных, которые указывались в юрских и даже триасовых отложениях. Скажем, в юрских отложениях описали пыльцу с тремя бороздами, сходную с пыльцой современной эвкоммии. В триасе нашли отпечатки, сильно напоминающие листья однодольных. Теперь большая часть этих остатков предполагавшихся покрытосеменных из домеловых отложений изучена вновь, и их принадлежность покрытосеменным признана или ошибочной, или очень сомнительной.
С другой стороны, в нескольких достаточно полных разрезах меловых отложений тщательно изучены остатки листьев и пыльцы, принадлежность которых к покрытосеменным достаточно определенна. Об этих работах стоит кратко рассказать. Им посвятили свои статьи палеоботаник Л. Д. Хики и палинологи Д. Дойл, М. Ван Кампо, Б. Люгардон и некоторые другие.
Главная особенность проведенного ими исследования в том, что они широко применили электронно-микроскопическую технику. Еще в прошлом веке, когда в распоряжении ботаников появились неплохие световые микроскопы, было обнаружено, что пыльца растений имеет многослойную оболочку. У разных растений структура слоев оказалась неодинаковой. В последние десятилетия для изучения пыльцы сначала современных, а затем и ископаемых растений стали использовать электронный микроскоп.
Из пыльцевого зерна приготавливают тончайшие срезы, которые потом изучают при огромных увеличениях. Для исследования поверхности пыльцевого зерна на нее напыляют в вакууме тончайший слой золота или другого металла и помещают зерно под сканирующий электронный микроскоп. Максимальное увеличение, применявшееся при изучении пыльцы, достигало миллиона раз. Как отмечал шведский палинолог Г. Эрдтман, который одним из первых применил электронную микроскопию для изучения спор и пыльцы, при таких огромных увеличениях пыльцевое зерно липы, имеющее в диаметре около 40 микрон, увеличивается до 40 м. По нему можно было бы совершать экскурсии. Правда, на практике такими увеличениями не пользуются. Обычно ограничиваются увеличениями в несколько тысяч или в первые десятки тысяч раз. Но и при увеличении в 10 тыс. раз диаметр пыльцы липы будет 40 см, т. е. больше футбольного мяча.
Пыльца одних растений имеет достаточно сложное строение, замысловатый рельеф. Различить разные группы растений по такой пыльце можно и в световой микроскоп. У других растений пыльца гораздо проще. Например, пыльца цикадовых, гинкговых, беннеттитов, некоторых птеридоспермов и покрытосеменных может выглядеть в световой микроскоп почти одинаково — в виде маленькой булочки без заметного рельефа. В этом случае без электронной микроскопии обойтись нельзя. Сделав срезы с пыльцевых зерен и рассмотрев тонкую структуру их оболочек, можно отделить пыльцу покрытосеменных от голосеменных и различить разные группы голосеменных (рис. 23). Именно так и изучили ту пыльцу из юрских и нижнемеловых отложений, которую предположительно считали принадлежащей покрытосеменным.
Оказалось, что первая достоверная пыльца покрытосеменных появляется в нижней половине нижнемеловых отложений. Почти на том же уровне в разрезе появляются и первые достоверные остатки листьев покрытосеменных. Разнообразие листьев и пыльцы довольно быстро увеличивается вверх по разрезу. Д. Доил и Л. Хики проследили этот процесс в нижнемеловых отложениях штатов Виргиния, Мэриленд и Делавэр и составили выразительную схему, которую я воспроизвожу (рис. 24).
Примерно такую же последовательность форм палеоботаники наблюдали и в других местах Земли, хотя в разных местах первую пыльцу покрытосеменных находят не на одном стратиграфическом уровне. Раньше всего она появляется в районах, прилежащих к нынешней Атлантике, — в Англии, на западе Экваториальной Африки. В Австралии это произошло с запозданием примерно в 10 млн. лет. Сравнительно поздно она появляется и в геологических разрезах Юго-Восточной Азии. Таким образом, палинологические данные никак не подтверждают идею о первом появлении покрытосеменных в Юго-Восточной Азии. Более вероятно их африканское происхождение, поскольку именно в Африке, если верить палинологическим данным, быстрее всего и раньше, чем в других местах, нарастало разнообразие покрытосеменных (рис. 25). Именно здесь они впервые становятся заметным компонентом растительности. В отложениях аптского века (второго от конца мелового периода) доля пыльцы покрытосеменных в спорово-пыльцевом комплексе достигает здесь 5 %. В других местах такого процента покрытосеменные достигают позже. Впрочем, данные на этот счет еще предстоит свести воедино и как следует проанализировать.
 Рис. 23. Структура оболочки спор и пыльцы у разных растений — прогимноспермов (предголосеменных), голосеменных и покрытосеменных, по данным электронной и световой микроскопии; а — современные гнетовые, юрский Eucommiidites (слева вверху — общий вид пыльцевого зерна Eucommiidites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); б — некоторые сережкоцветные, розоцветные, сложноцветные и другие покрытосеменные; в — многие покрытосеменные, в том числе древнейшие (в углах наверху показан общий вид пыльцевого зерна нижнемелового Clavatipolienites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); г — прогимноспермы, кипарисовые, араукариевые и другие голосеменные; д — цикадовые (левая часть рисунка), птеридоспермы, кордаитовые, сосновые, подокарповые (по Д. Дойлу, М. ван Кампо и В. Люгардону)
Рис. 23. Структура оболочки спор и пыльцы у разных растений — прогимноспермов (предголосеменных), голосеменных и покрытосеменных, по данным электронной и световой микроскопии; а — современные гнетовые, юрский Eucommiidites (слева вверху — общий вид пыльцевого зерна Eucommiidites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); б — некоторые сережкоцветные, розоцветные, сложноцветные и другие покрытосеменные; в — многие покрытосеменные, в том числе древнейшие (в углах наверху показан общий вид пыльцевого зерна нижнемелового Clavatipolienites с двух сторон; слева внизу — вид его оболочки в разрезе в электронный микроскоп); г — прогимноспермы, кипарисовые, араукариевые и другие голосеменные; д — цикадовые (левая часть рисунка), птеридоспермы, кордаитовые, сосновые, подокарповые (по Д. Дойлу, М. ван Кампо и В. Люгардону)
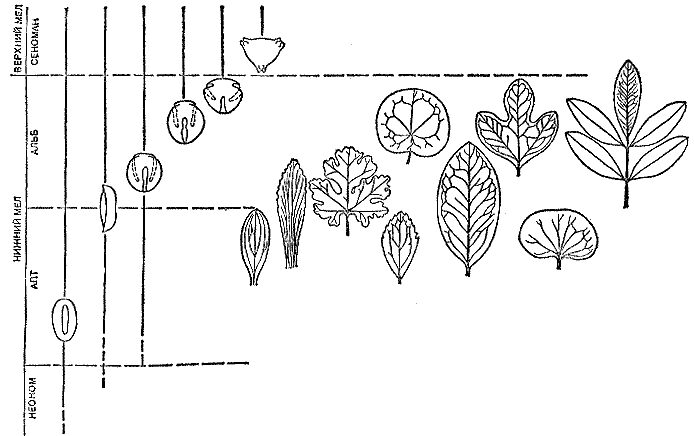 Рис. 24. Распространение в меловых отложениях Америки, Африки и Западной Европы основных типов пыльцы и листьев покрытосеменных; во второй колонке слева — ярусы (века) мелового периода (по Д. Дойлу и Л. Д. Хики с дополнениями)
Рис. 24. Распространение в меловых отложениях Америки, Африки и Западной Европы основных типов пыльцы и листьев покрытосеменных; во второй колонке слева — ярусы (века) мелового периода (по Д. Дойлу и Л. Д. Хики с дополнениями)
Сейчас пыльца меловых покрытосеменных интенсивно изучается в разных странах, и, надо полагать, нынешние представления о ее распространении будут существенно меняться. Однако едва ли мы когда-нибудь вернемся к гипотезе, что их колыбелью была Юго-Восточная Азия. Дело в том, что в течение мела и большей части палеогена Юго-Восточная Азия входила в единый палеогеографический пояс, примыкавший к океану Тетис (или его остаткам) и протягивавшийся далеко на восток вплоть до Западной Европы. Еще в 1968 г. голландский палинолог И. Муллер обнаружил, что палинологические комплексы палеогена (точнее, эоцена) Калимантана и Западной Европы очень сходны. Уже из этого можно заключить, что нынешняя Юго-Восточная Азия в ботаническом отношении — остаток обширной территории, когда-то заселенной примерно однотипной растительностью. Теперь она скорее дом для престарелых, убежище для тех, кого климатические перемены выгнали из других мест.
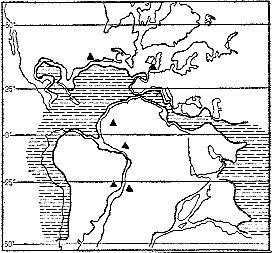 Рис. 25. Места находок древнейшей (доальбской) раннемеловой пыльцы покрытосеменных; жирной линией показана древняя береговая линия
Рис. 25. Места находок древнейшей (доальбской) раннемеловой пыльцы покрытосеменных; жирной линией показана древняя береговая линия
В последние годы тщательно изучались и листья древнейших покрытосеменных из нижнемеловых отложений. Если не обращать внимания на тонкие особенности жилкования, то эти листья можно сблизить с современными семействами и даже родами. Палеоботаники долго так и поступали. Кажущаяся близость раннемеловых и современных покрытосеменных была одной из причин существовавшего убеждения, что раннемеловая летопись этих растений свидетельствует лишь об их расселении, а не о первых шагах эволюции. Собственно происхождение покрытосеменных относили к юре, триасу или даже перми.
Как мы уже видели, палинологические материалы не поддержали этой гипотезы. Внимательное изучение отпечатков листьев подтвердило палинологические наблюдения. Оказалось, что если принять во внимание тонкие детали жилкования, то уже нельзя сблизить раннемеловые листья не только с родами, но и с семействами современных растений. Это что-то совсем иное, а сходство с современными родами и семействами чисто внешнее. Таким образом, более правдоподобно предположение, что в раннемеловую эпоху происходило не просто расселение форм, появившихся гораздо раньше, а сама выработка новых форм. Первые представители семейств, доживших до наших дней, появились позже, лишь в позднем мелу. Современные роды имеют еще более скромную историю. Немногие появились в конце позднего мела, а остальные лишь в палеогеновом и неогеновом периодах.
В литературе обсуждались и ландшафтные условия, в которых произошли покрытосеменные. Среди ботаников и палеоботаников довольно популярна гипотеза, что покрытосеменные спустились с гор. Ее впервые независимо друг от друга выдвинули В. А. Вахрамеев и Ч. Арнольд. Гипотеза «горной колыбели» действительно позволяла многое понять. В горах нет условий для захоронения растительных остатков. Накапливающиеся здесь толщи рано или поздно перерабатываются и переоткладываются. Захоронение растительных остатков может быть устойчивым только в низинах. Исключения крайне редки. Естественно, что от растений, населяющих горы, нам достается преимущественно пыльца. Пока раннемеловые покрытосеменные соотносились с современными семействами и родами, часто высокоорганизованными, горная гипотеза позволяла объяснить, почему мы не находим в мезозое по-настоящему примитивных покрытосеменных.
После того как раннемеловые покрытосеменные оказались более своеобразными, чем считалось раньше, и появились основания полагать, что именно в раннем мелу они сделали первые эволюционные шаги, потребность в горной гипотезе стала падать. Против нее были выдвинуты и прямые возражения, в том числе методологические. Она, как заметил В. А. Красилов, «полностью снимает контроль палеоботанических фактов и открывает простор для чисто умозрительных построений. Можно допустить существование цветковых в пермо-триасе, а если такой возраст покажется недостаточно древним, то отодвинуть их возникновение в карбон или даже докембрий… Эта свобода действия, вероятно, в немалой степени способствовала исключительной популярности теории горного происхождения цветковых, которую не смогли поколебать отдельные критические высказывания…».
Такие критические высказывания, как собственные, так и принадлежащие другим специалистам, В. А. Красилов и суммировал. Я остановлюсь лишь на некоторых из них и добавлю свои соображения. Во-первых, те горы, которые представлялись возможной родиной покрытосеменных, возникли геологически слишком поздно. В тех местах, где мы находим древнейшие остатки покрытосеменных, гор по соседству не было. Во-вторых, долго и безвылазно сидеть на горах покрытосеменные едва ли бы смогли. В течение юры и первой половины раннего мела (т. е. до появления первой пыльцы покрытосеменных в геологической летописи) климатические колебания вызывали движение поясов растительности по склонам гор. В периоды похолодания покрытосеменные имели все возможности спуститься в низины и попасть в устойчивые захоронения хотя бы в виде пыльцы. Далее, те современные горные формы, которые якобы сохраняют архаичность и свидетельствуют о горном происхождении первых покрытосеменных, известны в палеогеновых и неогеновых отложениях, накопившихся в низинах. Наконец, если покрытосеменные действительно появились в горах низких широт, то можно ожидать их спуск в низины в более высоких широтах. Тогда, чем ближе к экватору, тем позднее должны появляться в разрезе остатки покрытосеменных. На самом деле мы видим обратную закономерность.
К сожалению, от древнейших покрытосеменных нам пока достаются только пыльца и отпечатки листьев (фиг. XV). Ботаникам же больше всего хочется узнать, какими были первые цветки. Об этом геологическая летопись пока умалчивает. Наиболее древние органы размножения относятся уже к концу раннего мела и представлены не цветками, а плодами (фиг. XVI). Если бы эволюция покрытосеменных начиналась с крупных цветков, как у магнолии (так считают многие ботаники), то мы находили бы вместе с листьями и остатки крупных лепестков. Когда отцветает магнолия, то земля бывает усыпана побуревшими лепестками. Ничего похожего на лепестки крупных цветков палеоботаники не находят в нижнемеловых отложениях. Есть и другие палеоботанические свидетельства того, что первые покрытосеменные имели не крупные, двуполые и одиночные цветки, а мелкие, однополые, собранные в кистевидные соцветия. Не исключено, что и название «цветки» к ним мало подходило, поскольку у них, может быть, не было лепестков, так что они больше напоминали по внешнему виду органы размножения голосеменных.
Какая группа голосеменных растений дала начало покрытосеменным, мы все еще в неведении. Многие свойственные покрытосеменным признаки появляются независимо у разных голосеменных. У некоторых даже образовывалось подобие завязи. В качестве возможных прародителей перебывали чуть ли не все группы высших растений. Высказывалось предположение, что происхождение покрытосеменных сопровождалось гибридизацией между разными группами. Речь не обязательно идет об обычной половой гибридизации. Генетическую информацию от одной группы к другой могли переносить, скажем, вирусы. С точки зрения генетики в этом нет ничего невероятного.
Впрочем, как ни интересно обсудить возможных предков покрытосеменных и пути их эволюции, здесь не место этим заниматься. Иначе нам придется залезать в дебри морфологии и систематики растений. К сказанному я хочу добавить лишь одно. Проблема происхождения покрытосеменных настолько интересует ботаников, что они спешат ее решить, не дожидаясь, пока будут расшифрованы палеоботанические документы. Странным образом, интерес к проблеме почти не стимулирует исследователей заняться этими документами как следует. О происхождении покрытосеменных рассуждают сотни людей; статьи, посвященные умозрительному разбору предлагавшихся гипотез, исчисляются тысячами. Палеоботаников же, которые занимаются конкретными документами по древнейшим покрытосеменным, тщательно ищут и изучают их остатки в меловых отложениях, анализируют распределение в пространстве и времени уже открытых остатков, буквально единицы. Их можно пересчитать по пальцам. Если бы несколько процентов энергии, затрачиваемой на умозрительные дискуссии, направить на палеоботанические исследования, то, можно надеяться, многие аспекты проблемы стали бы быстро проясняться.
Глава IХ
ЧТО ЕСТЬ ИСТИНА В ИСТОРИИ ЗЕМЛИ?

В предыдущих главах шла речь о далеком прошлом Земли, об истории растительной жизни на ней. Не раз говорилось о том, что были-де какие-то представления о прошлом, а потом они оказывались ошибочными. В некоторых случаях ошибка была достаточно очевидна. Например, когда-то приняли остатки одноклеточных водорослей за пыльцу покрытосеменных. Был допущен просчет в классификации, который никто не берется оспаривать. В других случаях положение иное. Когда-то мысль о движении континентов просто не приходила серьезным ученым в голову. Потом одни стали мобилистами, а другие остались фиксистами. Потом к тем и другим прибавились сторонники расширения Земли. В этой книге читатель встретил сочувствие к мобилистским идеям, но скептик может заметить, что, наверное, и фиксисты не будут зря настаивать на заведомо чепуховых идеях. Такие же скептические вопросы могут возникнуть и по отношению к прочим мыслям о прошлом, которые разделяются всеми исследователями. Самый заядлый скептик может вообще заявить: «Кто все это видел? Кто мерил температуру на полюсе в раннем карбоне? Как можно доказать, что одна реконструкция прошлого верна, а другая ошибочна? Прошлого не вернуть, а значит, нечего упрекать кого-то в ошибках». Приходится признать правоту таких слов. Что-то окончательно доказать относительно прошлого действительно уже невозможно, ибо прошлое в самом деле невозвратимо. Все же из этого не следует, что все гипотезы о давно прошедших событиях равновероятны. Больше того, опыт истории науки убеждает, что и при изучении настоящего ничего нельзя окончательно доказать. Об этом хорошо писал П. А. Флоренский: «Век XIX ознаменовался в самом начале своем критикою знания, XX же производит критику методов знания. Итог этой критики — тот, что в настоящее время не нуждается уже в подробном объяснении мысль, высказанная ранее блестящим Стенли Джевонсом, а именно что «почти каждая проблема в науке принимает форму балансирования вероятностей»… Только бесконечный опыт мог бы дать достоверное знание; вероятность же в нашем знании и есть отражение потенциальной бесконечности опыта. Это относится и к физике, и к астрономии, и к химии, и в особенности к историческим дисциплинам».
О математическом выражении большей или меньшей вероятности какой-либо из конкурирующих гипотез пока не может быть речи. Приходится говорить о «субъективной вероятности» (в начале века ее называли «нравственной вероятностью»), о мысленном взвешивании каждым исследователем всех «за» и «против» определенной гипотезы. Взвешивая аргументы и контраргументы, недостаточно просто знать относящиеся к делу факты. Надо еще разбираться в обращении с фактами, в том, что называется принципами научного исследования. Об этих принципах и хотелось бы немного поговорить, когда читатель уже познакомился со «следами трав индейских».
Есть еще одна причина, почему уместен этот разговор. В последние десятилетия в географии, геологии, а до этого и в биологии начали распространяться антиисторические тенденции. В биологии они появились давно, почти сразу после становления эволюционной теории. Тогда биологи с увлечением рисовали многочисленные филогенетические древеса, а в историческом анализе видели цель и смысл чуть ли не любого биологического исследования. Массовые увлечения часто вызывают встречные тенденции. Так и случилось. В противовес историзму в биологии стал развиваться структурализм: «Будем изучать структуру, а с историей разберемся потом».
Тот же процесс начался в геологии сравнительно недавно. Бесконечные дискуссии о возникновении и истории континентов и океанов, происхождении нефти, механизмах горообразования и подобных вопросах вызвали стремление обойтись без этих проблем вообще. Были выдвинуты новые программы развития геологии, в которых исторической геологии отводилась роль завершающей главы, до которой надо добираться тогда, когда главные практические вопросы уже решены. Дескать, нет смысла спорить о происхождении нефти, надо учиться ее искать и добывать.
Дискуссия «историков» и «структуралистов» заняла уже не одну сотню страниц опубликованных статей и даже книг. Если правы структуралисты, то всё, о чем написано в этой книге, — развлекательное приложение к деловой части палеоботаники, которая вообще-то должна решать вопросы сопоставления разрезов и не тратить время на гипотезы о том, как вышли растения на сушу, как завоевали ее и что было потом с зеленым покровом Земли.
О том, какой практический смысл имеет палеоботаника, в том числе и география вымерших растений, пойдет речь в последней главе. А сейчас вернемся к достоверности реконструкции прошлого, поскольку именно из-за недостоверности выводов больше всего нареканий на историзм. Я попробую показать, что отказ от историзма даже на начальной стадии исследования неизбежно ведет к грубым ошибкам. Но сначала надо познакомиться с тем, что предлагают структуралисты.
Вкратце структуралистский подход к проблемам, которым посвящены предыдущие главы, был бы примерно таким. Предлагается описывать осадочные толщи в том виде, как они предстают перед исследователем, не обращая внимания на гипотезы об условиях осадконакоплення, образе жизни тех животных и растений, остатки которых попадаются при описании. Потом надо по составу пород и спискам найденных в них окаменелостей сопоставить разрезы и оконтурить отдельные геологические тела. Опираясь на полученную схему строения земной коры, можно обрисовать площади распространения тех или иных ископаемых организмов по разным интервалам разреза. Лишь затем можно вводить в эту картину понятие времени, обращаться к реконструкции истории. Короче говоря, главный лозунг формулируется так: сначала изучение современной структуры земной коры, затем реконструкция истории, сначала пространственные характеристики, затем временные.
Все выглядит разумно. Мы и в самом деле вроде не можем, глядя на слои горных пород, наблюдать время, а выводим временные отношения слоев из их пространственных отношений. Чтобы окончательно в этом убедиться, надо понять, что такое время. Здесь-то и начинаются главные трудности структурализма, как, впрочем, и историзма. Вроде все понимают, что это такое, а когда дело доходит до введения времени в теорию естественнонаучных исследований, так все становится куда сложнее. Легче всего проблема решается тогда, когда есть часы, с показаниями которых можно сверять отдельные стадии процессов. А если часов нет? Где тогда взять понятие времени?
Изучая прошлое Земли, мы оказываемся практически без часов. О длительности геологических эпох можно составить представление с помощью радиологических методов (по распаду радиоактивных элементов). Этот метод появился еще в начале века. В 30-х годах к нему еще относились с недоверием и окончательно приняли лишь в послевоенные годы. Радиологические датировки дали возможность составить представление о длительности геологических периодов, с их помощью иногда сопоставляют разрезы. Для стратиграфии докембрия роль «абсолютных датировок» очень велика. Они широко применяются и для датировки новейших отложений, возрастом в первые десятки тысяч лет, где можно опереться на радиоуглеродный метод. В остальном «абсолютная геохронология» играет подсобную роль в определении относительного возраста геологических тел. Показательно, что современные стратиграфические представления о кембрийских и более молодых толщах сложились до появления абсолютной геохронологии. Еще в начале 30-х годов в капитальной сводке А. Грэбо «Принципы стратиграфии» приводились общие оценки возраста Земли от 48 млн. до 7 млрд. лет. Грэбо пришел к выводу, что подобные предположения преждевременны и почти бесполезны. Ведь при расхождении в оценках длительности подразделений почти в 150 раз сама шкала периодов, эпох и веков оставалась одной и той же.
Можно сделать вывод, что, стало быть, понятие времени действительно не слишком важно для геологии, раз такие противоречия во взглядах на возраст Земли не играли практической роли. Такой вывод вовсе не обязателен. Геология обходилась без часов, а понятие времени широко использовала. Что же за время такое без часов, как его представить?
Вкратце концепция времени без часов выглядит так. В мире без часов наблюдатель вынужден опираться на свое собственное время, которое А. Бергсон назвал психологическим и которое упрощенно можно представить как ощущение собственной изменчивости. В «психологическом времени» каждый человек без труда устанавливает отношения «раньше» и «позже». Далее наблюдатель заметит, что окружающие его объекты тоже меняются. Время этих объектов выступит как их изменчивость. Обычно мы употребляем понятие изменчивости, говоря о различиях объектов одного класса. То же понятие, отнесенное к одному объекту с условием, что порядок его изменчивости задается наблюдателем, его «психологическим временем», дает возможность ввести индивидуальное время объекта.
Наблюдатель отметит, что изменчивость объектов-индивидов различна, т. е. различны процессы, происходящие с разными объектами. В соответствии с классами объектов можно выделить классы процессов и тем самым классы времен. Учение о разнообразии объектов нередко совершенно точно называют типологией. Тогда концепцию времени, отражающую разнообразие объектов в их изменчивости, можно назвать типологической концепцией времени. Примерно такой смысл вкладывал В. И. Вернадский в понятие природного времени.
Мы можем отвлечься от своеобразия природных объектов и рассматривать их как некие физические тела, отличающиеся простейшими физическими свойствами — массой покоя, твердостью и т. п. Тогда будет упрощена и их изменчивость, которая обратится в физическое время. Наконец, можно говорить об абстрактной изменчивости вообще некоего абстрактного объекта, материальной точки классической механики. Время такого объекта, выражающееся в изменении его координат, будет ньютоновским абсолютным временем.
Сравнивая изменчивость самых разных объектов, т. е. их времена, наблюдатель заметит, что разные свойства ведут себя неодинаково. Одни свойства как бы попадают на записывающее устройство, так что изменчивость в этих свойствах записывается. Соответствующий аспект изменчивости объекта, его времени, закрепляется в структуре объекта — это темпофиксация. Так записывается изменяющаяся толщина дерева в его годичных кольцах (слоях прироста). Изменчивость других свойств или отделяется вместе с отделяющимися частями объекта (как шкура змеи при линьке), или оставляется на окружающих объектах (например, следы на песке сохранят усиливающуюся усталость пешехода) — это отделение времени, темпосепарация. Изменчивость третьих свойств никак не фиксируется, исчезает без следа. Запах первых на Земле цветов мы, наверное, уже не узнаем — это «растворение» времени, темподесиненция.
Введя понятия темпофиксации, темпосепарации и темподесиненции, можно понять главные проблемы, встающие перед исследователем прошлого. Он должен прочесть все записанное при темпофиксации, собрать и воссоединить все разбросанное темпосепарацией и найти способы реконструировать свойства, подвергшиеся темподесиненции. Насколько велики эти трудности, объяснять не требуется, а обойти их чисто структуралистским анализом нельзя по следующей причине.
Когда четыре мушкетера защищали бастион, они разложили ружья и бегали от одного ружья к другому. Ларошельцы решили, что в бастионе засел целый отряд, и послали за подмогой. Им не пришло в голову, что один человек стреляет поочередно из нескольких ружей в разных местах. Следопыта, который число отпечатков лап примет за число зверей, впору поднять насмех. Считать надо уметь. Пренебрежение темпофиксацией и темподесиненцией ведет именно к таким ошибкам в счете. Увидев на песчаниковой плите правильную цепочку следов, оставленных динозавром, палеонтолог не имеет права рассматривать каждый след как самостоятельный экземпляр, как он это сделает, найдя несколько раковин моллюсков. Каждый моллюск может оставить лишь одну раковину, а следов от динозавра может остаться сколько угодно.
Со структурной точки зрения множество слоев, заполнивших впадину, — это множество самостоятельных геологических тел. Если же помнить, что слои принадлежат одной впадине, фиксировали ее время и теперь отражают ее изменчивость, то мы уже не сможем считать слои независимыми. Это последовательные следы видоизменения одного объекта, а не множество независимых объектов. Для грамотного первого описания разреза геолог должен помнить об этом. Увидев слоистую толщу, он почти сразу заключает о ее осадочном происхождении и соответственно строит стратегию описания. Пусть самое первое его восприятие было действительно восприятием структуры, но дальше он уже будет действовать с генетической гипотезой в руках, поскольку порядок описания и дальнейшего исследования слоистой осадочной толщи совсем иной, чем расслоенной магматической интрузии.
Осознание необходимости исторических реконструкций на первых же шагах исследования объектов прошлого еще не означает знания, как это делать. Опять же не буду докучать читателю долгими рассуждениями, а сразу перейду к выводу. При реконструкции прошлого используется тот же принцип («типологических экстраполяции»), что и при изучении современных объектов. Мы говорили, что у сосны пыльца с двумя мешками, Это не значит, что ботаники собрали всю пыльцу со всех сосен. Они обошлись выборочными спорангиями на выборочных деревьях, а дальше экстраполировали данные по нескольким спорангиям одного дерева на все его спорангии, а данные по нескольким экземплярам вида — на весь вид.
Точно так же поступает химик, утверждая, что молекула воды состоит из двух атомов водорода и одного атома кислорода. Никто не требует проверять это утверждение на всех молекулах. Мы открываем кран водопровода и знаем, что течет именно вода, а не пиво, не обращаясь к химическому анализу. Это тоже типологическая экстраполяция (в логике подобные суждения называют неполной индукцией, но здесь не место разбирать различия между обоими понятиями).
Несколько упрощая дело, можно сказать, что типологическая экстраполяция — это суждение о всех представителях данного класса объектов по результатам изучения выборочных представителей. Без этих суждений развитие науки моментально остановится. Можно показать, что все ошибки в науке (если исключить прямые ошибки в наблюдении) — это неверные типологические экстраполяции. Что касается наших суждений о прошлом, то это тоже типологические экстраполяции.
Мы говорим, что на плитке породы отпечатался лист и что это не игра природы. Выдвинув такую гипотезу (обычно она проскакивает незамеченной в мозгу палеоботаника, и он сразу уверенно относит отпечаток к листьям), мы можем ее проверить. Допустим, этот лист похож на лавровый. Мы знаем особенности жилкования современных лавровых листьев и строение его кожицы. Появляется возможность для типологических экстраполяции. Мы ищем на отпечатке характерный рисунок жилок и находим его. Если на отпечатке есть угольная корочка, то можно сделать препарат кутикулы и подтвердить определение.
Убедившись, что все признаки совпадают, мы делаем дальнейшие типологические экстраполяции. Можно начать искать в породе пыльцу лавра или искать другие подтверждения, что климат в том месте, где мы нашли отпечаток, был подходящим для лавра. Так шаг за шагом идет исследование. Каждый шаг дает основу для новых типологических экстраполяции, которые мы стараемся проверить. Иногда экстраполяции не оправдываются. Значит, неверно был выбран тот класс объектов, в сфере действия которого допустимы экстраполяции. Надо или относить объект, не оправдавший ожиданий, к другому классу, или менять саму классификацию (точнее, типологию).
Ошибки исторических реконструкций и ошибки в изучении современных объектов, как можно видеть, проистекают из одного источника — или плохой типологии объектов, или неправильного помещения данного объекта в хорошую типологию, или того и другого вместе. Кстати, то, что называют ошибкой наблюдения, проистекает из того же источника, о котором мы часто забываем.
К проблемам типологии, т. е. как лучше всего расклассифицировать и расчленить на части объекты исследования, нередко относятся с пренебрежением. По поводу К. Линнея рассказывают анекдот, что он, дескать, настолько был одержим манией все классифицировать, что не обошел вниманием даже туалеты жены. В биологии к систематикам относятся как к специалистам второго сорта, которые по недоразумению не занимаются молекулярными и тому подобными сверхсовременными исследованиями. Часто забывают, что любое тончайшее молекулярное исследование не стоит и ломаного гроша, если не знать того круга объектов, в пределах которого можно смело экстраполировать полученные сведения. Если же вспомнить, что львиная доля молекулярно-биологических исследований ведется на вирусах и бактериях, то становится не вполне понятным, из-за чего поднимают шум вокруг полученных результатов. Кто сказал, что полученные сведения имеют отношение к гораздо более высокоразвитым организмам, включая человека? На каком основании сплошь и рядом делаются столь широкие экстраполяции?
Читатель может упрекнуть меня и за слишком длинный экскурс в теоретические вопросы, и за не относящиеся к теме книги примеры, которые не раз приводились в этой главе? Все это сделано умышленно и по следующим причинам. Популярные книги, посвященные геологии, биологии, палеонтологии, обычно содержат результаты исследований, реже рассказывают о методах работы и еще реже о наиболее общих принципах, которыми руководствуются исследователи. Да и сами специалисты часто с пренебрежением относятся к анализу этих принципов, считая их излишним мудрствованием. Вести конкретное научное исследование, не задумываясь над фундаментальными научными принципами, можно лишь до поры до времени, пока не возникнут глубокие разногласия между исследователями или пока сама работа не коснется наиболее фундаментальных проблем.
Мне хотелось бы вступиться за «мудрствования», потому что попытки решить проблемы простым накоплением фактов не приводят к успеху. Люди убеждены, что если получен факт против какой-либо теории, то тем хуже для теории. Трудно представить противоположный вывод: тем хуже для факта. Тех, кто игнорирует этот факт, обвиняют в упрямстве и даже нечестности. Научная дискуссия перерастает в личную неприязнь и даже враждебность между учеными.
Среди исследователей прошлого Земли такие случаи не редкость. Люди не замечают, а часто просто не знают, что источник недоразумения не обязательно в моральных качествах противника, не в нежелании считаться с фактами, а в непонимании, что такое факт, каково его соотношение с теорией, что такое научная теория, что она может утверждать, а чего не может. Нельзя требовать от каждого исследователя, чтобы он был специалистом в философии науки и легко ориентировался в философской проблеме «что есть истина». Все же некоторое знакомство с современной философией вообще и философией науки в частности могло бы пусть не привести к решению самих проблем, а хотя бы правильнее поставить их.
Вернемся ненадолго к движению континентов и покрытосеменным. На чем основана уверенность фиксистов, что континенты не могли двигаться? Одни указывают на унаследованные структуры земной коры, например долгоживущие разломы, которые, как считается, уходят глубже того слоя, по которому скользят континенты. Если континенты движутся, то разлом не может держаться на одном месте. Надо принимать что-то одно: или движение континентов, или длительное существование на одном месте глубинных разломов. Другие указывают на то, что нет таких физических сил, которые могли бы сдвинуть о места континент и тащить его на тысячи километров, не раздробив на множество кусков. Ведь толщина континента — первые десятки километров, а ширина — тысячи километров. В масштабах Земли континент — кусок пленки, и тащить ее надо не по маслу.
Сторонники мобилизма тоже не могут себе чего-то представить без обращения к движению континентов: сходство очертаний и геологического строения материков по разные стороны от океанов, полосовые магнитные аномалии, распространение одних и тех же растений в Антарктике и Индии в позднем палеозое и многое другое.
Вдумаемся теперь в слова «не могу себе представить». По существу это означает несогласие с какой-то типологической экстраполяцией. Скажем, мы знаем свойства горных пород и «не можем себе представить», чтобы тонкая пленка континента могла скользить по более глубоко лежащим породам. Такая типологическая экстраполяция кажется недопустимой. Других она не пугает, а почему? Они считают, что там, на глубине, породы имеют иные свойства. Это тоже типологическая экстраполяция.
Кто же лучше разбирается в типологии горных пород, какие вообще экстраполяции здесь допустимы и почему? То же и с растениями верхнего палеозоя. Почему одни и те же виды не могли в палеозое заселять тропики и Южный полюс? Только из-за того, что современные растения так не расселяются? Мы приписываем палеозойским растениям свойства современных, т. е. опять же делаем типологическую экстраполяцию. Справедлива ли она?
Теперь вспомним покрытосеменных. Нашли в Африке древнейшую пыльцу с подходящим строением оболочки. Разве из этого следует с необходимостью, что такой пыльцы не было у голосемянных? И кто сказал, что у древнейших покрытосеменных не могло быть какой-то совсем иной пыльцы, например похожей на пыльцу хвойных? Единственный возможный ответ на эти вопросы — «до сих пор мы такого не встречали», Что ж из того? Раньше мы не встречали растений с папоротниковидными листьями и семенами. Теперь о них (птеридоспермах) знает каждый студент-ботаник. Снова мы опираемся на типологические экстраполяции, которые вообще-то вовсе не обязательны.
Еще до того как палеоботаники изучили древнейшие меловые покрытосеменные, нашли и посмотрели в электронный микроскоп их пыльцу, ботаники строили филогенетические отношения между разными покрытосеменными, познакомившись лишь с современными формами. И сейчас, после этих находок, в филогенетике покрытосеменных главную роль играют не палеоботанические аргументы, а наблюдения над современными растениями. В их формах, структуре, химическом составе видят надежных темпофиксаторов, несущих в себе следы своей истории. Следы-то, может быть, они и несут, да читать их можно в разном порядке. Кто из ботаников как следует проанализировал принципы, позволяющие отличить правильное прочтение следов от ошибочного? Не было такого анализа, а были лишь споры по конкретным вопросам: какими были первые цветки, листья, стволы, где покрытосеменные появились, как расселялись и т. д.
Скучно становится читать материалы этих дискуссий. Невозможно отделаться от впечатления, что наблюдаешь ничейную позицию в шахматной игре, когда партнеры, не искушенные в теории шахмат, продолжают двигать фигуры. Этой аналогией я не хочу сказать, что не вижу смысла и в самих проблемах, и в тех конкретных исследованиях, которые с ними связаны. Проблемы существуют, они интересны, и исследования по ним осмысленны, но, когда от наблюдений переходят к выводам, полезно еще раз вспомнить о пробелах в наших знаниях. Надо представить себе и масштабы геологической истории, и объем геологической летописи. Надо осознать, как мало еще мы успели в ней прочесть и как много утратилось навсегда. Полезно вспомнить ошибки наших предшественников и то, как они настаивали на взглядах, кажущихся нам теперь наивными.
Поразмышляв не спеша обо всем этом, мы поймем, что еще не доросли до окончательных выводов, что прочесть полуистлевшие и разрозненные страницы можно и так и этак. Конечно, трудно действовать, не имея хорошей рабочей гипотезы. Против рабочих гипотез, пусть одной, кажущейся наиболее приемлемой, нет смысла и возражать. Принимать же гипотезу за нечто окончательно установленное — не выражение принципиальности и не научная смелость, а лишь выражение самоуверенности.
Глава Х
ВЗГЛЯД В БУДУЩЕЕ

Прочтя предыдущую главу, одни из читателей могут сказать, что я сгущаю краски и преувеличиваю трудности реконструкции далекого прошлого. Ведь, несмотря ни на какие трудности, мы очень много узнали об истории Земли и жизни на ней. Другие читатели могут сделать вывод, что если все так сложно, то где гарантия истинности сказанного в первых восьми главах книги. Да и вообще стоит ли тратить время на реконструкцию прошлого, если все выводы ненадежны?
Мне бы не хотелось, чтобы меня поняли таким образом. Я как раз старался показать, но, может быть, сделал это не лучшим образом, что расшифровка истории не только возможна, но и необходима.
С расшифровкой истории все обстоит как с познанием вообще. Исследовательская работа теряет смысл или становится разделом технологии, если ученый не верит в свою способность накапливать частицы истины. И при этом он будет неосторожен, если решится перечислять, какие истины им установлены. Так, в общежитии мы знаем, что есть долгожители, но всегда рискованно предрекать долгую жизнь молодому человеку. Кто знает, как повернется его судьба! Среди научных истин есть будущие долгожители, но рискованно называть ту, которой предстоит долгая жизнь. Мы уверены, что каким-то из нынешних идей предстоит сохраниться в веках, но каким — об этом узнают наши потомки.
Смысл занятия историей не только в ее научной необходимости. В конце концов обращение к истории удивительно интересно. Профессиональные палеоботаники (сужу об этом по себе) часто забывают, что перед ними остатки давно ушедших живых существ. Думаю, что этим грешат все исследователи прошлого. Привычная рутинная работа с образцами, химикалиями, аппаратурой, описаниями разрезов приводит к тому, что теряется ощущение документов прошлого. Так врач, выслушивающий массу пациентов, порой перестает видеть в них живых людей с их болью, переживаниями и заботами. Но это плохой врач. Хороший обо всем этом не забывает.
Палеоботаник тоже никогда не должен забывать, что тот лист, оттиск которого остался на плитке сланца, когда-то прорезался из почки, рос, был зеленым и трепыхался на ветру. По его жилкам текли соки, он ловил солнечный свет. Потом все кончилось. Лист расстался с веткой, немного пролетел и упал в воду. Его затянуло илом. Миллионы лет он лежал в темноте, пока при ударе молотка не увидел снова солнечный свет. Специалист должен заставлять себя помнить обо всем этом и видеть зелень в обугленном остатке листа, теплое, голубое море в пласте известняка. Палеонтолог должен почаще приглядываться ко всему, что еще живет. Лишь положив рядом отпечаток и только что сорванный лист, осознаёшь, как много ушло и как интересно все это восстановить.
С тем, что это интересно, мало кто будет спорить, но надо ли? Палеонтолог, очутившийся в незнакомой компании неспециалистов, часто слышит вопрос: «Зачем все это изучать?» Надеюсь, что хотя бы частью я ответил на него в предыдущих главах. Палеонтология настолько тесно связана с геологией, что в нашей стране палеонтологов готовят только на геологических факультетах.
Труд палеонтолога часто незаметен, как труд хорошей домашней хозяйки. Недальновидные работники геологической службы не раз предпринимали поход на палеонтологов, лишая их условий для работы. Известны случаи, когда на палеонтологических исследованиях экономили тысячи рублей, а потом теряли миллионы, поскольку листы геологической карты оказывались непригодными для поисковых работ, а глубокие и дорогостоящие скважины не помогали разобраться в геологическом строении района.
Я совсем не уверен, что люди становятся палеонтологами, имея в виду только практический смысл палеонтологии. Если человека действительно интересуют ископаемые животные и растения, то именно этот интерес и приводит его в палеонтологию, а к практическим потребностям он подстраивается потом. Космонавт К. П. Феоктистов говорил на одной из пресс-конференций: «Я убежден, что, готовясь к плаванию на своих каравеллах, Христофор Колумб в какой-то степени втирал очки королю и королеве, когда говорил о заокеанских сокровищах… Он просто искал и хотел найти нечто новое. Любознательность и любопытство были и остаются одним из важнейших рычагов человеческого прогресса».
Приложение. Таблицы

Фиг. I. Проблематичный организм алданофит (Aldanophyton) из нижнего кембрия Сибири; образцы из коллекции проф. А. Р. Ананьева (хранятся в музее Томского университета)

Фиг. II. Спороподобные микроскопические оболочки из верхнего докембрия (венда) Приднестровья (левый ряд) и кембрия Прибалтики (правый ряд); случайные складки имитируют трехлучевую щель; длина линейки — 10 микрон (фотографии Н. А. Волковой)
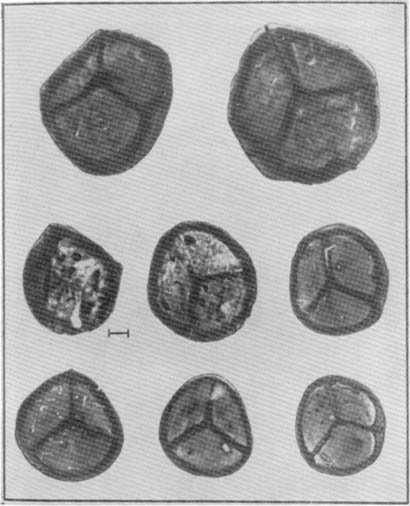
Фиг. III. Споры с настоящей трехлучевой щелью из силура Ливии, длина линейки — 10 микрон (по Д. Б. Ричардсону и Н. Иоаннидес)

Фиг. IV. Проблематичные водоросли вендотениды из верхнего докембрия Ленинграда, собранные при строительстве метро (коллекция Н. А. Волковой); внизу — мельчайшие спороподобные оболочки с трехлучевой складкой из верхов докембрия Ленинградской области, длина линейки — 10 микрон (фотографии Н. А. Волковой)

Фиг. V. Отпечатки коры томиодендрона (Tomiodendron) с рядами листовых подушек и отдельная листовая подушка (слева вверху); образцы собраны в нижнекаменноугольных (острогских) отложениях Кузбасса и северо-востока СССР (два нижних образца)

Фиг. VI. Листья кордаитов (слева вверху и справа) и чешуевидный лист (Nephropsis) слева внизу из пермских отложений Тунгусского бассейна; длина линейки — 2 мм; правая фотография в натуральную величину

Фиг. VII. Ветка ангароптеридия (Angaropteridium) из среднего-верхнего карбона Кузбасса; длина линейки — 1 см (по М. Ф. Нейбург); справа — микроструктура листа руфлории из нижней перми Тунгусского бассейна на отпечатке (А) и реплике (Б); длина линейки — 0,5 мм

Фиг. VIII. Слева вверху и внизу — пыльцу такого типа продуцировали еврамерийские кордаиты и хвойные (по Р. Флорину); справа — пыльца из спорангия Cladostrobus (вверху; см. рис. 9) и того же типа пыльца, извлеченная из породы (внизу)
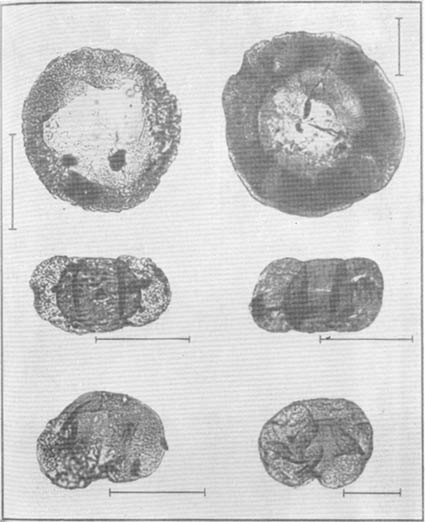
Фиг. IX. Сходные типы ангарской (левый ряд) и гондванской (правый ряд) верхнепалеозойской пыльцы; длина линейки — 50 микрон

Фиг. X. Копролит (слева вверху, длина линейки — 1 см) и извлеченные из него пыльца и обрывки листовой кутикулы с устьицами (длина линейки на микрофотографиях — 50 микрон

Фиг. XI. Эти невзрачные растительные остатки из Афганистана крайне интересны, поскольку они указывают на смешение растений разных флор — катазиатской (а — мутовка листьев членистостебельного типа Lobatannularia, б — лист Taeniopteris), субангарской (в — листья Phylladoderma) и еврамерийской (г — папоротник еврамерийского или катазиатского типа); длина линейки — 5 мм

Фиг. XII. Пельтаспермовый птеридосперм (Madygenopteris) из корвунчанской пермо-триасовой флоры Тунгусского бассейна (коллекция Г. Н. Садовникова)

Фиг. XIII. Войновския из перми Печорского бассейна; длина линейки — 1 см (по М. Ф. Нейбург)

Фиг. XIV. Орган размножения войновскии, ошибочно считавшийся обоеполым (по М. Ф. Нейбург)

Фиг. XV. Отпечатки листьев покрытосеменных и побегов хвойных из нижнемеловых (альбских) отложений Западного Казахстана (коллекция В. А. Вахрамеева)

Фиг. XVI. Раннемеловые покрытосеменные СССР: наверху — соплодие из альбских отложений Западного Казахстана; внизу — лист из аптских отложений Забайкалья, в которых была обнаружена и пыльца покрытосеменных; длина линейки — 1 см (коллекция В. А. Вахрамеева)
---

Оглавление
Сергей Викторович Мейен
Следы трав индейских
ПРЕДИСЛОВИЕ
Глава I
ЗАВОЕВАНИЕ СУШИ
Глава II
ПЕРВЫЙ РАСКОЛ В РАСТИТЕЛЬНОМ ЦАРСТВЕ
Глава III
«ВЫМЕРШИЙ» КЛИМАТ
Глава IV
АНГАРИДА — ДРЕВНИЙ МАТЕРИК
Глава V
ПАЛЕОЗОЙСКИЕ АНТИПОДЫ
Глава VI
ПАССАЖИРЫ ИЛИ ПОСТОЯЛЬЦЫ?
Глава VII
КОРНИ МЕЗОФИТА
Глава VIII
«ОТВРАТИТЕЛЬНАЯ ТАЙНА»
Глава IХ
ЧТО ЕСТЬ ИСТИНА В ИСТОРИИ ЗЕМЛИ?
Глава Х
ВЗГЛЯД В БУДУЩЕЕ
Приложение. Таблицы